
Sauropodomorphs
|
publication ID |
https://doi.org/ 10.7717/peerj.4417 |
|
DOI |
https://doi.org/10.5281/zenodo.5614775 |
|
persistent identifier |
https://treatment.plazi.org/id/03FE9007-FFA1-8317-FE21-FBF97FBBD248 |
|
treatment provided by |
Plazi |
|
scientific name |
Sauropodomorphs |
| status |
|
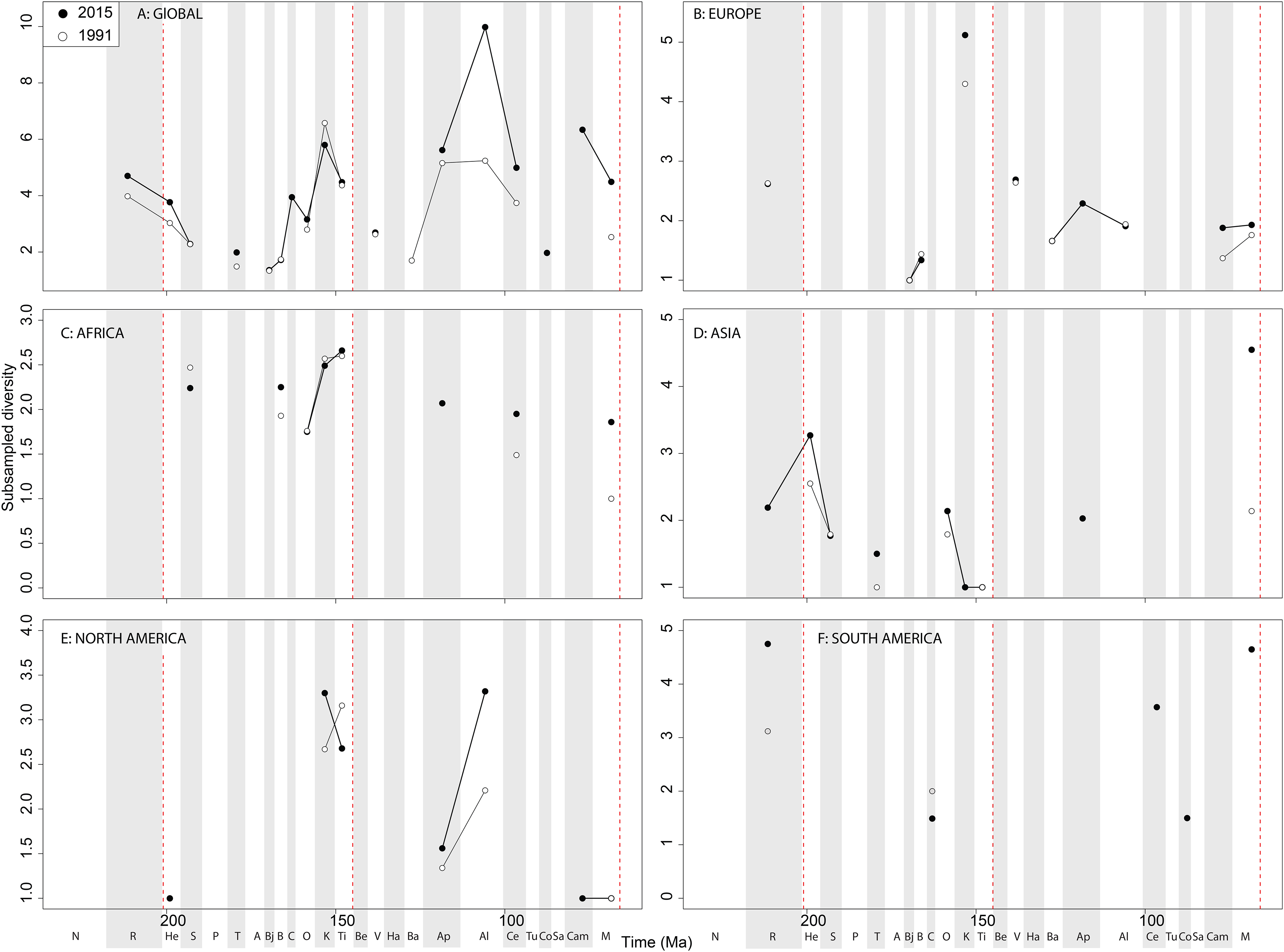
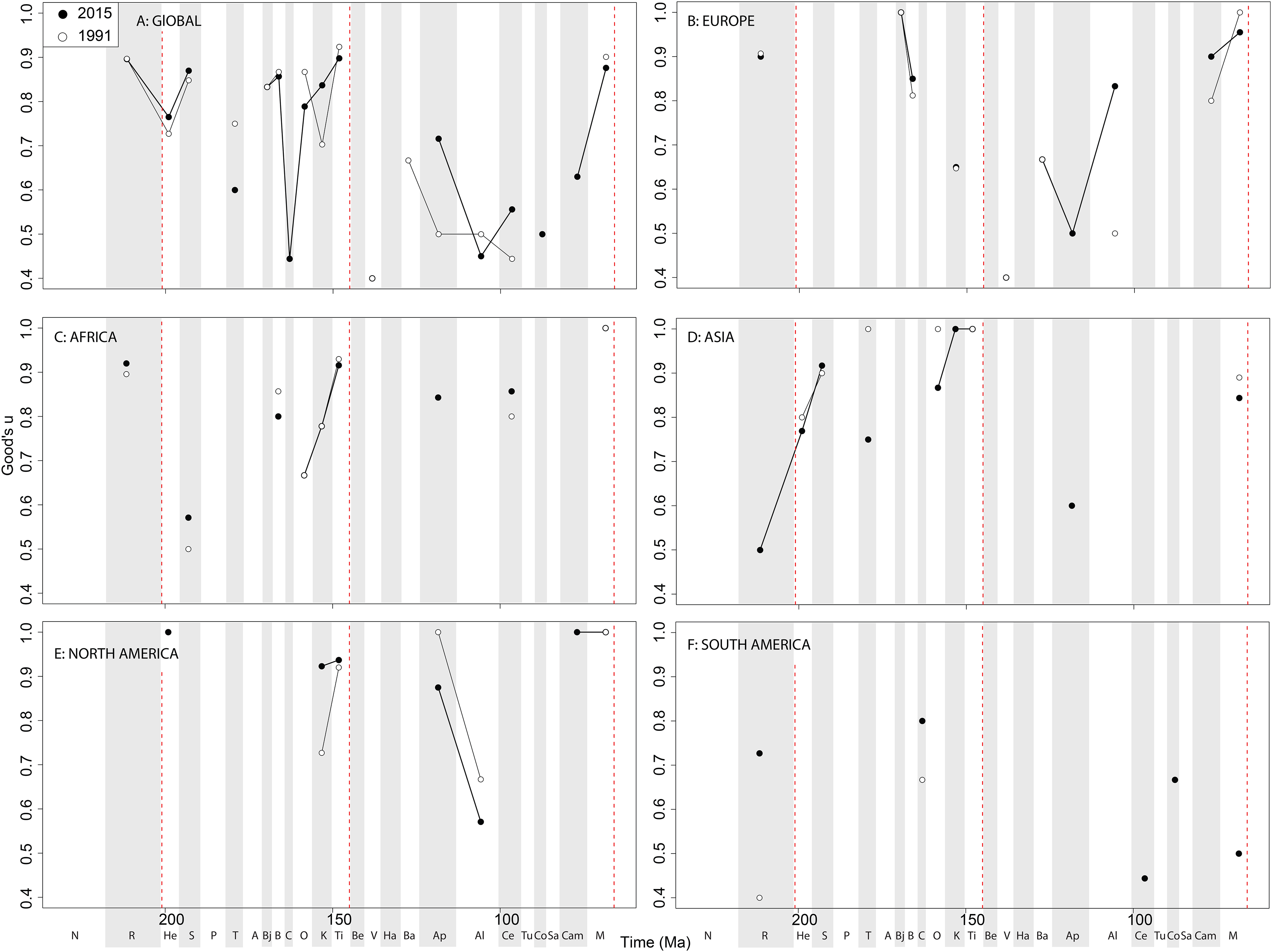
Subsampled diversity patterns of sauropodomorphs share some characteristics of those of theropods and ornithischians, despite having a different fossil record due to taphonomic differences (i.e. larger, more robust skeletons being preferentially preserved in different environmental settings) ( Mannion & Upchurch, 2010, 2011; Dean, Mannion & Butler, 2016). This is compounded by a difficulty in assigning a large number of taxa to specific stage bins, which unfortunately excludes many of them from our analyses (Supplemental Information 1). Differences in diversity patterns between sauropodomorphs and ornithischians have classically been interpreted as being due to exclusive competition between the two main herbivorous dinosaurian subtaxa ( Butler et al., 2009), with an explosive radiation in ornithischians during the Early Cretaceous resulting from the apparent decline in diversity of sauropodomorphs. In fact, the J/K transition represents a major extinction ‘event’ for sauropodomorphs, reflecting the decline of non-neosauropods, diplodocoids and basal macronarians ( Mannion et al., 2013; Tennant, Mannion & Upchurch, 2016b). Sauropodomorph faunas have a low subsampled diversity in the earliest Cretaceous, coupled with a generally poor fossil record ( Mannion & Upchurch, 2010), but at a time when we otherwise see rapid increases in theropod and ornithischian diversity and a prolonged phase of faunal turnover ( Upchurch & Mannion, 2012; Tennant, Mannion & Upchurch, 2016b). Sauropodomorph subsampled diversity levels fluctuate from the ‘middle’ Cretaceous until the final latest Cretaceous radiation, with a possible small decline in the Maastrichtian. This finding is somewhat contrary to that of Sakamoto, Benton & Venditti (2016) who found that their decline was initiated in the Early Cretaceous, and that the diversification of titanosaurs was at an insufficient rate to compensate for the overall loss of sauropodomorph lineages throughout the rest of the