
Hipposideros boeadii, Bates & Rossiter & Suyanto & Kingston, 2007
|
publication ID |
https://doi.org/ 10.3161/1733-5329(2007)9[13:ANSOHC]2.0.CO;2 |
|
DOI |
https://doi.org/10.5281/zenodo.4323904 |
|
persistent identifier |
https://treatment.plazi.org/id/03FF3346-8E15-FF8B-FD52-EC28FCE7FBE7 |
|
treatment provided by |
Valdenar |
|
scientific name |
Hipposideros boeadii |
| status |
sp. nov. |
Hipposideros boeadii View in CoL sp. nov.
Holotype
Museum Zoologicum Bogoriense M28253 (Field No. TK000803.2), adult ♀, body in alcohol, skull extracted, collected by TK and SJR on 3 August, 2000.
Measurements (in mm) — forearm (FA): 40.7; fifth (5MET), fourth (4MET) and third (3MET) metacarpal lengths
respectively: 28.7, 32.0, 32.3; length of first and second phalanges of third digit (3D1P and 3D2P) respectively: 13.6, 14.0; tail (TAIL): 16.0; foot (HF): 8.7; tibia (TIB): 17.0; ear (E): 18.0; body mass (WGT) 8.5 grams; greatest skull length (GTL): 18.0; condylobasal length (CBL): 15.6; condylo- canine length (CCL): 15.2; zygomatic breadth (ZB): 10.2; breadth of braincase (BB): 7.7; mastoid width (MW): 9.0; rostral width (RW) 5.0; postorbital constriction (PC): 2.9; upper toothrow length (C–M 3): 6.7; breadth of palate (M 3 –M 3): 7.3; lower toothrow length (C–M 3): 7.2; mandible length (M): 11.9; first upper premolar width (P 2 W): 0.50 and length (P 2 L): 0.45; first lower premolar width (P 2 W): 0.45 and length (P 2 L): 0.45; second lower premolar width (P 4 W): 0.80 and length (P 4 L): 0.90.
