
Chiropterotriton cieloensis, Rovito, Sean M. & Parra-Olea, Gabriela, 2015
|
publication ID |
https://doi.org/10.11646/zootaxa.4048.1.3 |
|
publication LSID |
lsid:zoobank.org:pub:A5C40F83-2369-41E7-BB51-A848E77F5A74 |
|
DOI |
https://doi.org/10.5281/zenodo.5614618 |
|
persistent identifier |
https://treatment.plazi.org/id/03FA87CF-FFC0-0054-E68E-FEA7B881FB96 |
|
treatment provided by |
Plazi |
|
scientific name |
Chiropterotriton cieloensis |
| status |
sp. nov. |
Chiropterotriton cieloensis View in CoL sp. nov.
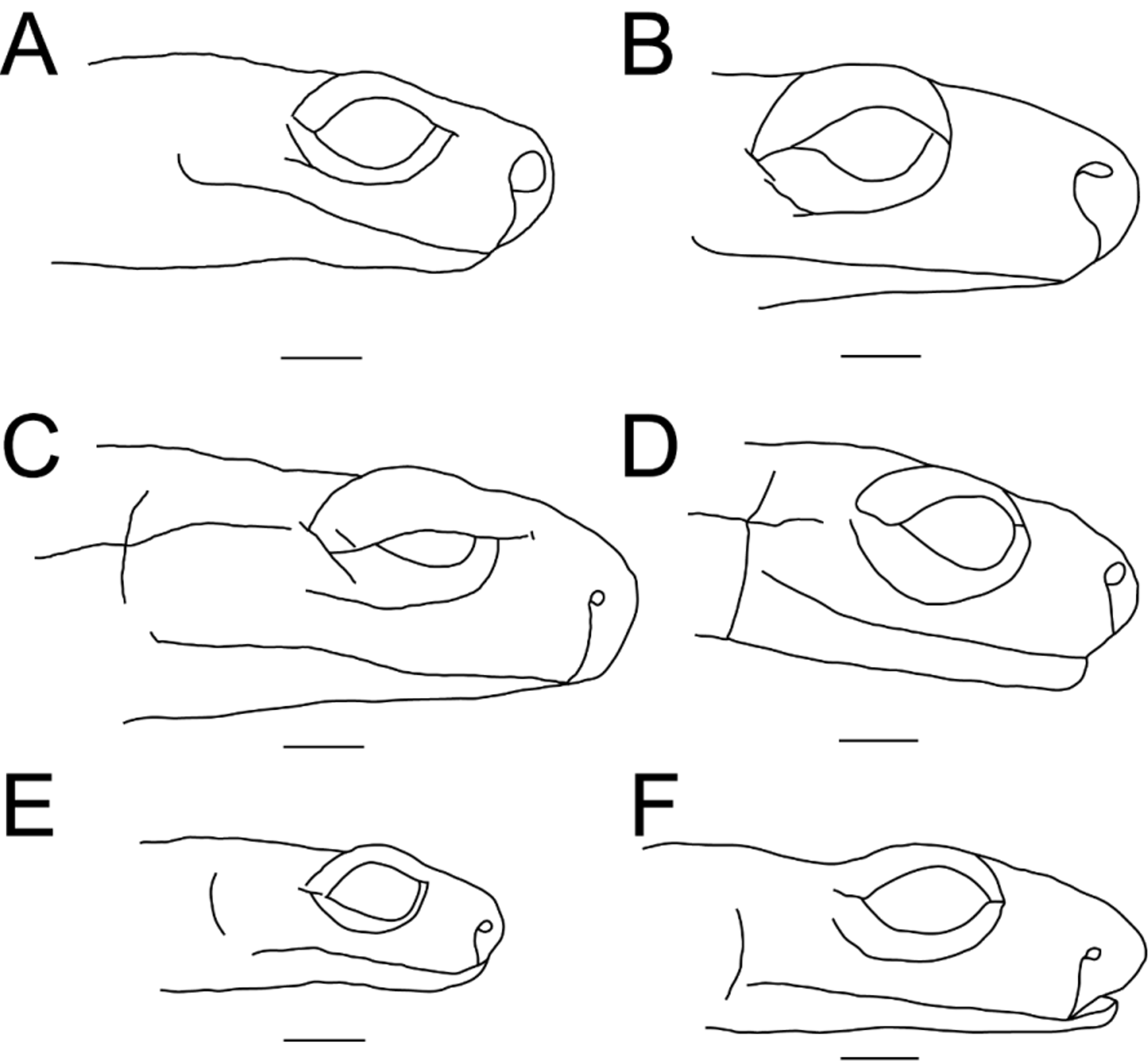
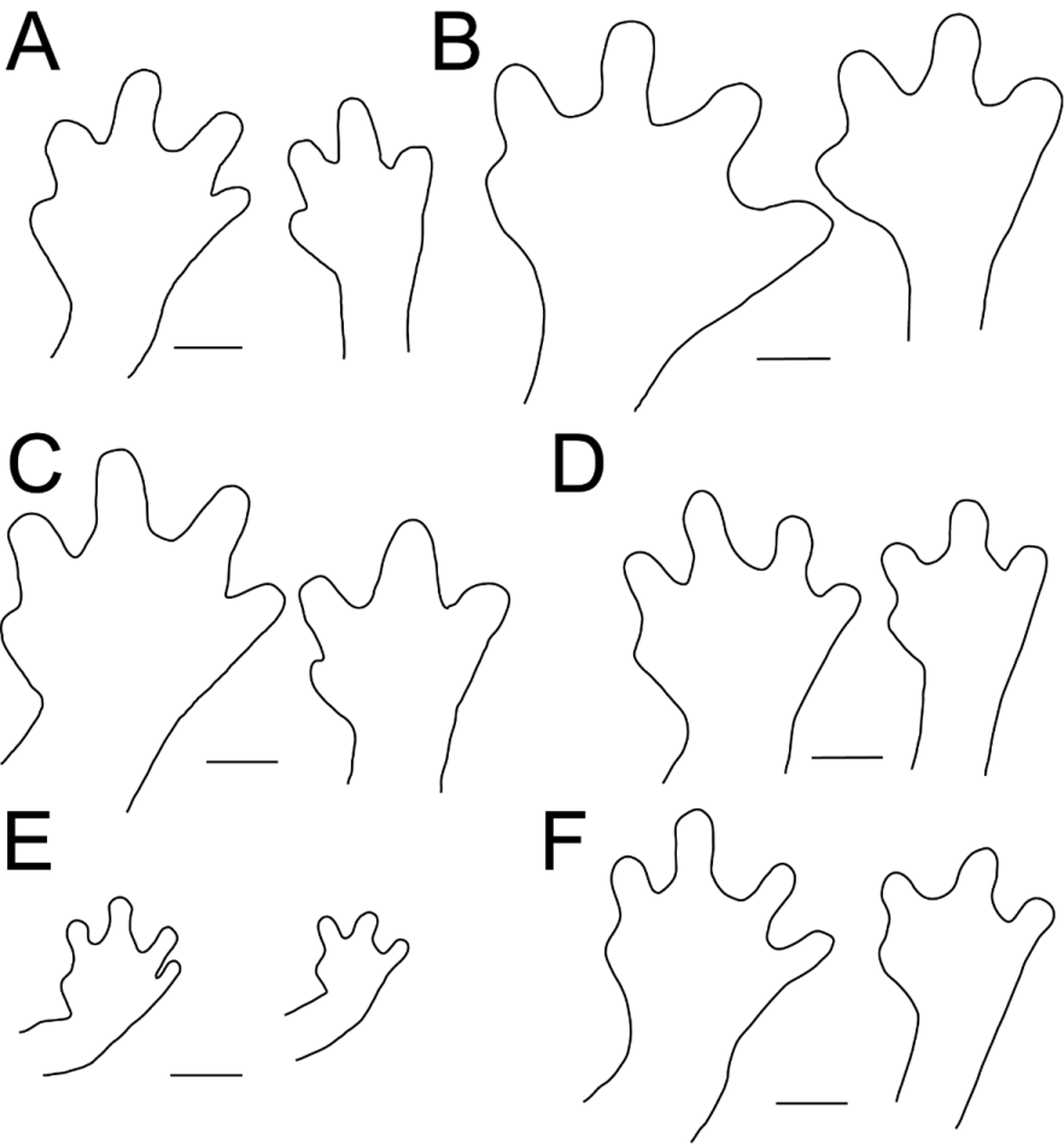
El Cielo Salamander, Salamandra de El Cielo ( Figs. 3 View FIGURE 3 A, 4F, 5F)
Chiropterotriton multidentatus View in CoL (part): Martin, 1958; Rabb, 1958; Darda, 1994; Parra-Olea, 2003. Chiropterotriton View in CoL cf multidentatus View in CoL (part): Campbell, Streicher, Cox, and Brodie, Jr., 2014
Holotype. IBH 29561 (original field number SMR1224), an adult male from 0.2 km E (by air) of Rancho El Cielo, 6.9 km NNW (by air) of center of Gómez Farías, Reserva de la Biósfera El Cielo, Municipio Gómez Farías, Tamaulipas, Mexico, 23.10043° N, 99.19044° W (WGS84 datum), 1174 m elevation, collected 3 August 2010 by Sean M. Rovito, Gabriela Parra Olea, Jorge F. Sánchez Solís, Thomas J. Devitt, Susan Cameron, and Elí García Padilla.
