
Anaphes Haliday 1833
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4376.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:04E7DF99-B411-4F40-AA15-50299868BE5D |
|
DOI |
https://doi.org/10.5281/zenodo.5949521 |
|
persistent identifier |
https://treatment.plazi.org/id/03FE87E6-FF80-FFCE-C282-EBB5CD4B384D |
|
treatment provided by |
Plazi |
|
scientific name |
Anaphes Haliday 1833 |
| status |
|
Anaphes Haliday 1833 View in CoL View at ENA
Synonymy. Given in Huber (1992); the junior synonyms may also be determined from Tables 2, 3, and 6.
Type species. The type species is Anaphes fuscipennis Haliday ( ICZN 2017). Huber et al. (2011, 2014) had petitioned successfully to have the type species changed.
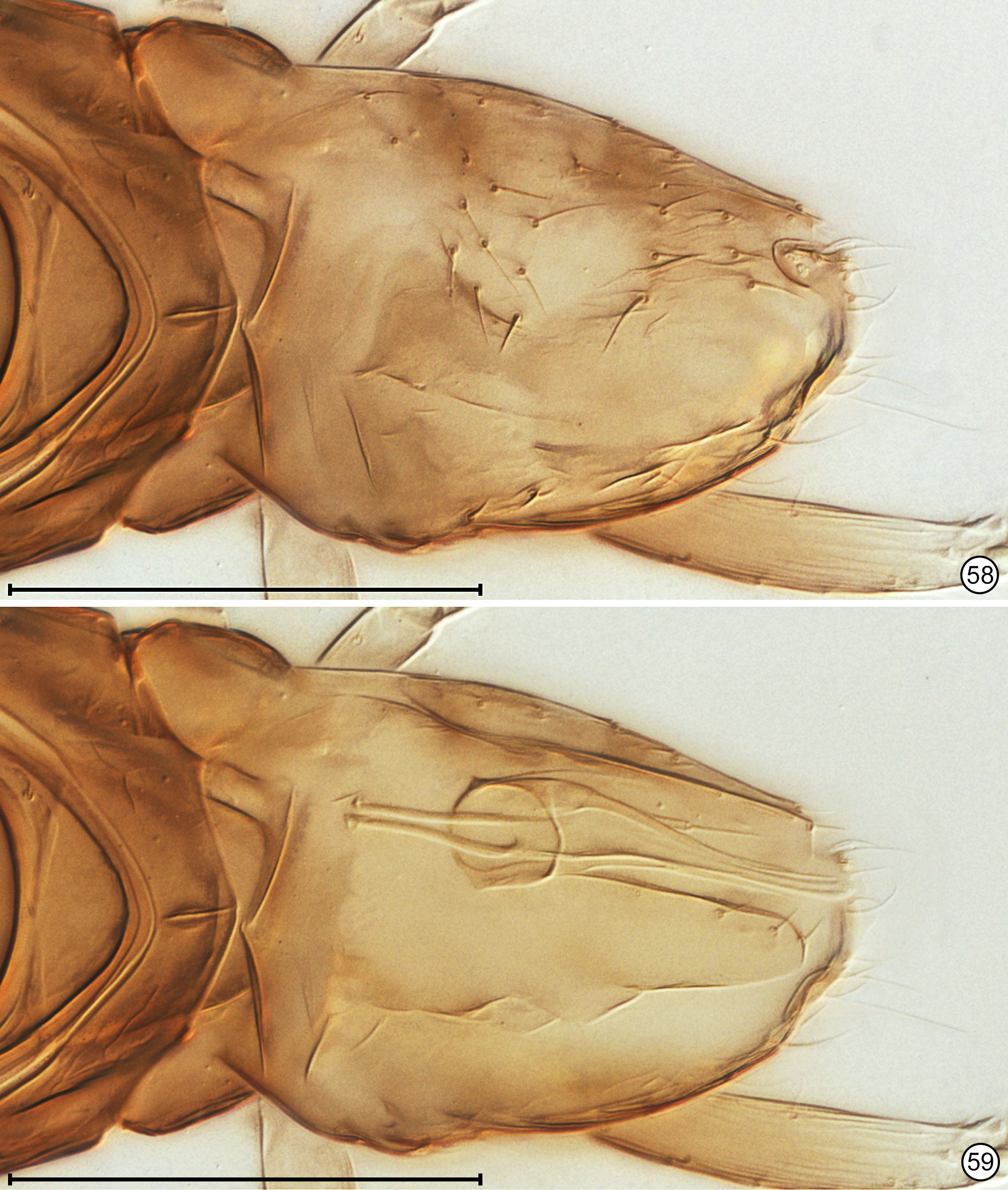
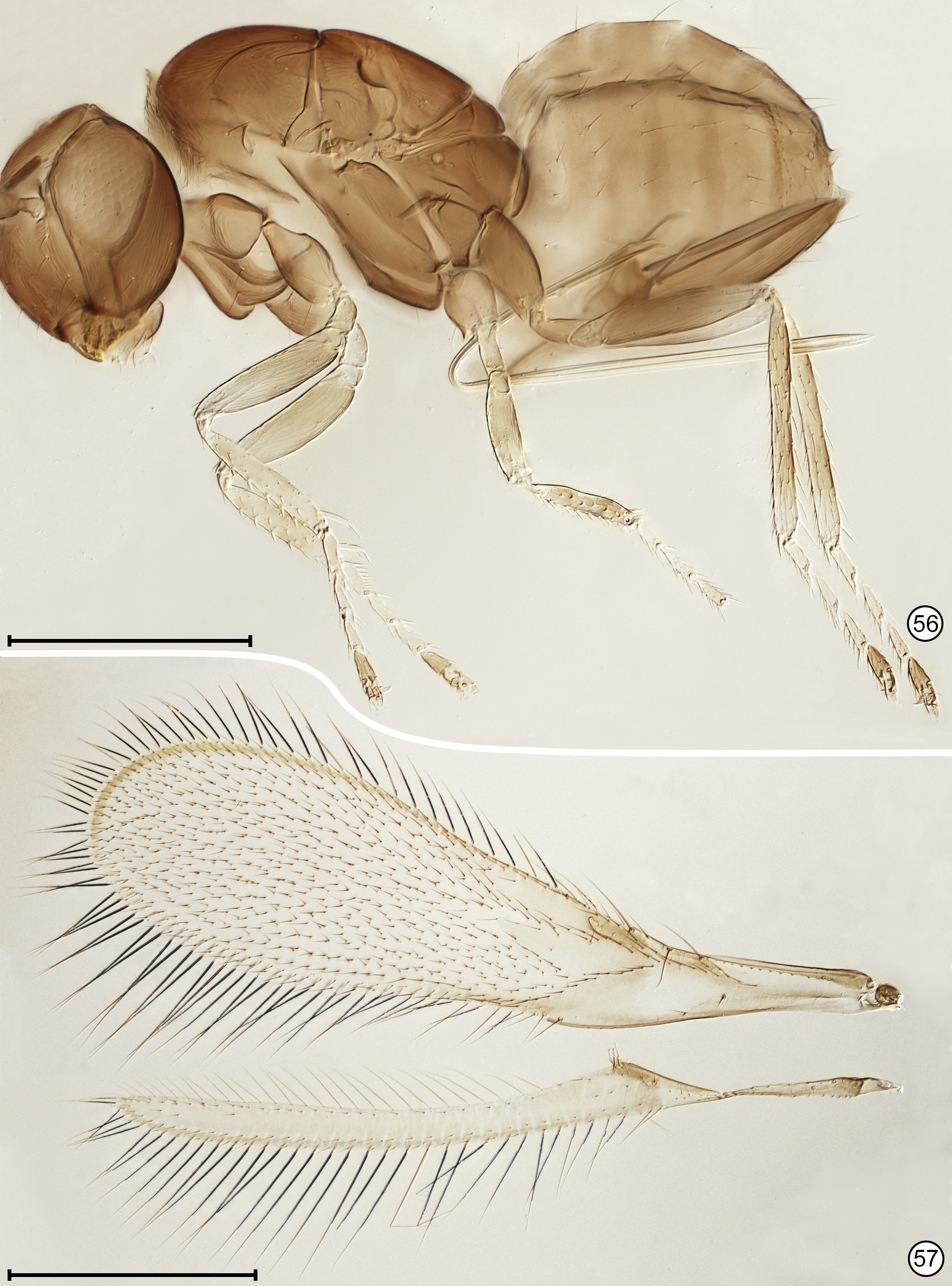
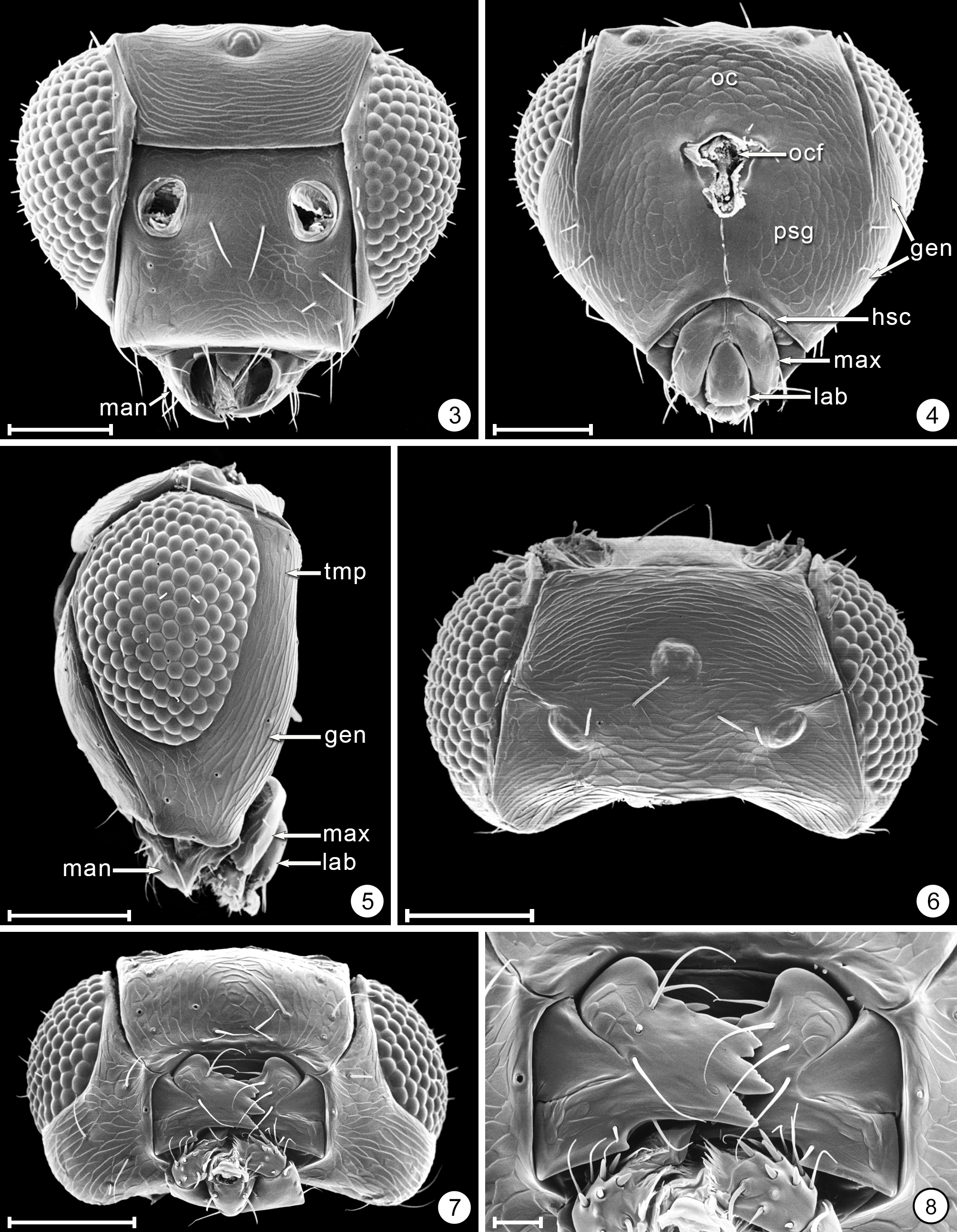
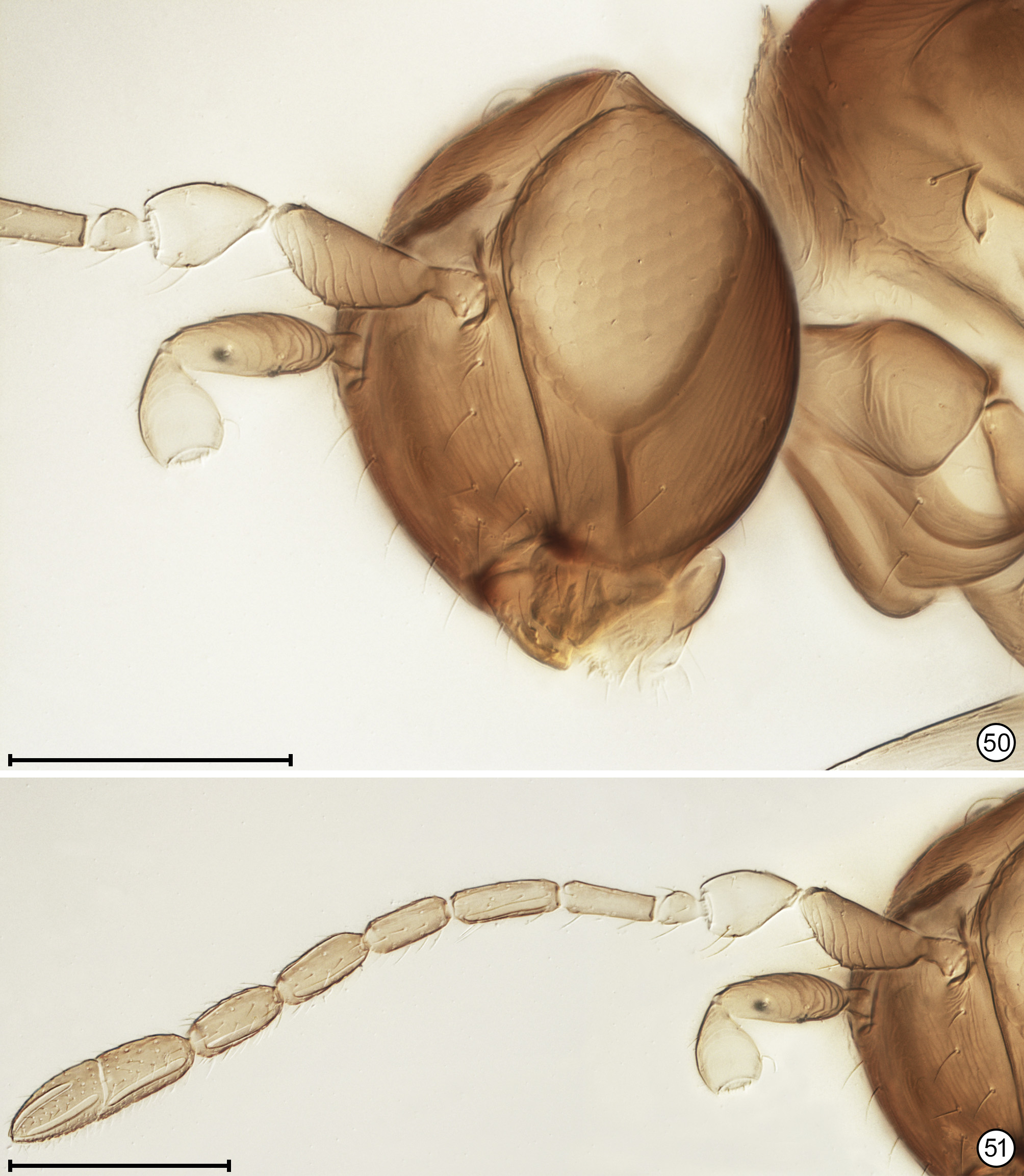
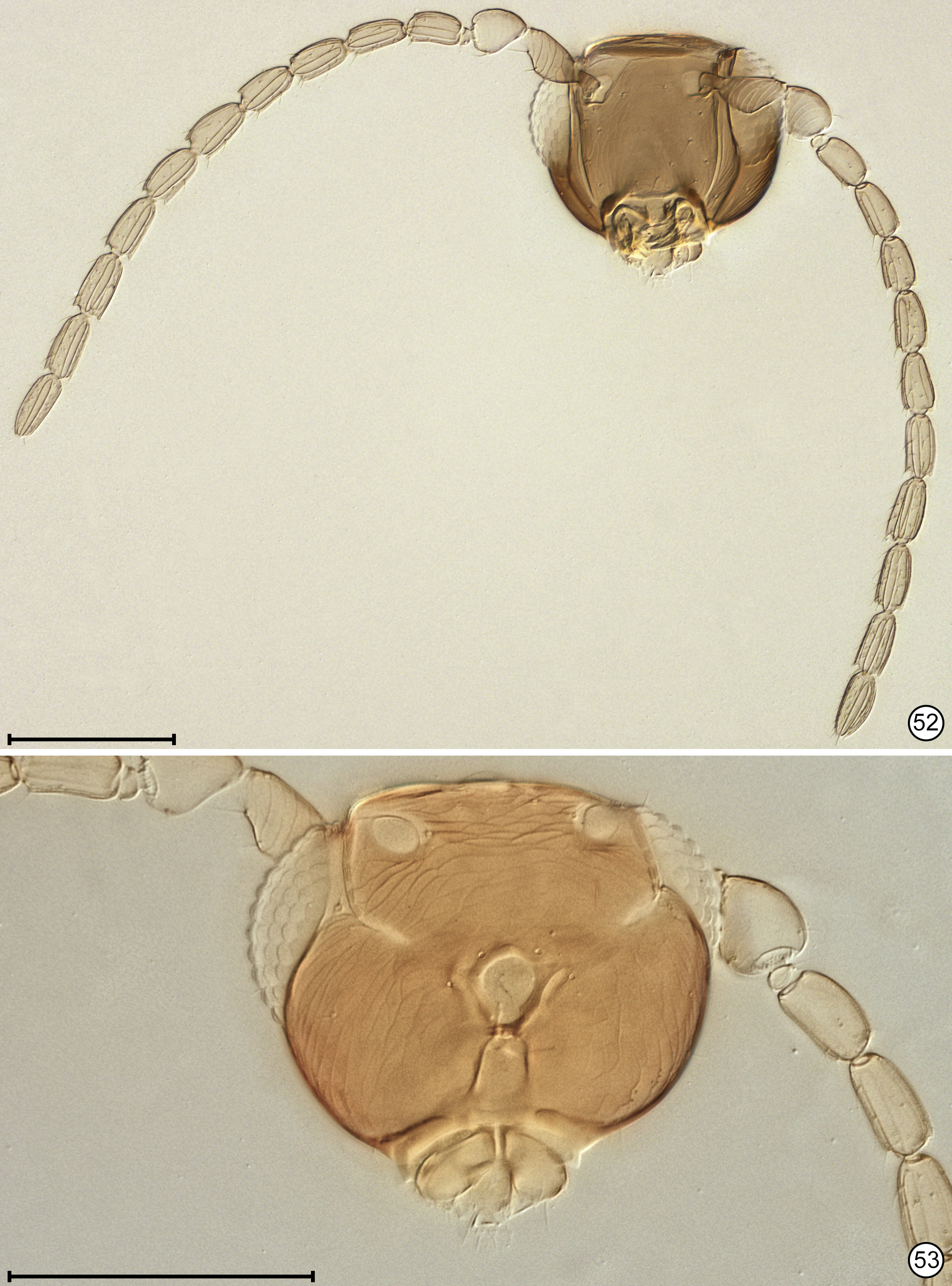
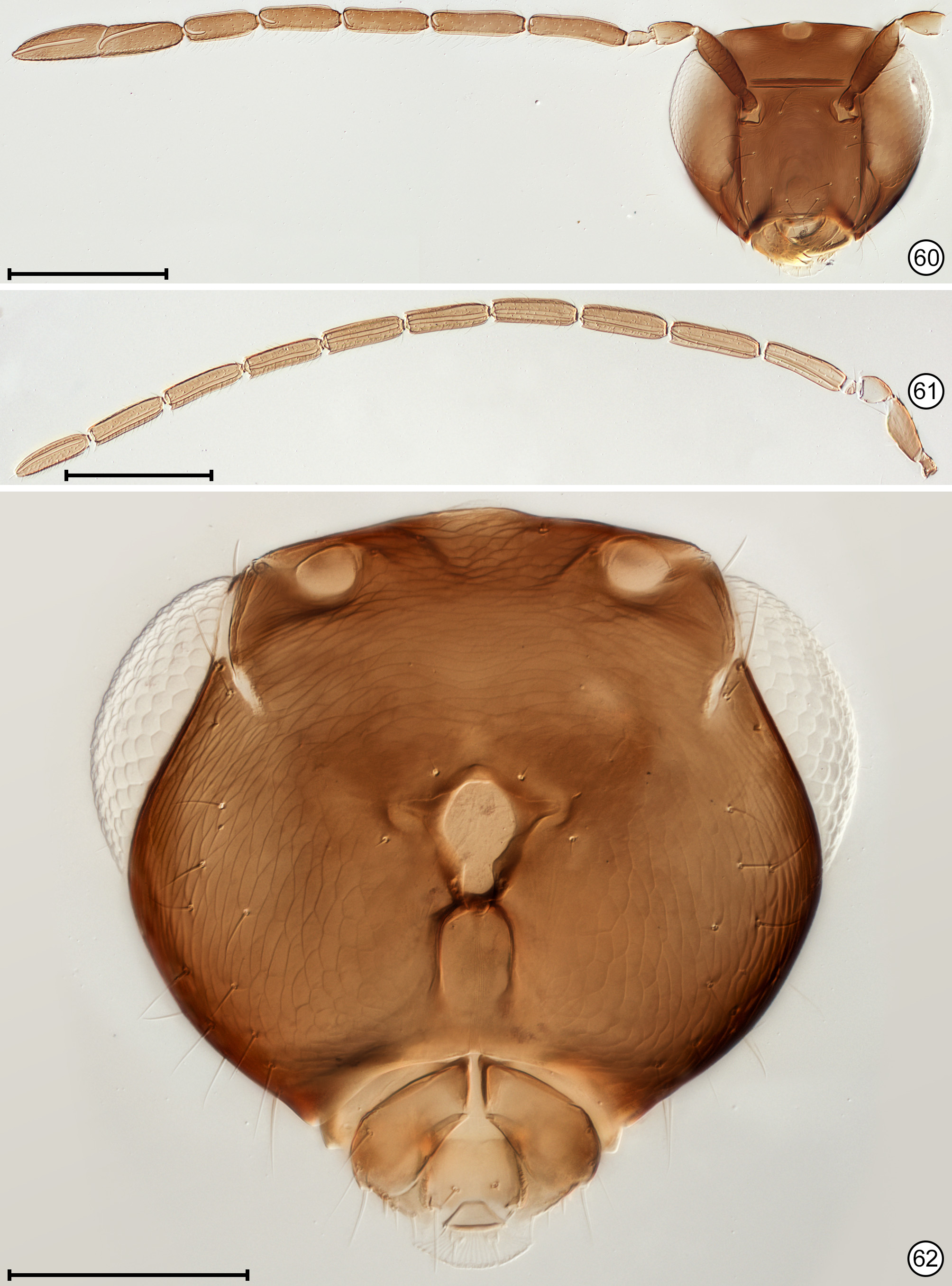
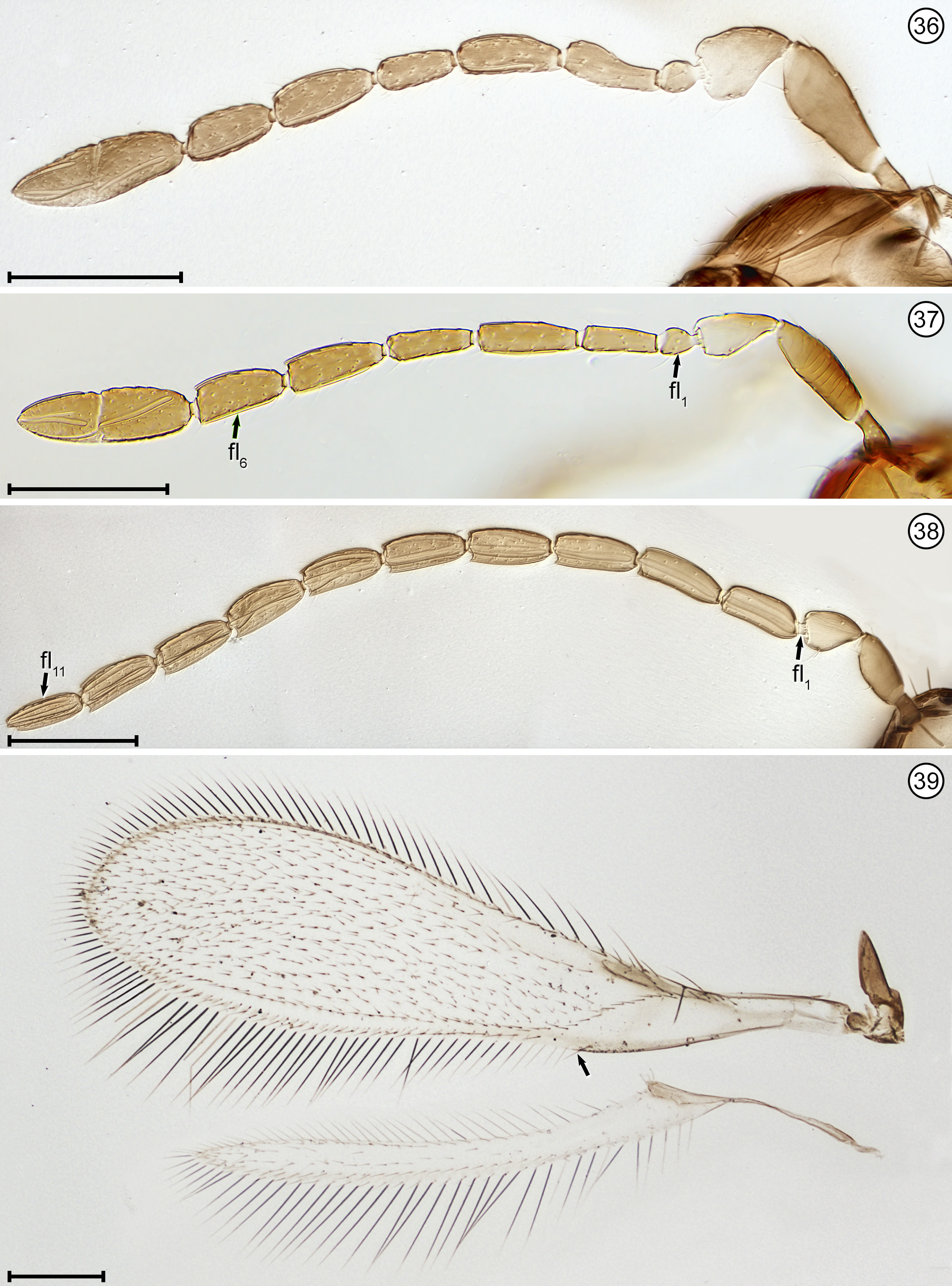
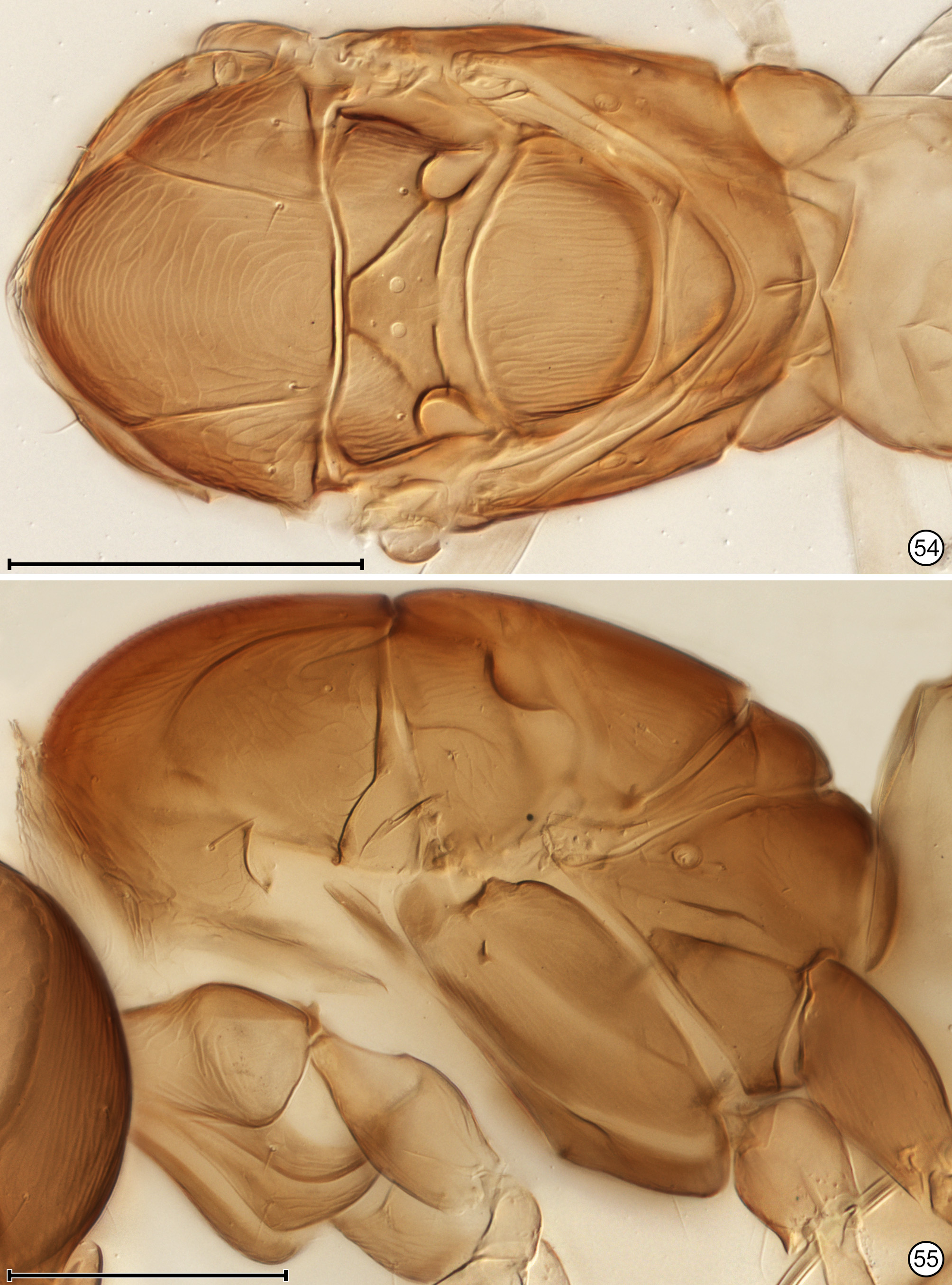
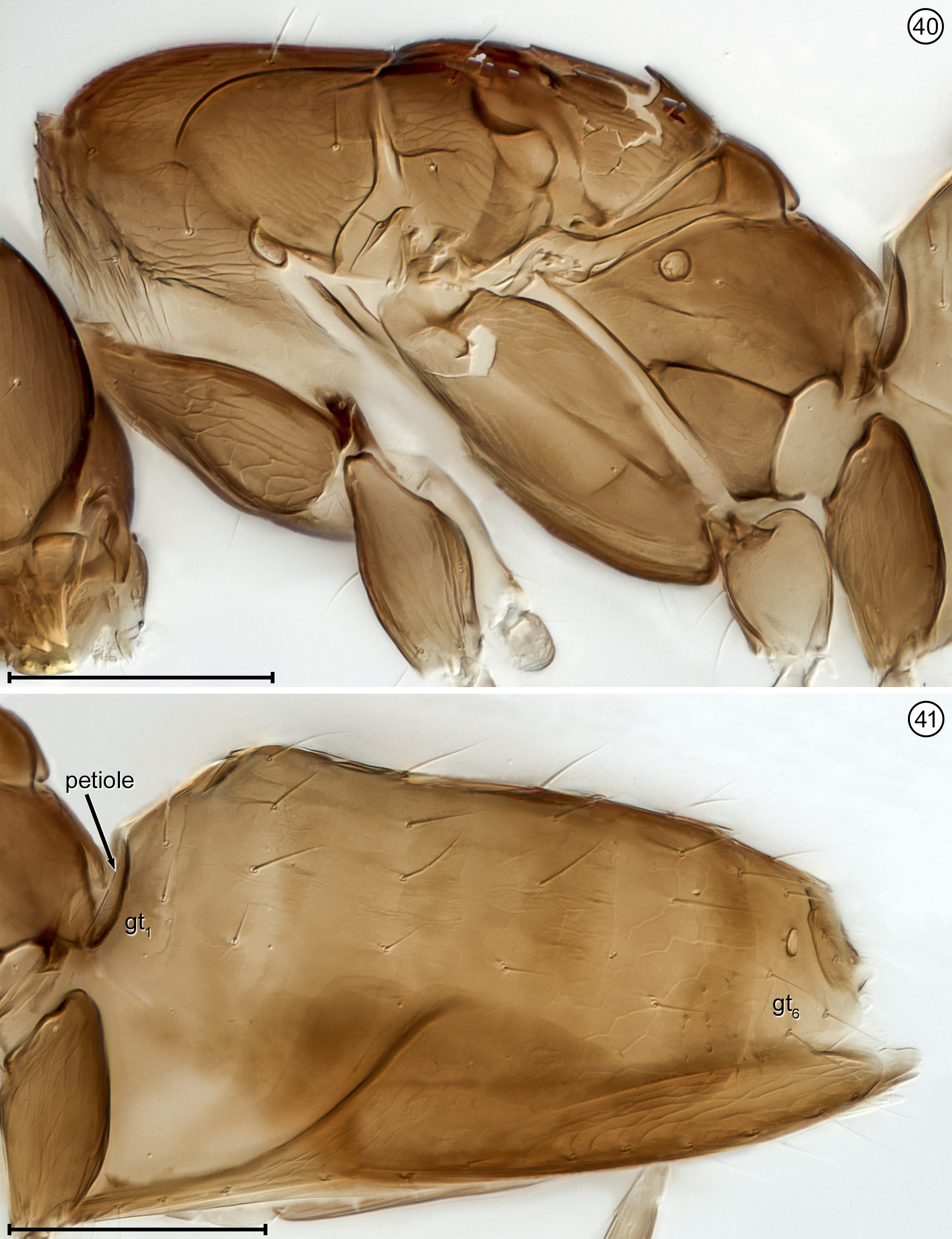
Diagnosis. Both sexes. Body almost always uniform black or dark brown, exceptionally with metasoma almost yellow; appendages usually lighter brown, often with off-white apices of femora, tibiae and tarsi except apical tarsomere ( Figs 45 View FIGURES 44, 45 , 56 View FIGURES 56, 57 ). Head. Face without subantennal grooves ( Fig. 3 View FIGURES 3–8 ); malar sulcus absent, at most indicated exteriorly by slight change in sculpture ( Fig. 5 View FIGURES 3–8 ), but visible internally in cleared slide mounts ( Figs 34, 35 View FIGURES 32–35 , 50 View FIGURES 50, 51 , 52, 53 View FIGURES 52, 53 , 60 View FIGURES 60–62 ); vertex without stemmaticum but with a groove from anterior margin of posterior ocellus to supraorbital trabecula ( Fig. 6 View FIGURES 3–8 ); occiput with occipital groove [evidently an extension onto the occiput of the supraorbital suture of the vertex, incorrectly named the occipital suture in Huber (2006, fig. 32)] usually in line with supraorbital suture and extending ventrally to about level of lower margin of occipital foramen ( Fig. 4 View FIGURES 3–8 ) but sometimes short and angled inwardly towards occipital foramen ( Figs 33 View FIGURES 32–35 , 62 View FIGURES 60–62 ). Mandible with 3 teeth, the dorsal tooth sometimes subdivided, appearing serrate, i.e. with 2 or 3 small apical teeth ( Figs 7, 8 View FIGURES 3–8 ) but exceptionally (quinquearticulatus) with 2 large ventral teeth distinctly contrasting with 3 small, dorsal teeth ( Figs 24, 25 View FIGURES24–28 ); maxilla and labium as in Figs 26 and 