Type Locality
Lanowulu, Rawa Aopa Watumohai Na- tional Park, South-East Sulawesi (04°22’S, 121°44’E).
Paratypes
Museum Zoologicum Bogoriense M22935 (Field No. TK 000804.2) adult ♁ (4 August, 2000); M22937 View Materials (Field No. TK000809.2) adult ♁ (9 August, 2000); M22934 (Field No. TK 000804.1) adult ♀ (4 August, 2000); all bodies in alcohol with skulls extracted collected at Lanowulu, Rawa Aopa Watumo-hai National Park GoogleMaps , South-East GoogleMaps Sulawesi, (04°22’S, 121°44’E).
Diagnosis
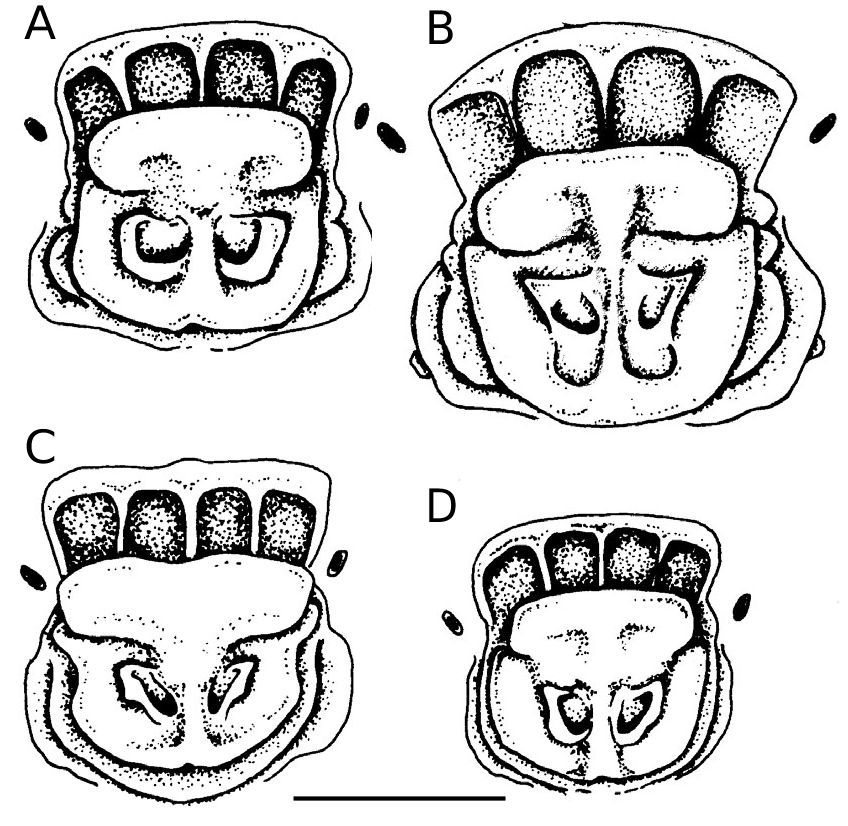
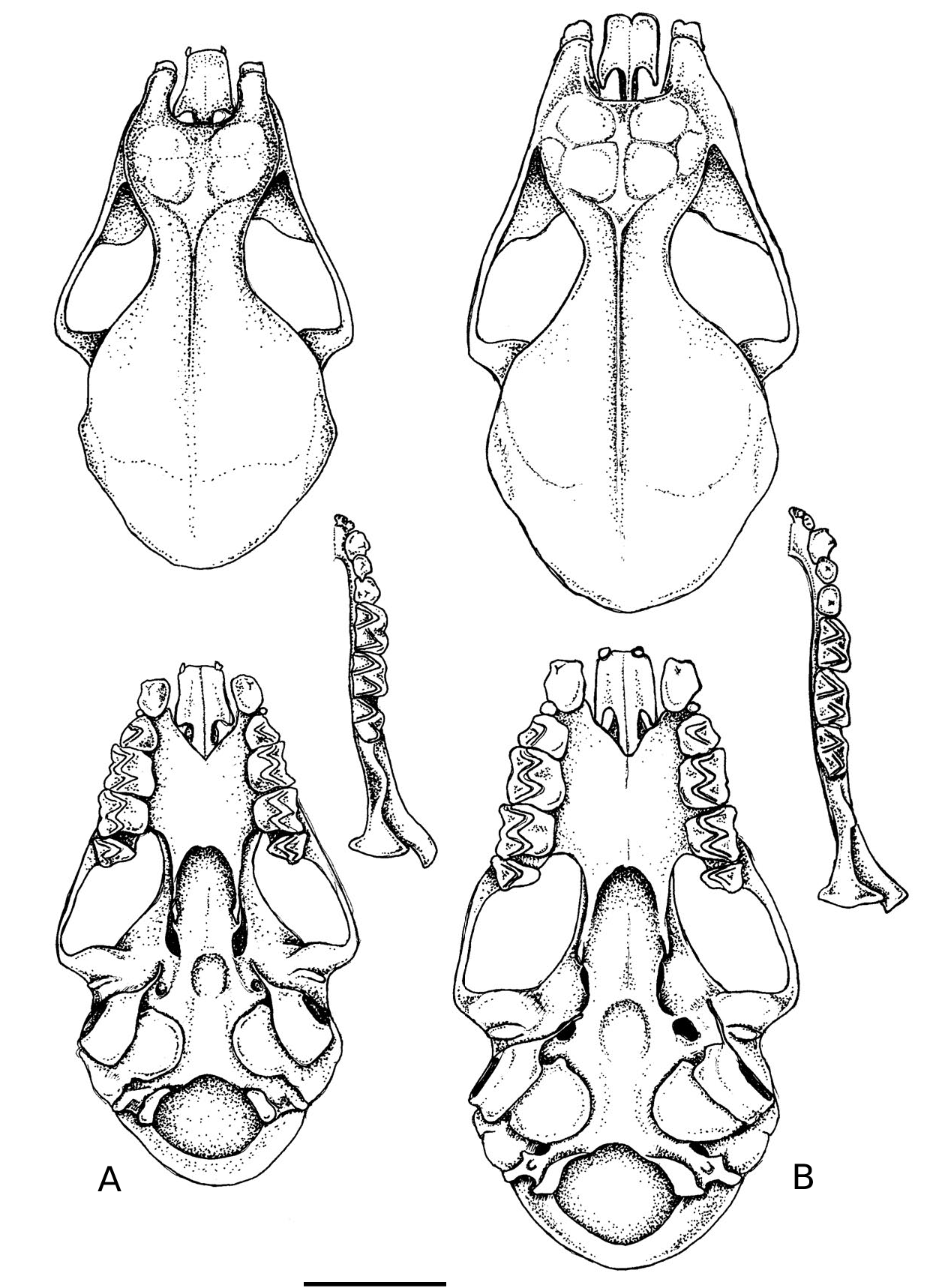
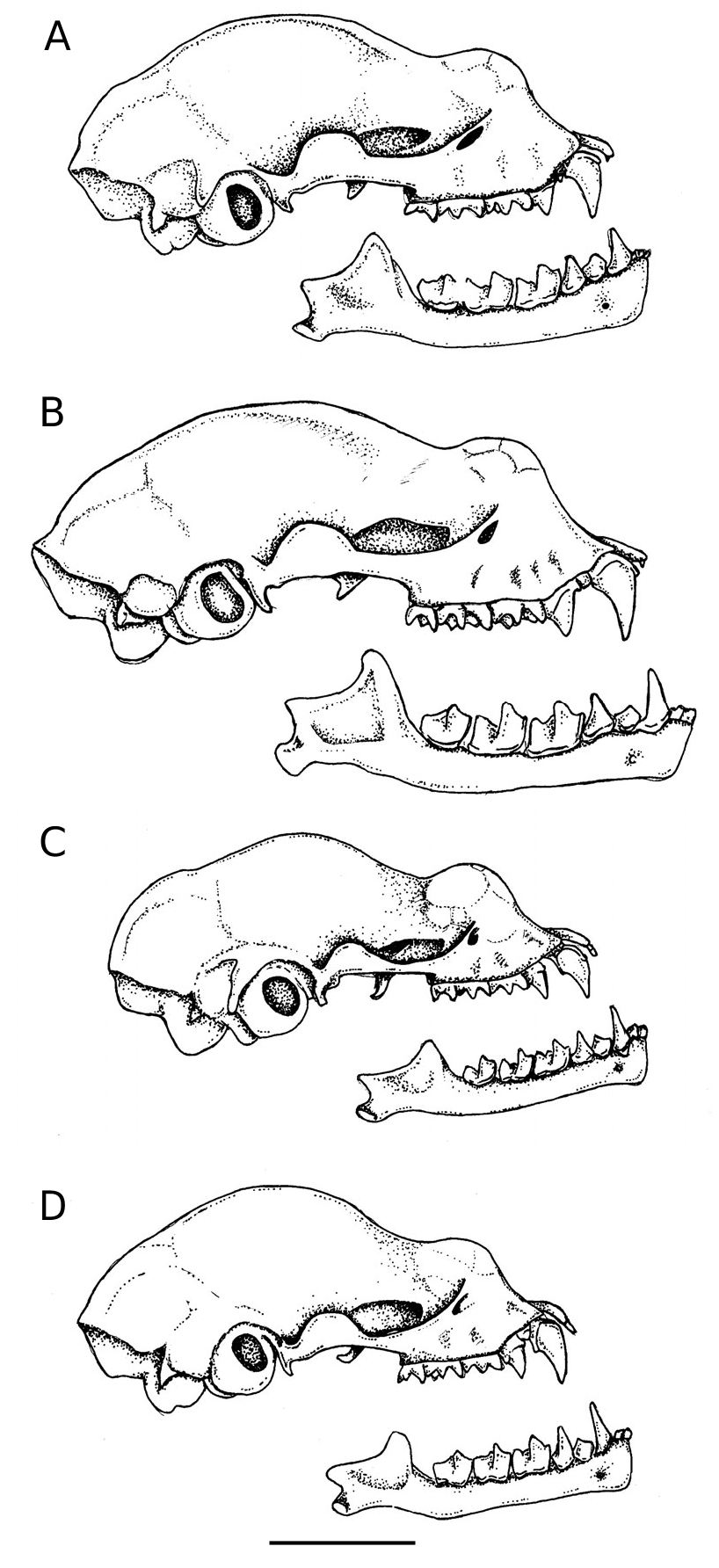
This is a medium-small hipposiderid bat with a forearm length of 40.5–42.7 mm tak- en from specimens preserved in alcohol ( Table 1 View TABLE ). The ears have rounded tips. The noseleaf has two supplementary leaflets; the inner is short; the outer virtually reach- es the midline anteriorly ( Figs. 1 View FIG and 2A View FIG ). In the wing, the fifth metacarpal is be- tween 70–75% of forearm length and the first phalanx of the third digit is short (13.6–13.9 mm). The tail is also very short (16.0–21.0 mm). The skull, with a condylo- canine length of 15.2–15.5 mm ( Table 2), is large in comparison to forearm length. The zygomata are robust, broad and consider- ably exceed