Paratypes. Eleven specimens, all from El Cielo Biosphere Reserve, Tamaulipas, Mexico. Seven males: IBH 28178, El Valle del Ovni, 1 km N (by air) of San José ( 23.05687° N, 99.22837° W); IBH 28184, same locality data as holotype; IBH 28182, Cueva Govenia, 4.3 km NW (by air) of center of Gómez Farías on road to Rancho El Cielo ( 23.0706° N, 99.18823° W); MVZ 158569, 158572, 158573, 173408, Rancho El Cielo, trail to Agua Linda. Four females: IBH 28183, same locality data as holotype. MVZ 158570, 158571, 173409, Rancho El Cielo, trail to Agua Linda.
Referred specimens. IBH 28181, 28185–28189, 29574. UMMZ 111314 – 111316, 102903 – 102906, 105336, 105354, 105363–105368. MVZ 173385, 173386, 173393, 173411.
Diagnosis. Assigned to Chiropterotriton because of small size, slender body, relatively long tail, and presence of sublingual fold, and based on mtDNA sequence analysis. Distinguished from all other described species except C. magnipes by large size of eye relative to head length ( Rabb 1965) ( Fig. 4 View FIGURE 4 ). Additionally, it differs from C. arboreus by having slightly longer limbs relative to body size (LI males: C. arboreus 0.2± 0.63 costal folds (mean±s.d.) vs. C. cieloensis -0.2±0.46; females: C. arboreus 0.8±0.29 vs. C. cieloensis 0.1±0.85) ( Rabb, 1958) and a shorter tail relative to body size (TL/SVL males: C. cieloensis 1.17±0.106 vs. C. arboreus 1.23±0.097; females: C. cieloensis 1.04±0.219 vs. C. arboreus 1.08±0.023 (Rabb, 1958)) and from C. multidentatus by slightly more elliptical nostrils (NL/NW males: C. cieloensis 1.6±0.19 vs. C. multidentatus 1.4±0.22; females: C. cieloensis 1.8±0.2 vs. C. multidentatus 1.5±0.18) and relatively longer limbs in females (LI C. multidentatus females 1.0±0.97). The latter two species are the sister group of C. cieloensis in our phylogenetic analysis. Distinguished from C. chondrostega and C. terrestris by larger body size (SVL males: C. chondrostega 26.3±2.06, C. terrestris 25.4±0.60 vs. C. cieloensis 32.6±1.96; females: C. chondrostega 27.7±2.43, C. terrestris 28.3±1.15 vs. C. cieloensis 31.1±2.73), and from C. dimidiatus by larger size ( C. dimidiatus maximum SVL 29.0), relatively longer tail ( C. dimidiatus TL/SVL 0.77–0.80 ( Campbell et al., 2014)) and more maxillary teeth ( C. dimidiatus MT 12.2±0.99 vs. C. cieloensis 31.4±3.66) and vomerine teeth ( C. dimidiatus VT 6.6±0.56 vs. C. cieloensis 11.5±2.00) in males ( Rabb 1958). Chiropterotriton orculus and C. terrestris have relatively shorter tails than C. cieloensis (TL/ SVL: C. orculus 0.70 –1.03, C. terrestris 0.90–1.07 ( Campbell et al., 2014)), and C. chiropterus has fewer premaxillary-maxillary teeth in males (MT C. cieloensis males 31.4±3.66 vs. about 12 in C. chiropterus ( Campbell et al., 2014)) . Differs from Chiropterotriton magnipes by much smaller size, fewer maxillary and vomerine teeth, and much narrower feet ( Rabb 1965) and C. infernalis by reduced foot webbing and narrower hands and feet (FW males: C. cieloensis 3.2±0.40 vs. C. infernalis 4.2±0.70; Fig. 5 View FIGURE 5 ). Differs from sympatric C. cracens by larger body size (SVL C. cracens males SVL 25.7±1.98, females 27.4±3.30) and by relatively longer limbs (LI C. cracens males 1.8±0.65, females 2.1±0.89) ( Rabb 1958). Chiropterotriton