27 View FIGURES24–28 . Wings. Wings fully developed, with posterior margin straight to slightly in basal half and slightly outcurved in apical half ( Figs 39 View FIGURES 36–39 , 57 View FIGURES 56, 57 , 72 79 View FIGURES71, 72 View FIGURES 73, 74 View FIGURES 75, 76 View FIGURE 77 View FIGURE 78 View FIGURE 79 ) to distinctly ( Fig. 31 View FIGURES 29–31 ) incurved, with apex slightly asymmetrical (anterior margin usually more curved than posterior margin), and with both dorsal and ventral surfaces almost always uniformly covered with microtrichia except for two more or less distinct bare areas (the marginal and medial spaces, small in Fig. 72 View FIGURES71, 72 ) separated by an oblique row of microtrichia extending from just behind apex of stigmal vein towards wing apex ( Figs 39 View FIGURES 36–39 , 57 View FIGURES 56, 57 ); wing retinaculum with a socketed seta near apex ( Fig. 39 View FIGURES 36–39 ). Mesosoma. Anterior scutellum ( Figs 29, 30 View FIGURES 29–31 , 54 View FIGURES 54, 55 , 63 View FIGURES 63, 64 ) without setae, with campaniform sensilla separated by about 2 or 3× their own diameter from each other and well separated from transscutal articulation, with fenestra a small triangle with rounded apices ( Fig. 43 View FIGURES 42, 43 , faintly indicated since below surface), and with frenum entire ( Figs 13 View FIGURES 9–16 , 29 View FIGURES 29–31 ); propodeum with a median longitudinal groove ( Figs 13 View FIGURES 9–16 , 29 View FIGURES 29–31 , 43 View FIGURES 42, 43 , 46 View FIGURES 46, 47 , 54 View FIGURES 54, 55 , 58, 59 View FIGURES58, 59 , 63 View FIGURES 63, 64 ). Metasoma. Petiole a thin, narrow and almost vertical crescent ( Figs 40, 41 View FIGURES 40, 41 , 48 View FIGURES 48, 49 ) longitudinally divided medially ( Figs 46, 47 View FIGURES 46, 47 ); gaster with gt1 divided medially by a longitudinal groove ( Figs 17 View FIGURES 17–23 , 29 View FIGURES 29–31 , 43 View FIGURES 42, 43 , 46 View FIGURES 46, 47 ).
Female. Antenna. Clava 1- or 2-segmented, funicle 6-segmented ( Fig. 37 View FIGURES 36–39 ) or, in one species, 5-segmented ( Fig. 28 View FIGURES24–28 ); when 6-segmented, fl1 usually the shortest segment ( Figs 9, 10 View FIGURES 9–16 ). Genitalia. Ovipositor often projecting anteriorly under mesosoma, apparently enclosed in a somewhat membranous anterior extension or sac of gaster perhaps consisting mainly of gs6 ( Figs 15 View FIGURES 9–16 , 18 View FIGURES 17–23 , 56 View FIGURES 56, 57 , 67–70 View FIGURES 67, 68 View FIGURES69, 70 ).
Male. Antenna. Flagellum distinctly 11-segmented only in species placed in Anaphes (Yungaburra) whereas in most nominal species flagellum appearing 10-segmented because fl1 minute and either with only 1 seta ( Figs 11, 12 View FIGURES 9–16 , 38 View FIGURES 36–39 ) or with a seta and 1 transverse mps ( Fig. 61 View FIGURES 60–62 , Huber & Prinsloo 1990, figs 13, 14). Genitalia. Parameres thin, each almost as long as aedeagus and with a long seta more or less midway along paramere length ( Figs 22, 23 View FIGURES 17–23 , 47 View FIGURES 46, 47 , 49 View FIGURES 48, 49 , 59 View FIGURES58, 59 , 71 View FIGURES71, 72 ).
Variation. A teratological specimen ( UCRC) from Greece with 4 funicle segments on one antenna and 5 on the other is known. Both antennae have fl1 + fl2 almost completely fused and one antenna also has fl4 + fl5 almost completely fused ( Fig. 103 View FIGURE 102–104 ). All the tarsi are clearly 3-segmented ( Fig. 102 View FIGURE 102–104 ). Otherwise the wings ( Fig. 104 View FIGURE 102–104 ) and body are normal for an Anaphes specimen.
Subgeneric concepts. As mentioned above, Debauche (1948) proposed two subgenera, A. ( Anaphes ) and A. ( Patasson ), for females with a 1- or 2-segmented clava, respectively, and provided a discussion on how they differ in other, more subtle ways. Ogloblin (1962) proposed A. ( Austranaphes ) for species from Argentina with a 2- segmented clava and males with a clearly 11-segmented flagellum. Huber (1992) treated both Debauche subgenera as species groups within Anaphes sensu stricto and also proposed two subgenera, A. ( Anaphes ), and A. ( Yungaburra ) for most of the southern hemisphere species, with Ogloblin’s A. ( Austranaphes ) treated as one species group in A. ( Yungaburra ). A major problem with Huber’s classification is that the number of flagellomeres bearing mps in the male antenna must be known to classify a species into one or other subgenus. Males are so rarely associated with females that it is better to return to Debauche’s subgeneric classification and reclassify the morphologically diverse species that Huber classified in A. ( Yungaburra ) into one or other of Debauche’s subgenera. This, incidentally, would also be preferable for biological control workers, where Patasson , either as a genus or a subgenus, was used extensively prior to Huber (1992). Patasson should only be used as a subgenus of Anaphes , not a genus.
Alternatively, one could eliminate subgenera entirely and retain only species groups within Anaphes , reclassifying all the species previously placed in A. ( Yungaburra ) into them. If classified within the fuscipennis species group, for example, the species previously placed in A. ( Yungaburra ) could perhaps be distinguished from other species in the group using features such as size of the marginal and medial spaces (often considerably reduced, e.g., in nitens ), mps shape in females (often curved apically, e.g., in nitens ) and sculpture of the head and mesosoma (often with isodiametric reticulations, as in quinquearticulatus).
Finally, one could propose an entirely new species group classification but the problem would be to delimit them properly so they are easily identifiable. Huber’s subgenera cannot be distinguished based on the number of claval segments in females. Some undescribed Anaphes from Australia and one from Indonesia, as well as saintpierrei, have a 1-segmented clava in contrast to other, undescribed Australian region species and nitens , which have a 2-segmented clava. The former would key to the amplipennis group of Anaphes (Yungaburra) which includes the South American species only ( Huber 1992) and nitens obviously would key to his nitens group of A. ( Yungaburra ), in which he included only the Australian-region species. We emphasize again that the subgeneric classification of Anaphes proposed by Huber (1992) is therefore rejected in favour of a return to that of Debauche (1948), i.e., with the subgenera A. ( Anaphes ) and A. ( Patasson ). The species classified previously in A. ( Yungaburra ), are reclassified as follows: amplipennis, fijiensis, nunezi and pucarobius into A. ( Anaphes ), and galtoni, inexpectatus , nitens , and tasmaniae into A. ( Patasson ) ( Tables 3 and 6).



















| UCRC |
University of California, Riverside |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.