mastoid width ( Fig. 3A View FIG ). There is a well developed dorsal projection on the jugal bone ( Fig. 4A View FIG ). The rostrum is moder- ately inflated with a slight depression in the midline and with the anterior and postero- medial rostral chambers about equally in- flated ( Fig. 5A View FIG ).
Description
With GoogleMaps a body mass of 6.8–8.5 grams and a forearm length of 40.2–43.1 mm (live and alcohol preserved specimen data combined, n = 15), this is a medium-small Hippo- sideros ( Table 1 View TABLE ). Pelage colour is uniform- ly reddish-brown. The wing and tail membranes are dark brown to blackish, as are the ears, noseleaf and lateral leaflets. The tail is very short, on average slightly exceeding the tibiae in length. In the wing (based on four alcohol preserved specimens), the fifth metarcarpal is between 5.7–10.3% shorter than the fourth ([4MET-5MET]/4MET) and is 70–75% of forearm
length. The first phalanx of the third digit is short, between 43.2 and 44.4% of third metacarpal length. The ear is 16.5–18.0 mm in length, with a rounded tip and a slight concavity on the posterior border. The an- titragus has a fold on its upper border ( Fig. 1 View FIG ); when flattened, the fold appears as a concavity which divides the antitragus in to two parts. In the noseleaf ( Fig. 2A View FIG ), the internarial septum is slightly swollen anteri- orly and the narial lappets have a smoothly rounded outline. The intermediate leaf is simple and slightly inflated medially. It is subequal in width to the anterior leaf and distinctly narrower than the posterior one. Although its medial part is essentially naked, it has a number of hairs of various lengths on the lateral lappets. The anterior noseleaf is a simple horseshoe shape with a small notch in the mid-part of the anterior border. The upper part of the anterior leaf
is attached to the intermediate leaf. The posterior leaf has an evenly rounded upper margin and four well-defined concavities. A well-developed frontal sac is present in males on the forehead behind the posterior leaf; it is not present in female specimens. Small projections on the forehead above each eye are present in both males and females.
There are two supplementary lateral leaflets ( Figs. 1 View FIG and 2A View FIG ). The first is about half the length of the anterior leaf and its upper part is attached to the posterior leaf. The second supplementary leaflet nearly reaches the midline anteriorly; there is an indication of the leaflet as far as the midline, although at this point the leaflet is attached completely to the muzzle. Its upper part stops about 1.3 mm below the eye and is not attached to the posterior leaf. Both supplementary leaflets are lightly covered with bristly hairs of different lengths (these have been omitted from Fig. 2 View FIG ).