lavae has fewer premaxillary-maxillary teeth in males (MT C. lavae males 6–10 ( Campbell et al., 2014)) and is larger (SVL C. lavae males 36–50 ( Campbell et al., 2014)). Differs from C. mosaueri by smaller body size (max SVL C. cieloensis 36.1 vs. min SVL C. mosaueri 39.6), relatively shorter tail (TL/SVL C. mosaueri 1.31±0.159), and fewer maxillary and vomerine teeth (MT C. mosaueri 56–69) ( Woodall 1941; Rabb 1958; Campbell et al., 2014). Distinguished from C. priscus by having relatively longer limbs (LI C. priscus males 3.2±0.40, females 3.7±0.30) and a less robust body, and from C. miquihuanus by much smaller nostril (NL males: C. miquihuanus NL 0.82±0.07 vs. C. cieloensis 0.19±0.02; females: C. miquihuanus 0.85±0.04 vs. C. cieloensis 0.21±0.02; Fig. 4 View FIGURE 4 ).
Description of the holotype. Moderately sized (SVL 33.8 mm) adult male with relatively long tail (TL/SVL 1.09), thin, approximately round at base, tapering toward end. Relatively wide head (HW 4.8 mm), with eyes protruding just beyond margin of jaw when viewed from above. Head relatively flattened, nostril moderately sized and oval shaped (NL/NW 1.37), snout blunt, nasolabial protuberances well developed. Relatively large, nearly circular mental gland present. Premaxillary teeth (2) pierce upper lip. Vomerine teeth (11) extend in a line nearly to outer margin of choanae. Limbs relatively long (LI 0.5), hands and feet moderately sized with well-developed interdigital webbing. Webbing extends just onto penultimate phalanx. Toe tips bluntly rounded with well develop subterminal pads. Phalangeal formulae: manus 1-2-3-2, pes 1-2-3-3-2. Digits in order of increasing length: manus I-IV-II-III, pes I-V~II-IV-III.
Measurements (in mm), limb interval and tooth counts of the holotype. SVL 33.8, TL 36.8, AX 17.7, HL 6.7, HW 4.8, head depth at posterior angle of jaw 2.4, snout projection beyond mandible 0.4, IO 1.8, EN 1.5, IN 1.7, eyelid length 2.2, eyelid width 1.4, horizontal orbit diameter 1.9, FLL 8.2, HLL 9.0, snout to forelimb 9.7, width of right hand 2.1, FW 3.2, T3 1.4, T5 0.7, snout to anterior angle of vent 30.2, tail width at base 1.9, tail depth at base 2.1, shoulder width 3.75. NL 0.26, NW 0.19. Adpressed limbs separated by 0.5 costal folds. Premaxillary teeth 2, maxillary teeth 24, vomerine teeth 11.
Variation. One of the male paratypes is slightly larger than the holotype ( 36.1 mm); this specimen (IBH 28182) has a much longer tail ( 48.1 mm) and longer limbs than the holotype. The nostril shape in this species appears to vary from oval (NL/NW 1.4) to elongated elliptical (NL/NW 2.0). Several of the paratypes have substantially more premaxillary-maxillary teeth than the holotype, up to 56 premaxillary-maxillary teeth in female paratype MVZ 158571.
Coloration of the holotype in life (from photos). Dorsal surface of head and dorsum Ferruginous (35) with extensive Cream Yellow (82) speckles. Dorsal surface of tail same colors with fewer, more scattered speckles. Upper side of limbs Ferruginous with some Cream Yellow speckles. Gular region Light Russet Vinaceous (246). Venter and underside of limbs immaculate Cinnamon Drab (259). Ventral side of tail pale Raw Umber (280). Underside of toe tips Crimson (65). Iris Light Yellow Ocher (13).
Coloration of the holotype in alcohol. Dorsum and dorsal surface of tail uniform medium brown. Dorsal surface of head slightly lighter brown, eyelids dark brown, dorsal coloration extends to lateral midline where it changes to pale grey brown. Dorsal surface of limbs light yellowish brown extending to onto upper side of hands and feet. Gular region ventral surface of tail and ventral side of limbs pale yellowish cream color with uniform scattering of tiny dark brown flecks. Underside of hands and feet pale yellowish brown.
Habitat and distribution. Chiropterotriton cieloensis is known only from moderate to higher elevations (approximately 1000–1860 m elevation; Martin 1958) of the El Cielo Biosphere Reserve in Tamaulipas, Mexico. This mountain range is known as the Sierra de Guatemala ( Martin 1958). The species is found in broadleaf cloud forest and has been taken from arboreal bromeliads and caves, and has been found occasionally under the bark of rotten logs ( Martin 1958). It occurs in sympatry with C. cracens , which is also endemic to the El Cielo Biosphere Reserve. Chiropterotriton cieloensis appears to be more common in bromeliads than C. cracens , and the latter does not occur in caves ( Martin 1958). Aquiloeurycea scandens is commonly found in caves and occasionally in bromeliads with C. cieloensis , and a member of the Aquiloeurycea cephalica group and Isthmura bellii are also found in sympatry with C. cieloensis . We also assign specimens to this species from a lower elevation cave ( 420 m elevation) at Aserradero El Paraíso, in tropical deciduous forest ( Martin 1958; Rabb 1958). The species is unlikely to occur more widely. Chiropterotriton multidentatus is found in highland areas to the southwest, and C. miquihuanus and C. infernalis are found to the northwest. El Cielo is near the northern limit of cloud forest in Mexico, so the species is unlikely to occur further north.
Etymology. The specific epithet makes reference to the El Cielo Biosphere Reserve, where the species is found. El Cielo means “heaven” in Spanish, an appropriate term for the spectacular cloud forest habitat that is protected by the biosphere reserve.
Remarks. While it has long been known that there are two species present at El Cielo, distinguishing the two has not always been easy, especially before the advent of molecular systematic methods. The assignment of juveniles to species based on morphology was especially difficult. The allozyme study of Darda (1994) clearly showed the presence of two species at El Cielo, but the mtDNA study of Parra-Olea (2003) found that all individuals had nearly identical mtDNA sequences, indicating the presence of only one species. Our expanded mtDNA study, however, shows the presence of two species from El Cielo, in agreement with the allozyme results. We assigned a single individual with a sequence different from all of those from Parra-Olea’s study to Chiropterotriton cracens based on morphological features, while all other individuals in our study had sequences closely matching those from the species previously referred to as C. multidentatus . This means that the El Cielo sequences from Parra-Olea et al. (2003) correspond to the second species, which we name here as C. cieloensis . Of the individuals used in the Darda (1994) study from El Cielo, Parra-Olea included eight in her study. Three of the specimens identified as C. multidentatus in the allozyme study are in the clade of C. cieloensis samples from our study, and morphological data also support their assignment to C. cieloensis . Four more of the specimens are small juveniles or could not be located, but both allozymes and mtDNA support their assignment to C. cieloensis . Only a single specimen remains in conflict between molecular studies: MVZ 173380, was assigned to C. cracens by Darda (1994), however, this same specimen was assigned to C. multidentatus by Parra-Olea (2003). This individual corresponds more closely to C. cracens in morphology, especially in having relatively short limbs (LI 2.0). It is possible that an error in one of the two studies resulted in the disagreement between allozyme and mtDNA results. We choose not to include this individual in our morphological comparisons because of the difficulty in assigning it to either of the two species.
Most specimens in other museums from El Cielo currently classified as C. multidentatus are most likely assignable to C. cieloensis . Without examining these specimens, however, we have chosen not to refer them to the species, with the exception of specimens identified by Rabb (1958). We assign specimens from the lower elevation population at Aserradero El Paraíso to this species based on the morphological comparisons of Rabb (1958), who determined that the El Paraíso specimens were very similar morphologically to those from El Cielo and likely belonged to the same species. Molecular comparisons would be instrumental in confirming the conspecific status of this population with C. cieloensis , but we were unable to reach the El Paraíso locality during our fieldwork to obtain tissue samples for phylogenetic analysis.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |