
Theropods
|
publication ID |
https://doi.org/ 10.7717/peerj.4417 |
|
DOI |
https://doi.org/10.5281/zenodo.5614773 |
|
persistent identifier |
https://treatment.plazi.org/id/03FE9007-FFA0-8311-FE21-FE127944D531 |
|
treatment provided by |
Plazi |
|
scientific name |
Theropods |
| status |
|
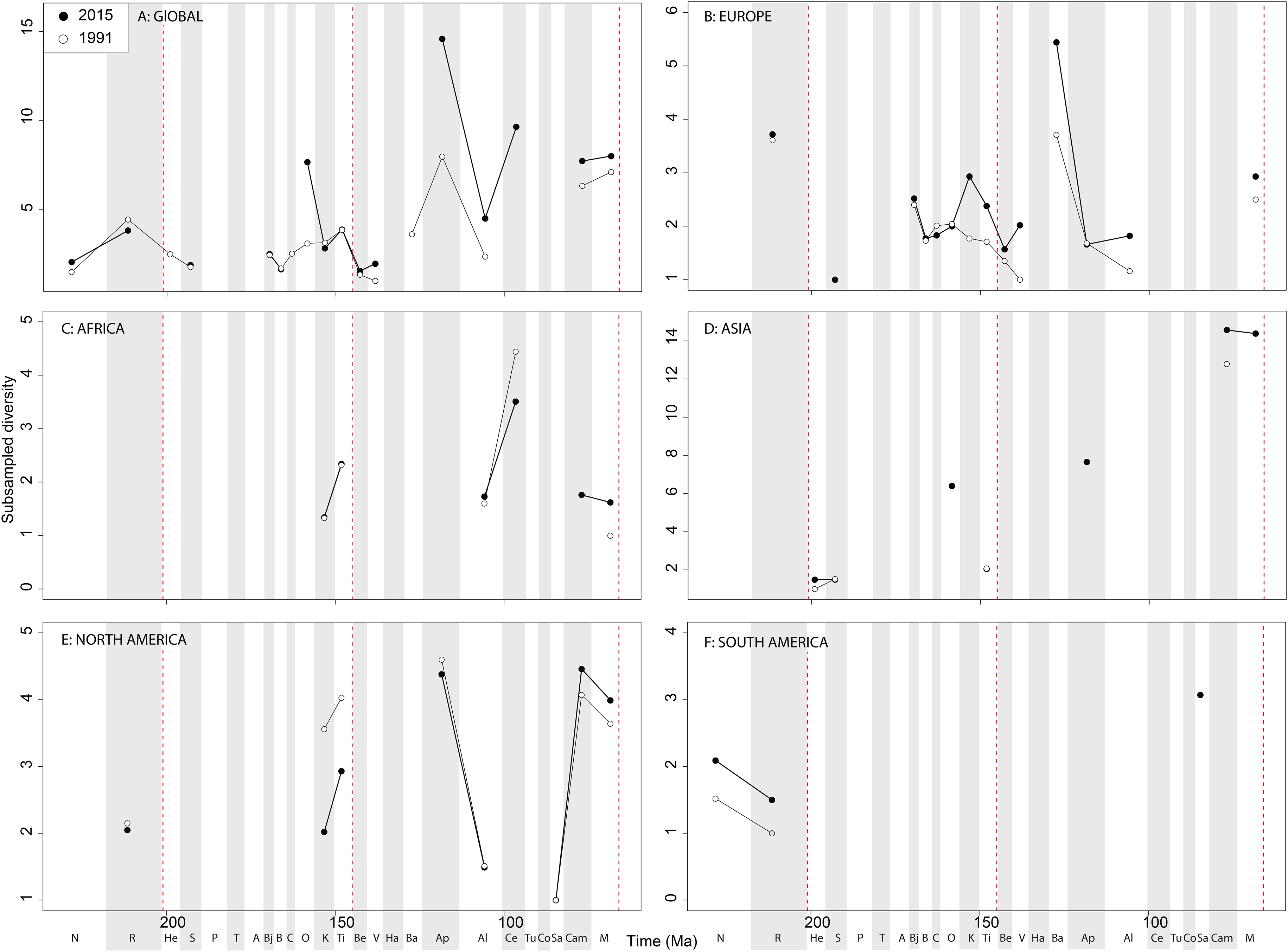
As already shown elsewhere ( Barrett, McGowan & Page, 2009; Brusatte et al., 2012), ‘global’ theropod diversity trends are overall very similar to that of Ornithischia , with subsampled diversity increases during the Late Jurassic (Oxfordian and Tithonian peaks punctuated by a Kimmeridgian decline), late Early Cretaceous (Aptian), early Late Cretaceous (Cenomanian) and latest Cretaceous. Moderately high Middle and Late Jurassic diversity subsampled levels represent the radiation of major avetheropodan clades, and a wealth of new discoveries in recent years, particularly from Asia ( Upchurch et al., 2011; Xu et al., 2011; Carrano, Benson & Sampson, 2012; Benson et al., 2014; Tennant, Mannion & Upchurch, 2016b).
European subsampled theropod diversity is more constant than in other regions, with a Bajocian peak followed by a Bathonian–Oxfordian trough, and a Kimmeridgian peak followed by a Tithonian to Valanginian drop. This can, at least in part, be explained by an abundance of well-sampled Late Jurassic formations from across Western Europe ( Upchurch et al., 2011; Benson et al., 2013; Tennant et al., 2016). Barremian diversity is increasing rapidly through publication history, and is now as high as calculated for the Kimmeridgian. As with the Late Jurassic, at least part of this signal represents the influence of a Lagerstätten effect (e.g. Las Hoyas, Spain) ( Buscalioni et al., 2008; Upchurch et al., 2011; Sánchez-Hernández & Benton, 2012), highlighting that single, well-sampled formations can have a profound historical effect on our understanding of regional diversity patterns, even when subsampling methods are applied. The European Aptian–Albian record is increasing slower through time compared to the Campanian– Maastrichtian. However, this might possibly change in the future, as the ichnological record in southern Europe is quite abundant for the Aptian–Albian interval, and suggests a currently unrecognised dinosaurian diversity present there ( Dalla Vecchia, 2002; Meyer & Thuring, 2003).
The North American theropod record is dwarfed by an oversampling of latest Cretaceous dinosaur-bearing formations (e.g. Dinosaur Provincial Park, Hell Creek Formation). An increasingly even representation of latitudinally diverse localities from the Cenomanian–Campanian of Utah, Colorado, New Mexico, and Mexico (e.g. Wahweap Formation), may increase the magnitude of the small subsampled diversity drop through the Maastrichtian. Subsampling highlights a latest Jurassic peak in diversity (due to the abundance of remains from the well-sampled Morrison Formation; Foster (2003) ), although Jurassic subsampled diversity never attains that of the Cretaceous highs during the Aptian and Campanian. In contrast to Brusatte et al. (2015), who found no evidence for a progressive Campanian-Maastrichtian decline in North American theropod faunas using similar SQS analyses (implemented in R; see ( Tennant, Mannion & Upchurch, 2016a, 2016b) and ( Alroy, 2010a, 2010c) for comparative discussions), we find a very slight decline that remains constant through publication history, that likely relates to our usage of a slightly different subsampling approach. Aptian subsampled diversity is relatively high due to the more heavily sampled localities from Montana to Texas ( Kirkland et al., 1997; Cifelli et al., 1999; Kirkland & Madsen, 2007).
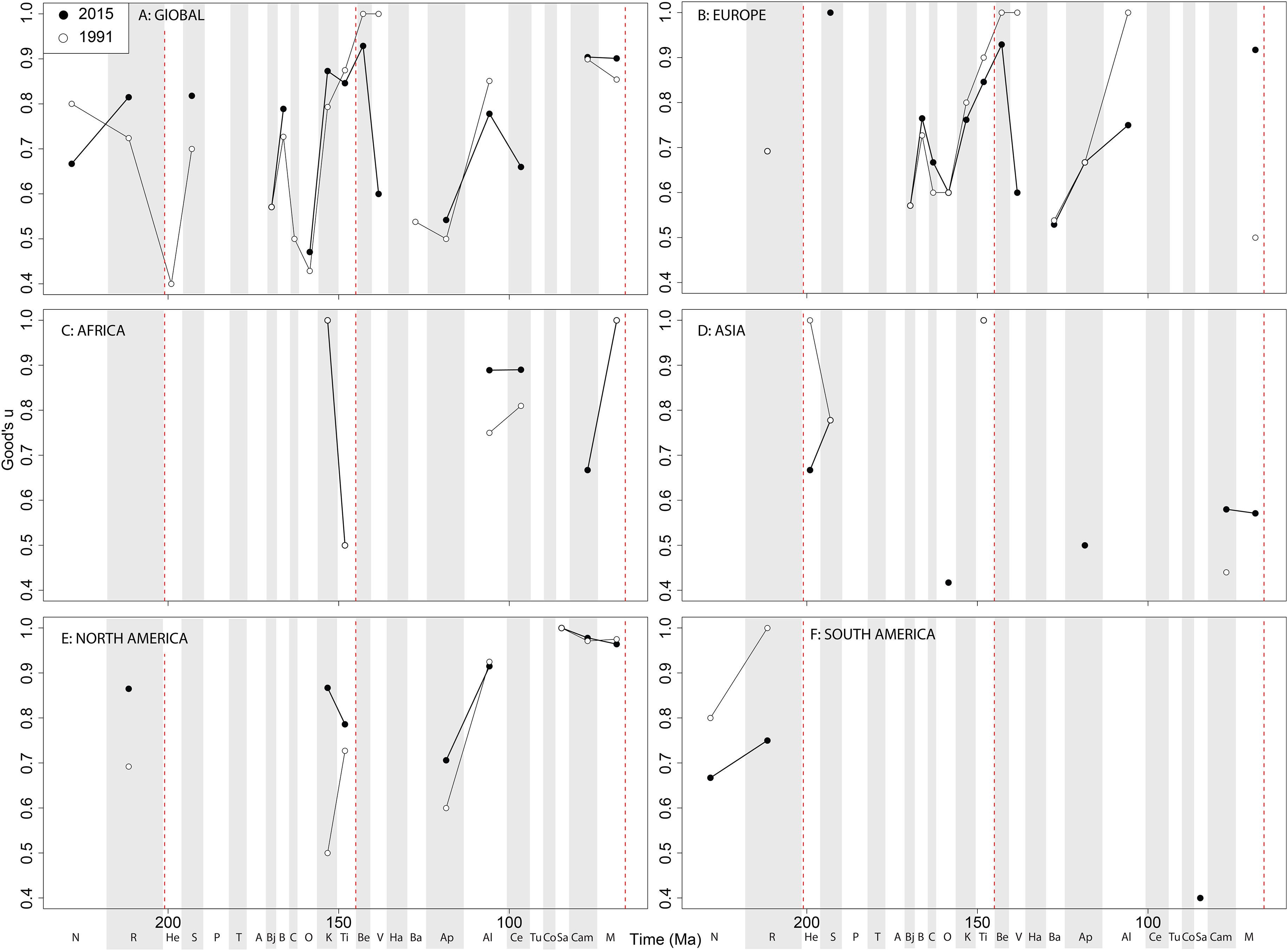
In Africa, there is a Cenomanian radiation ( Fig. 9C View Figure 9 ) mainly due to the multitaxic theropod dominated Kem Kem beds and other Albian–Cenomanian (‘middle’ Cretaceous) equivalents in Northern Africa, but this signal might have been altered by time averaging effects constraining a more temporally diluted diversity in a single unit ( Mannion & Barrett, 2013; Evers et al., 2015; Chiarenza & Cau, 2016). Asian subsampled diversity peaks in the Aptian, Campanian, and Maastrichtian might be explained by a Lagerstätten ‘bonanza’ effect, especially considering the high quality preservation deposits discovered and heavily sampled in the last 20 years (e.g. Liaoning) (Lloyd et al., 2008; Zhou & Wang, 2010; Godefroit et al., 2013; O’Connor & Zhou, 2015; Tennant et al., 2016), although coverage remains only moderate (around 0.5) in each of these intervals ( Fig. 10 View Figure 10 ). Similarly to the pattern in Africa and Asia, South American theropod subsampled diversity stands out compared to other North America and Europe, remaining relatively signal deficient. Despite an increasing rate of discovery of new taxa, which often alter our knowledge of dinosaur phylogeny and biogeography from the ‘middle’ Cretaceous of Patagonia and Brazil ( Novas et al., 2005a, 2013; Novas & Pol, 2005; Canale et al., 2009), coverage remains poor at the stage level, emphasising the need for greater stratigraphic resolution of the theropod-bearing formations here.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.