Cretaceous. This discrepancy could be due to the differences in datasets used, and that several recently named titanosaurs taxa have yet to be included in published phylogenies, or the distinction between estimation diversity levels against diversification rates. However, we find that sauropodomorphs are at their most diverse during the Albian ( Fig. 12 View Figure 12 ). Sauropodomorphs appear to be overrepresented with respect to what we might expect for almost the entire duration of the Jurassic, whereas the opposite is true for the Cretaceous ( Mannion et al., 2011; Upchurch et al., 2011; Tennant, Mannion & Upchurch, 2016b). The general patterns of ‘global’ subsampled diversity shows a steady increase from Middle to the end of Jurassic with a decline through J/K transition ( Upchurch & Mannion, 2012; Tennant, Mannion & Upchurch, 2016b). The relatively high Late Cretaceous subsampled diversity levels can at least be partially explained by the constant discovery of new titanosaurian taxa, especially from Gondwanan continents ( Vieira et al., 2014; de Jesus Faria et al., 2015; Bandeira et al., 2016; Poropat et al., 2016), and only recently a more appreciated diversity of diplodocoids (e.g. dicraeosaurids, rebbachisaurids) from relatively poorly sampled regions such as Africa ( Mannion & Barrett, 2013; Wilson & Allain, 2015; Ibrahim et al., 2016).
Large-bodied sauropodomorph diversity in the Tithonian is certainly influenced by the intense sampling history of the North American Morrison Formation, where there is an unusually high diversity and cranial disparity of megaherbivores within a relatively resource-poor environment (Button, Rayfield & Barrett, 2014). Here, high diversity remains in spite of our accounting for large collection biases associated with Konzentrat- Lagerstätten ( Alroy, 2010a, 2010c), implying that sauropodomorphs reached their zenith in diversity during the Late Jurassic. Sauropodomorphs appear to be better sampled than theropods and ornithischians in Africa ( Fig. 13C View Figure 13 ), although their records remain largely too inconsistent and patchy record to reveal any major patterns. Asian subsampled diversity is constantly low until the Maastrichtian, where it increases moderately due to a series of recent discoveries from Pakistan and China ( Malkani, 2010; Junchang et al., 2013). However, the Asian Cretaceous sauropodomorph record is otherwise very poorly sampled, especially compared to ornithischians and theropods. This phenomenon could be explained by a taphonomic size bias discriminating against the preservation of largerbodied animals in pre-Late Cretaceous Konservat-Lagerstätten, while they are more present although more rare in the dense bone assemblages from the latest Cretaceous of Mongolia, China, and India ( Kidwell, 2001).
There is a notable subsampled diversity decline in European sauropodomorphs through the J/K transition, as with other dinosaurian groups ( Upchurch & Mannion, 2012; Tennant, Mannion & Upchurch, 2016b). This is distinct from results obtained with other methods (e.g. TRiPS, True Richness estimated using a Poisson Sampling) which do not find any evidence for such a decline ( Starrfelt & Liow, 2016). Subsampling also reveals that sauropodomorph diversity in the latest Cretaceous of Europe was relatively flat. The sauropodomorph record in South America is poor and mostly confined to the Late Cretaceous, with diversity levels rising and resolution improving through publication time as coverage increases and as new taxa get identified from emerging Patagonian and Brazilian deposits ( Novas et al., 2005b; Novas, 2009).
Here, it is worth noting the distinction between global and regional sauropodomorph records. On a global level, our results provide strong evidence for a substantial sauropod subsampled diversity decline from the Campanian to Maastrichtian. However, this decline is not represented in any of the regional sauropodomorph diversity signals. Instead, the ‘global’ signal in the Maastrichtian is comprised of a medley of regional records, which are only continuous with the Campanian record in Europe and North America. Therefore, the ‘global extinction’ of sauropods in the latest Cretaceous is actually due to regionally heterogeneous sampling signals that are summed into a misleading ‘global’ curve. A similar case can be made for the apparently ‘global’ radiation in the Albian, which is primarily a reflection of a well-sampled North American Albian sauropodomorph record ( Fig. 12 View Figure 12 ). Thus, when looking at diversity signals, interpretation of global patterns without considering structural changes on a regional level is not recommended.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.