The skull has a condylo-canine length of 15.2–15.5 mm ( Table 2). The zygomata are robust and broad, considerably exceed- ing mastoid width ( Fig. 3A View FIG ). There is a well developed dorsal projection on the jugal bone; the upper border of which is evenly rounded, almost semi-circular in outline ( Fig. 4A View FIG ). The anterorbital foramen, which has a thin narrow bar of bone above it, is long, narrow and elliptical in shape. The rostrum is moderately inflated with a slight depression in the midline; the anterior chambers are about equally inflated to the postero-median chambers; the postero- lateral chambers are not greatly inflated ( Fig. 5A View FIG ).
The sagittal crest is well defined ante- riorly. It bifurcates into the supraorbital ridges which are indicated but not devel- oped. The postorbital constriction, although narrower than the rostrum, is not conspicu- ously constricted. The mastoid is inflated. The supraoccipital and foramen magnum
are directed downwards and only slightly backwards. The palate is emarginated ante- riorly to a point corresponding to the ante- rior border of the first upper molar (M 1) ( Fig. 3A View FIG ). Posteriorly, it is emarginated to the mid-point of the third upper molar (M 3). The anterior border of the mesopterygo- id space is essentially U-shaped and there is no bony projection (spicule) from the midpoint of the posterior palate. There is a shallow sphenoidal depression. The width of each cochlea exceeds the width between the cochleae. Each tympanic bulla covers less than half of its respective cochlea. Each half mandible has a robust horizontal ramus and a well developed coronoid process, which exceeds the lower canine in height ( Fig. 4A View FIG ).
Upper toothrow length is 6.7–6.9 mm. The upper incisor (I 2) is very small and
wedge shaped. The upper canine (C 1) is of moderate size, with the faintest indication of a groove on its antero-lateral surface and
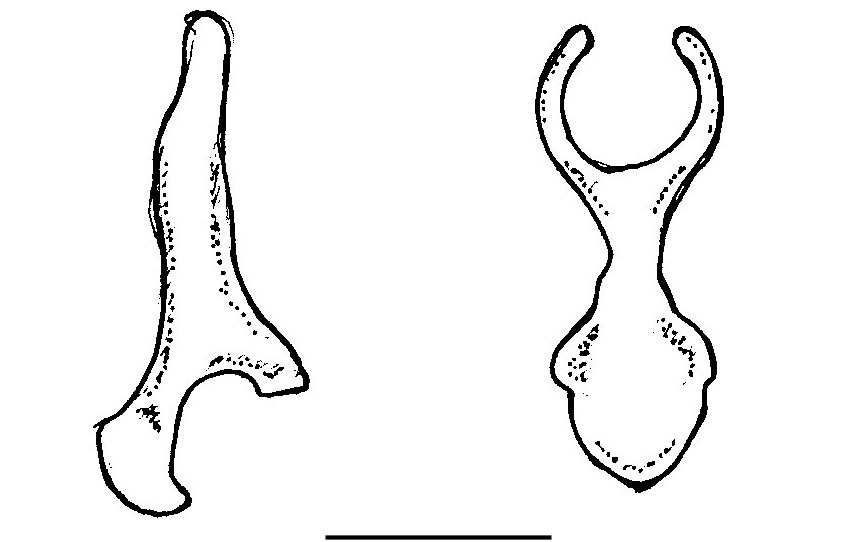
with an ill-defined cingulum. The first up- per premolar (P 2) is minute and displaced externally ( Fig. 3A View FIG ). The canine (C 1) and second upper premolar (P 4) are almost in contact. All the cheekteeth are robust. The first (M 1) and second upper molars (M 2) have the typical W-shaped cusp pattern with parastyle and metastyle more developed than the mesostyle; the paracone and metacone are about equal in size. The third upper molar (M 3) is reduced; it includes a parastyle and mesostyle and two commissures, with a vestige of a third commissure posteriorly. Both the lower incisors are tricuspidate. The second (I 2) is partly situated behind the first (I 1). The lower canine (C 1) is not especially tall ( Fig. 4A View FIG ). The first lower premolar (P 2) is about two thirds the crown area of the second (P 4). The second lower molar (M 2) very slightly exceeds the first (M 1) in crown area; in the third (M 3), the talonid is reduced in size. The baculum is small (1.9 mm in to- tal length) with a horseshoe shaped tip, short shaft and a broad base, which is hollowed underneath and with a small keel ventrally ( Fig. 6 View FIG ).
Echolocation
The echolocation calls of H. boeadii are typical of the Hipposideridae , with most energy in a constant frequency (CF) component of approximately 2.5–4.0 ms, terminating in a steep frequency modulated tail (1.0–1.5 ms duration). Mean value for the frequency of maximum energy of the CF portion of the call is 116.6 kHz, with a range of 114.2–118.3 kHz (n = 13).
Etymology
Hipposideros boeadii is named after the Indonesian zoologist Boeadi, in rec- ognition of his long-standing contribu- tion to bat research in Indonesia. Its pro- posed English name is ‘Boeadi’s roundleaf bat’.
DISCUSSION
Comparison with Other Taxa
Hipposideros boeadii is one of a number of Hipposideros taxa described from south- ern and South-East Asia, which have two supplementary leaflets in the noseleaf. In H. boeadii , the inner leaflet is shorter than the outer. This distinguishes it from the two geographically widespread species, H. ga- leritus Cantor, 1846 (described from Pe- nang, Malaysia) and H. cervinus Gould, 1854 (described from Queensland, Austral- ia), both of which were also collected from Buton (South-East Sulawesi) during 2000. In both these latter taxa, the inner leaflet ex- ceeds the outer in length ( Fig. 2C and 2D View FIG ). Other characters that distinguish H. boeadii
from H. galeritus and its associated taxa [brachyotus Dobson, 1874 (described from Central India), insolens Lyon, 1911 (southeastern Kalimantan), and longicaudus Peters, 1861 (Java)] are its shorter forearm length, larger skull, broader zygomata, more robust dentition and the absence of a greatly inflated postero-lateral chamber either side of the rostrum [ Tables 1 View TABLE and 2, Figs. 4 View FIG and 5 View FIG — for further details of these latter taxa, including measurements, see Hill (1963) and Jenkins and Hill (1981)]. In the noseleaf, the intermediate leaf is nar- rower than the posterior, whereas in H. galeritus and its associated forms, it is broad- er ( Fig. 2 View FIG ).
In comparison to H. cervinus , the skull of H. boeadii is longer relative to forearm length, the zygomata are broader, the cheek-
teeth less robust relative to skull size and the postero-lateral chambers of the rostrum not greatly inflated ( Fig. 5 View FIG ). Comparison was made with a further five taxa that have been referred to H. cervinus (Jenkins and Hill, 1981). These are labuanensis Tomes, 1859 (described from Labuan Island and Sarawak), batchianus Matschie, 1901 (Bat- jan Island, northern Moluccas), schneider- si (misprint for schneideri) Thomas, 1904 (Sukaranda, Sumatra), misorensis Peters, 1906 (Biak Island, Papua Province) and ce- lebensis Sody, 1936 (southern Sulawesi).
The noseleaf morphology of the holo- type of labuanensis is difficult to determine since it is a dry skin. The forearm is longer (47.8 mm) but the dentition of the damaged skull is similar in size to that of H. boeadii . However, unlike H. boeadii , the lateral compartments of the rostrum in labuanen- sis are inflated (typical of cervinus ) where- as those of H. boeadii are not enlarged. The taxon schneidersi was included as a syn- onym of H. c. labuanensis by Jenkins and Hill (1981). Although it is typical of H. cer- vinus in the morphology of the noseleaf and rostrum, it differs in having a more robust skull with significantly broader zygomata (116.1% of mastoid width), a character shared with H. boeadii . These differences in the skull suggest that Tate (1941) may have been correct in treating H. schneidersi
as a distinct species. The taxon batchianus is poorly known ( Hill, 1963). It was exam- ined by Tate (1941) who included it in the galeritus group but was subsequently re- ferred to H. cervinus (Jenkins and Hill, 1981 ). Its forearm, tail and upper toothrow lengths of 47.5 mm, 25 mm and 6.4 mm, re- spectively, suggest that it is a typical H. cer- vinus. The taxon misorensis is also poor- ly known, with all the current informa-
tion based on a plate published by Peters (1906). However, according to Jenkins and Hill (1981), the noseleaf closely resem- bles that of H. cervinus . Jenkins and Hill (1981) included celebensis as a synonym of H. c. cervinus and the two taxa appear to be indistinguishable.
In addition to H. cervinus and H. galeritus, there are six species which not only have two supplementary leaflets but also have the inner leaflet shorter than the outer, a character shared with H. boeadii . Four of these, muscinus Thomas and Doria, 1886 (described from Fly River, Papua New Guinea), wollastoni Thomas, 1913 (Uta- kawa River, Irian Jaya), semoni Matschie, 1903 (Queensland, Australia), and stenotis Thomas, 1913 (Northern Territory, Australia) were included in the cyclops group by Hill (1963). Two others, H. corynophyllus Hill, 1985 (West Sepik, Papua New Guinea) and H. edwardshilli Flannery and Col- gan 1993 (Bewani Mountains, Papua New Guinea) were added subsequently.
In comparison to H. boeadii , H. muscinus has larger, more inflated anterior chambers of the rostrum, larger cochleae, which are more closely situated together, and with the first upper premolar (P 2) less reduced and situated in the toothrow. The posterior leaf of the noseleaf has a small tubercle which is not present in H. boeadii . The taxon wollastoni has a distinctive noseleaf. The intermediate leaf has a large projecting median eminence and the upper edge of the leaf has two lateral elevations. In addition,
there is a transverse supplementary struc- ture which is developed from the posterior face of the posterior leaf and is almost equal in height to the posterior leaf. There are also significant differences in the skull, most notably in the greatly expanded cochleae. Hipposideros semoni , H. stenotis , H. cory- nophyllus and H. edwardshilli can be dis- tinguished externally from H. boeadii by well developed club-shaped projections on the intermediate and posterior leaves of the noseleaf. In the skulls of H. semoni and H. stenotis , the cochleae are enormous and almost touching; the posterior commissure of M 3 is not greatly reduced ( Hill, 1963).
The taxon H. crumeniferus (Lesueur and Petit, 1807) (described from Timor) was listed as incertae sedis by Corbet and Hill (1992), pending the rediscovery of the hol- otype or the collection of topotypes. Pre- viously, Tate (1941) suggested that it might have affinities to H. papua (Thomas and Doria, 1886) (Biak Island, Papua Province). However, based on the plate in the Atlas of Lesueur and Petit (1807), Hill (1963) noted that unlike H. papua the posterior (second) leaflet does not extend beneath the anterior part of the horseshoe; this would also dis- tinguish it from H. boeadii . Hill (1963) sug- gested that it was referable to H. cervinus or a closely related form.
Hipposideros boeadii appears most sim- ilar to H. papua . This taxon has not been recorded from Sulawesi but is known from the Molucca Islands and western New Guinea ( Flannery, 1995). With a forearm length of 49.6–53.6 mm and a condylocanine length of 16.7–17.6 mm, H. papua considerably exceeds H. boeadii (40.5–42.7 mm and 15.2–15.5 mm respectively) in size ( Tables 1 View TABLE and 2). In addition, in H. papua there is usually (but not always) a very small third supplementary leaflet to the noseleaf (Hill and Rozendaal, 1989); this was not detected in the four specimens of H. boeadii currently known from Sulawesi. The wing morphology also appears to differ slightly between the two species (although current sample sizes are small). In H. boe- adii, the fifth metacarpal is relatively longer, 70.5–74.8% (n = 4) of forearm length; in H. papua , the comparable figure is 65.1– 65.6% (n = 2). The skull of H. bo- eadii is also long in comparison to fore- arm length. In H. boeadii , it is 36.2–38.2% (n = 4) (CCL/FA × 100), whereas in H. pa- pua, it is 33.2–34.3% (n = 4). The morphol- ogy of the skulls and dentition is otherwise similar between the two species with the possible exception of the first upper premo- lar (P 2), which is minute and extruded from the toothrow in H. boeadii and is larger and situated in the toothrow in H. papua (Fig. 3), although further specimens may show this to be an individually variable character.
Hipposideros boeadii is morphologically discrete from H. speoris ( Schneider, 1800) (described from Tamil Nadu, India) and H. larvatus ( Horsfield, 1823) (Java) , two species with which H. papua is sometimes linked (for full discussion see below). Unlike H. boeadii , both H. speoris and H. larvatus have three supplementary leaf- lets, with the first leaflet exceeding or about equal in length to the second. Hipposide- ros boeadii also differs in its rostral mor- phology and smaller size. The forearm and condylo-canine lengths of these latter two taxa are respectively: FA: 45.6–54.0 mm and 61.2–64.8 mm; CCL: 15.9–17.5 mm and 20.8–21.4 mm (measurements from Bates and Harrison, 1997).
All individuals of H. boeadii were cap- tured in lowland rainforest between one and four kilometres from the adjacent sa- vannah. The forest was disturbed, and veg- etation near to the edge was characterised by a high incidence of bamboo and rattan. Other bat species recorded at this site, in
order of relative abundance, were Kerivoula hardwickii, Rhinolophus euryotis , Murina florium, K. papillosa , H. cervinus , R. cele- bensis, Phoniscus jagorii , and an uniden- tified Hipposideros species morphological- ly close to H. ater .
Distribution
To date, H. boeadii is only known from the type locality. Future studies may show it to be more widespread elsewhere in Indo- nesia and beyond.
Reproductive Biology
The nipples and pubic nipples of eight female H. boeadii were examined. Four in- dividuals were parous (including the two fe- male voucher specimens) and four nul- liparous. Pregnancy was not detected by either external abdominal palpation of the released individuals, or dissection of the two female specimens. However, vaginal plugs were found in three individuals (in one of the parous specimens and in two of the nulliparous females released at the site), indicating that mating had recently occurred.
Taxonomic Notes
Based on external and cranial morpholo- gy H. boeadii appears to have strong affini- ties to H. papua , a species whose taxonom- ic position is uncertain. Tate (1941) consid- ered H. papua to be a ‘perplexing species’ which ‘may have been derived from near the base of the speoris and galeritus (cervi- nus) stems’. According to Hill (1963), the ears and cranial characters of H. papua sug- gest affinity with the bicolor group, where- as its specialised noseleaf indicates no close connection with any other members. Some affinity to the cyclops group is suggested by the extension of the second leaflet beneath the anterior leaf, a feature otherwise pecu-
liar to cyclops Temminck, 1853 (described from Ghana, West Africa) and its allies, but H. papua differs from these in its broad ears, the presence of an incipient third leaflet, unenlarged bullae and convergent toothrows. Hill (1963) concluded that H. papua should be regarded as a species of remote origin within the bicolor group, as- sociated probably with the line leading to the muscinus group and possibly with that leading to the speoris group. This view was followed by Corbet and Hill (1992) and Koopman (1994), who also assigned H. papua to the bicolor group, which in- cluded galeritus but omitted speoris . How- ever, it was not followed by Hand and Kirsch (1998) or Bogdanowicz and Owen (1998). The former found no close affinities between H. papua and other members of the genus, whilst the latter tentatively suggested that H. papua was most closely related to H. larvatus , which is a member of the spe- oris group (sensu Hill, 1963). Based on preliminary phylogenetic comparisons, H. boeadii appears to be more closely relat- ed to H. diadema (E. Geoffroy, 1813) (de- scribed from Timor) and H. larvatus than to H. galeritus and H. cervinus (S. Murray, un- published data).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |