
Supramontana argentina, Negrete, Lisandro, Leal-Zanchet, Ana Maria & Brusa, Francisco, 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3753.2.7 |
|
publication LSID |
lsid:zoobank.org:pub:74D353B7-4D92-4674-938C-B7A46BD5E831 |
|
DOI |
https://doi.org/10.5281/zenodo.5614761 |
|
persistent identifier |
https://treatment.plazi.org/id/03FF8795-9453-4E20-299C-2A5CFE7393DB |
|
treatment provided by |
Plazi |
|
scientific name |
Supramontana argentina |
| status |
sp. nov. |
Supramontana argentina View in CoL sp. nov.
Type series. Holotype: MLP 6699. Argentina , Misiones, INP, 13.IV.2013, L. Negrete, coll.; anterior region 1: transverse sections on 17 slides; anterior region 2: sagittal sections on 30 slides; pre-pharyngeal region: transverse sections on 6 slides; pharynx: sagittal sections on 18 slides; copulatory apparatus: sagittal sections on 35 slides.
Paratypes: MLP 6698. Argentina , Misiones, RVSU, 05.V.2008, L. Negrete, coll.; anterior and pre-pharyngeal regions: transverse sections on 17 slides; pharynx and copulatory apparatus: sagittal sections on 7 slides. MLP 6476. Argentina , Misiones, RVSU, 21.VIII.2009, L. Negrete, coll.; anterior region: sagittal sections on 14 slides; pre-pharyngeal region: transverse sections on 3 slides; pharynx and copulatory apparatus: sagittal sections on 28 slides. MLP 6475. Argentina , Misiones, CAMB, 20.III.2010, L. Negrete, coll.; anterior region: sagittal sections on 15 slides; pre-pharyngeal region: transverse sections on 3 slides; pharynx and copulatory apparatus: sagittal sections on 30 slides.
Diagnosis. Species of Supramontana with dorsal surface with a yellowish median band, bordered by two black para-median stripes, and two dark grey lateral bands; ventral surface whitish; ventral cephalic retractor muscle almost circular in cross section; prostatic vesicle extrabulbar, tubular and very long; penis papilla conical and blunt with a sinuous ejaculatory duct.
Type locality. Iguazú National Park, in native subtropical forest.
Habitat. The flatworms were found on the ground under fallen logs and when they were crawling at night, in environments with native vegetation.
Etymology. The specific name honours Argentina , the country where the species occurs.
External morphology. The body is elongate, with parallel margins, slightly wider at the pharyngeal level, narrowing towards the ends. The anterior tip is rounded while the posterior tip progressively narrows ending sharpedged. The colour pattern of the dorsal surface is dark grey, constituting two lateral bands (2/3 of body width) and black para-median stripes (~1/10 of body width each one). In addition, there is a yellowish median band (1/5 of body width) that extends along the body except for a small portion of the anterior region. Body margins are whitish as the ventral surface ( Figures 1 View FIGURE 1 , 2 View FIGURE 2 ). The eyes present clear halos. They surround the anterior tip uniserially and extend toward both margins of the body for 2-4 mm. Further back, they spread to both sides of the dorsal surface, forming various rows and extending over para-median stripes ( Figure 2 View FIGURE 2 ). They are numerous until the pharyngeal level, and posteriorly maintain the same position but in less quantity. They become marginal at the level of the copulatory apparatus, reaching the posterior end.
The largest specimen reached 60 mm in length in vivo. After fixation, body length ranged from 17 mm to 45 mm, maximum width 1.6–2.9 mm, and maximum height 0.6–1.7 mm. The mouth was located at a distance of 54– 65% from the anterior tip, and the gonopore at 76–83% ( Table 1 View TABLE 1 ).
Internal morphology. Cephalic region. The position and arrangement of the cutaneous muscle layers differ from those in the pre-pharyngeal region (see below) in that the ventral longitudinal cutaneous muscle layer is sunk into the parenchyma and forms a retractor muscle that is almost circular in cross section ( Figure 3 View FIGURES 3 – 5 ). This retractor muscle diminishes in thickness towards the anterior end until disappear next to the tip. Backward it becomes progressively wider and finally form two layers of longitudinal muscles that occupy ventrally the whole width of body, separated each other by a nerve plexus ( Figures 4, 5 View FIGURES 3 – 5 ). Circular, oblique and dorsal longitudinal layers of the cutaneous musculature are thinner than in the pre-pharyngeal region. The parenchymatic musculature is arranged as supra-intestinal and sub-intestinal transverse layers, thinner as the intestine becomes less apparent towards the tip. Dorso-ventral fibres are scarce. There is a sub-neural transverse parenchymatic muscle layer, located just below the nerve plate and above the retractor muscle ( Figure 3 View FIGURES 3 – 5 ). Sensory pits, as a simple invagination of ventral epidermis (25–45 µm deep), surround the cephalic region ( Figure 3 View FIGURES 3 – 5 ).
Numerous rhabditogen cells with xanthophil secretion (rhammites), cells with coarse granular erythrophil secretion (~2.5 Μm), and cells with strong cyanophil, coarse granular secretion (~2 Μm) open through anterior tip and dorsal epidermis. The narrow creeping sole—between 14% and 51% of body width on first 0.7 mm of body length—receives abundant coarse granular strong cyanophil secretion (~2.5 Μm). In addition, less frequent rhabditogen cells (small rhabdites) and erythrophil cells with fine granular secretion (about 1 Μm) open through the creeping sole. The remaining ventral surface presents abundant rhabditogen cells with xanthophil secretion (rhammites), numerous cells with strong cyanophil, coarse granular secretion (~2 Μm), and less frequent cells with ill-defined, coarse granular secretion (~3 Μm).
Epidermis and musculature at pre-pharyngeal region. Five types of secretory cells open through dorsal epidermis (15–25 Μm high) and body margins: (1) numerous cells with coarse granular erythrophil secretion (~3 Μm), (2) abundant rhabditogen cells with xanthophil secretion (rhammites), and less frequent (3) erythrophil cells with fine granular secretion (<1 Μm), (4) weakly cyanophil cells with amorphous secretion and (5) cells with strong cyanophil, coarse granular secretion (~2 Μm). There is no glandular margin ( Figures 4, 5 View FIGURES 3 – 5 ). Ventral epidermis (22.5– 30 Μm high) is ciliated on creeping sole, which is approximately 85-90% of body width ( Figure 4 View FIGURES 3 – 5 ). Four types of glands discharge their secretion through the creeping sole: abundant cells with coarse granular, strong cyanophil secretion (~3 Μm), rhabditogen cells (with small rhabdites) and erythrophil cells of two types, one with fine (<1 Μm) and other with coarse (~2 Μm) granular secretion.
The cutaneous musculature consists of three layers: a thin external subepithelial layer of circular muscles, followed by a diagonal layer with decussate fibres, and a thicker longitudinal layer arranged in bundles. Ventrally, the longitudinal layer is thicker than dorsally, being subdivided into two portions, one just below the diagonal fibres, and the innermost one sunk into the parenchyma ( Figures 4, 5 View FIGURES 3 – 5 , Table 2 View TABLE 2 ). The CMI varies between 14% and 21% ( Table 2 View TABLE 2 ).
The parenchymatic musculature is composed by four layers: a dorsal subcutaneous with oblique fibres, a supra-intestinal and a sub-intestinal transverse layers, and a sub-neural transverse layer ( Figures 4, 5 View FIGURES 3 – 5 ). Also, dorsoventral fibres are arranged among intestinal branches. The thickness of the parenchymatic musculature represents 6–11% of body height ( Table 2 View TABLE 2 ).
Digestive system. The pharynx (1.4–2.75 mm in length) is bell-shaped, with the dorsal insertion at mouth level or a bit posteriorly displaced ( Figure 6 View FIGURE 6 ). The mouth is situated in the anterior third of the pharyngeal pouch (1.45– 3.75 mm in length). The epithelial lining of the outer surface of the pharynx is cuboidal and ciliated. The outer pharyngeal musculature is arranged in two layers: a thin longitudinal subepithelial layer (2.5–5 Μm thick) followed by a subjacent circular layer (25–27.5 Μm thick). The epithelium of the pharyngeal lumen is columnar, ciliated, and strongly erythrophil. The inner pharyngeal musculature consists of a circular subepithelial layer (25–50 Μm thick) followed by a subjacent longitudinal one (10–37.5 Μm thick). Six secretory cell types, three of them with cell necks close to the epithelium of the pharyngeal lumen, open through this epithelium: cells with fine granular erythrophil secretion (<1 Μm); cells with fine granular cyanophil secretion (1 Μm), and cells with fine granular weak cyanophil secretion (1 Μm). Other three types of glands show cell necks close to the outer epithelium of the pharynx, opening through the pharyngeal tip: cells with cyanophil amorphous secretion; cells with coarse granular erythrophil secretion (~3 Μm); and cells with fine granular weak erythrophil secretion (<1 Μm). The oesophagus is absent ( Figure 6 View FIGURE 6 ).
Male reproductive system. The testes are arranged in two or three rows on each side of the body, dorsal to the intestinal branches and just below the supra-intestinal parenchymatic muscle layer ( Figures 4, 5 View FIGURES 3 – 5 ). They are prepharyngeal, appearing behind the ovaries and extending to near the level of the ventral pharyngeal insertion. They are located at a distance between 18–23% and 48–61% of the body length from anterior end ( Table 3 View TABLE 3 ). Testes are rounded or ovoid, occupying 9–23% of the height of body ( Table 3 View TABLE 3 ). The sperm ducts are located above the subintestinal parenchymatic muscle layer or among its fibres, being dorsal to the ovovitelline ducts, laterally displaced ( Figures 4, 5 View FIGURES 3 – 5 ). The sperm ducts expand distally with their lumen full of spermatozoa. They bend toward the dorsum and slightly backward and open laterally into the proximal portion of the prostatic vesicle ( Figures 7, 8 View FIGURES 7 – 8 ). The extrabulbar prostatic vesicle is a long unpaired tube. Its proximal portion is almost straight and dilated; the rest of the prostatic vesicle is tortuous. The ejaculatory duct runs almost centrally in sinuous way through the penis papilla. The latter occupies the entire length of the male atrium, being conical and blunt in shape, with its insertions at the same transverse level ( Figures 7–10 View FIGURES 7 – 8 View FIGURES 9 – 13 ). The walls of the male atrium are not folded.
The prostatic vesicle is lined with ciliated columnar epithelium followed by a muscular layer with intermingled circular and less abundant longitudinal fibres (10–30 Μm thick) ( Figures 7–9 View FIGURES 7 – 8 View FIGURES 9 – 13 ). The cell necks of numerous secretory cells with fine granular erythrophil secretion (<1 Μm) and less abundant cyanophil cells with coarse granular secretion (~2 Μm) open through the epithelium of the prostatic vesicle. The cell bodies of these glands are found in the parenchyma, mainly around the prostatic vesicle ( Figures 7–9 View FIGURES 7 – 8 View FIGURES 9 – 13 ). The ejaculatory duct is lined with ciliated columnar epithelium, followed by a thin muscle layer of circular fibres mixed with some longitudinal fibres (5–15 Μm thick). The ejaculatory duct receives abundant secretion from cells with coarse granular cyanophil secretion (~2 Μm) and less abundant cells with fine granular erythrophil secretion (<1 Μm). The epithelium of the penis papilla is columnar, progressively lower towards the tip, and the musculature is arranged in a circular subepithelial layer (5–15 Μm thick) followed by a longitudinal one (2.5 Μm thick). Five types of secretory cells run longitudinally in the papilla, with numerous openings through its lining epithelium: abundant cells with fine granular erythrophil secretion (<1 Μm) (type I); cells with coarse granular erythrophil secretion (~2 Μm) (type II); cells with cyanophil coarse granular secretion (>2 Μm) (type III); cells with fine densely arranged granular xanthophil secretion (~0.5 Μm) (type IV); and cells with cyanophil fine granular secretion (~0.5 Μm) (type V). Cell types I, II and V have their cell bodies located externally to the common muscle coat, the latter concentrating their openings mainly close to the ventral insertion of the papilla. Cell types III and IV have subepithelial cells bodies. The male atrium is lined by a columnar epithelium progressively higher towards the gonopore ( Figure 10 View FIGURES 9 – 13 ). Its muscularis is composed by a subepithelial circular layer (5–15 Μm thick) followed by a longitudinal subjacent layer (25–50 Μm thick). The same five types of glands described for the penis papilla discharge their secretions through the epithelium of the male atrium. Type V mainly opens into the proximal portion of the male atrium, close to the ventral insertion of the papilla.
Female reproductive system. The ovaries are situated at a distance of 16–21% body length from anterior end ( Table 3 View TABLE 3 ). They are ovoid and distally elongate. The ovaries are located between the sub-intestinal muscle layer and the nerve plate. The ovovitelline ducts emerge dorsally from anterior half of the ovaries and pass along the dorsal side of them, situated dorsally to the nerve plate ( Figure 11 View FIGURES 9 – 13 ). Approximately at the level of the gonopore, the ovovitelline ducts begin to ascend describing a sinuous course and then rise almost vertically, bending toward sagittal plane, along 150–200 Μm, and open into a common glandular ovovitelline duct. The common glandular ovovitelline duct (300–600 Μm in length) is an almost horizontal canal, dorsal to the female atrium ( Figures 7, 8 View FIGURES 7 – 8 , 13 View FIGURES 9 – 13 ). The vagina is a tubular diverticulum (200–300 Μm in length) that curves downward to open dorsally into the female atrium. The female atrium is a wide cavity with scarcely folded walls, continuous with the male atrium ( Figures 7–9 View FIGURES 7 – 8 View FIGURES 9 – 13 ).
The epithelium lining the ovovitelline ducts is ciliated and cuboidal. Cells with fine granular erythrophil secretion and scarce shell glands open into the vertical portions of the ovovitelline ducts, while the sagittal distal tracts receive abundant secretion from the shell glands ( Figure 12 View FIGURES 9 – 13 ). The common glandular ovovitelline duct is lined with a ciliated columnar epithelium ( Figure 13 View FIGURES 9 – 13 ), receiving abundant secretion of two types of shell glands: xanthophil coarse granular secretion (~2–3 Μm) and cyanophil coarse granular secretion (~3–4 Μm). Both glands have their cell bodies in the parenchyma, posteriorly to the copulatory apparatus ( Figure 9 View FIGURES 9 – 13 ). The vagina is lined with a ciliated columnar epithelium and the musculature is composed by circular subepithelial layer (5–15 Μm thick) and a longitudinal subjacent layer (10–15 Μm thick). The epithelium of the vagina receives few secretory cells of three types, similar to types I, III and IV of the penis papilla and male atrium. The female atrium is lined by a non-ciliated pluriestratified epithelium and the muscularis is arranged in two layers, circular subepithelial (10–25 Μm thick) and longitudinal subjacent (10–20 Μm thick). Four types of secretory cells, similar to types I, II, III and IV of the penis papilla and male atrium, open into female atrium. The common muscular coat is mainly composed by longitudinal fibres, thicker dorsally (~50 Μm thick) than ventrally (~25 Μm thick), besides some oblique and circular fibres.
Vitellaria are well developed and are dispersed throughout the body. In the anterior and pre-pharyngeal regions vitellaria extend dorsally, ventrally and between intestine branches ( Figures 4–6 View FIGURES 3 – 5 View FIGURE 6 ), being abundant behind the copulatory apparatus.

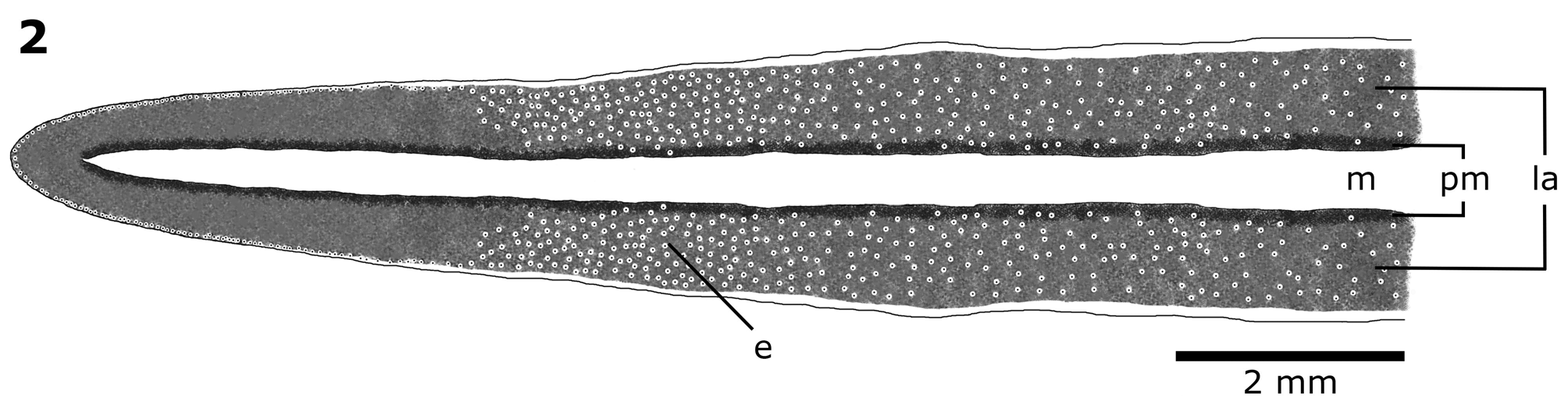
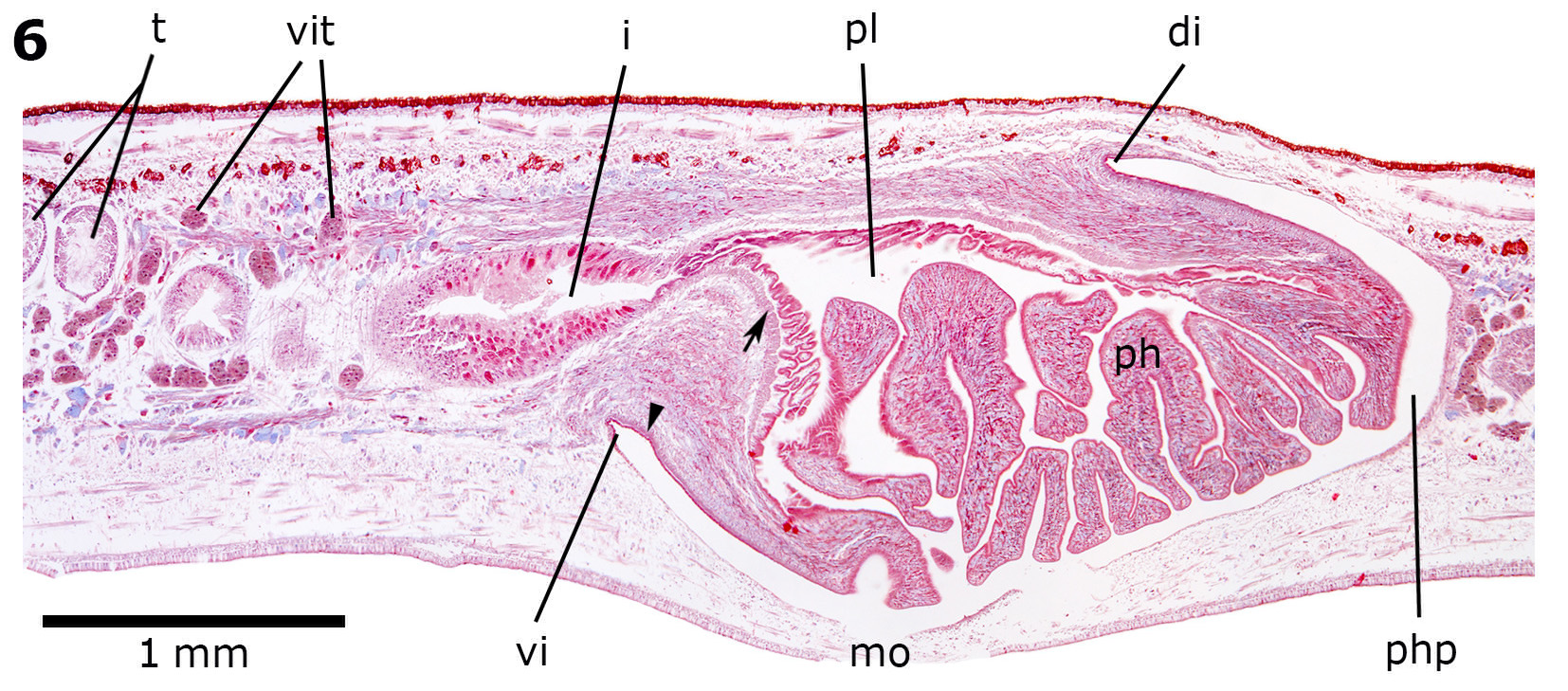
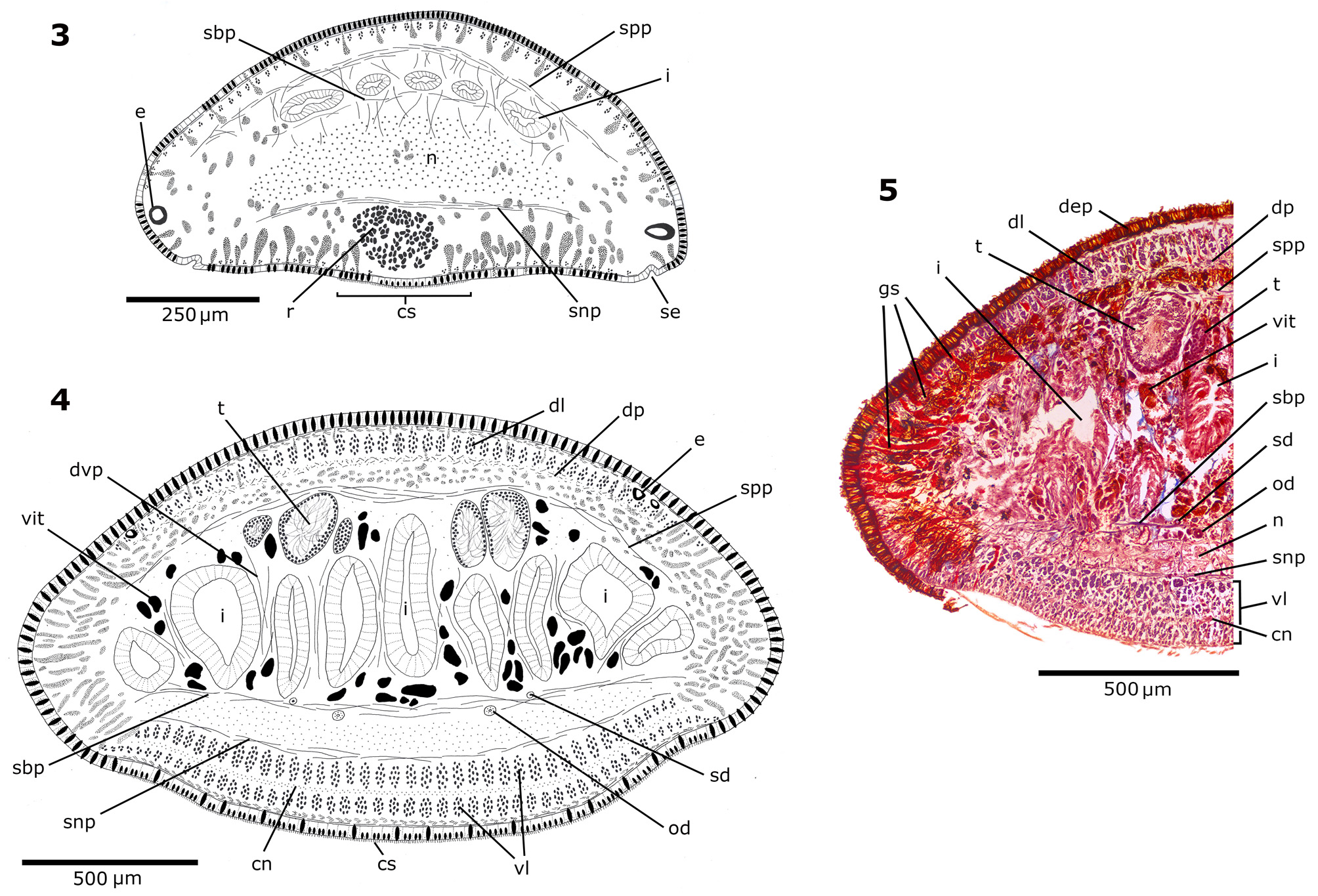
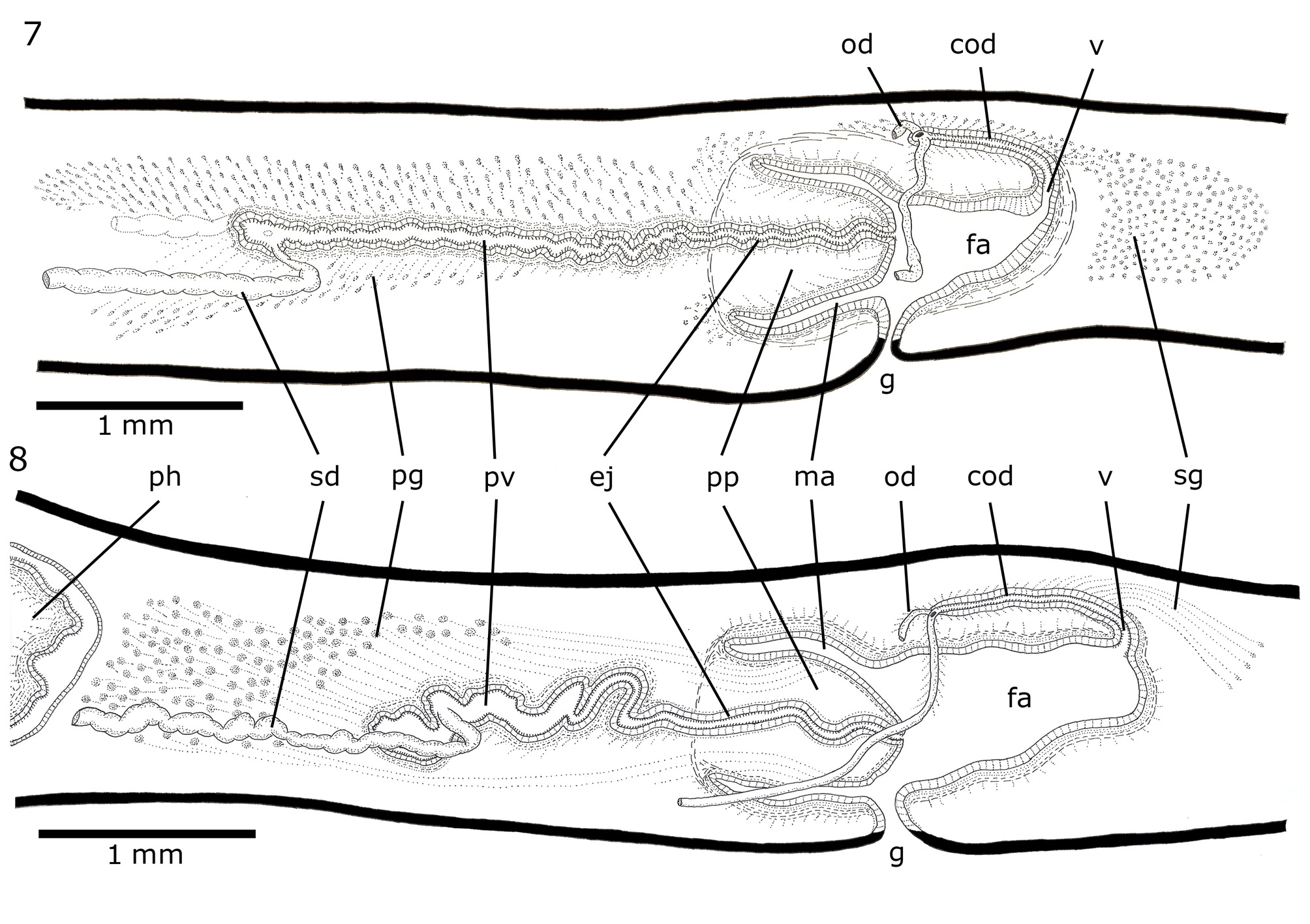
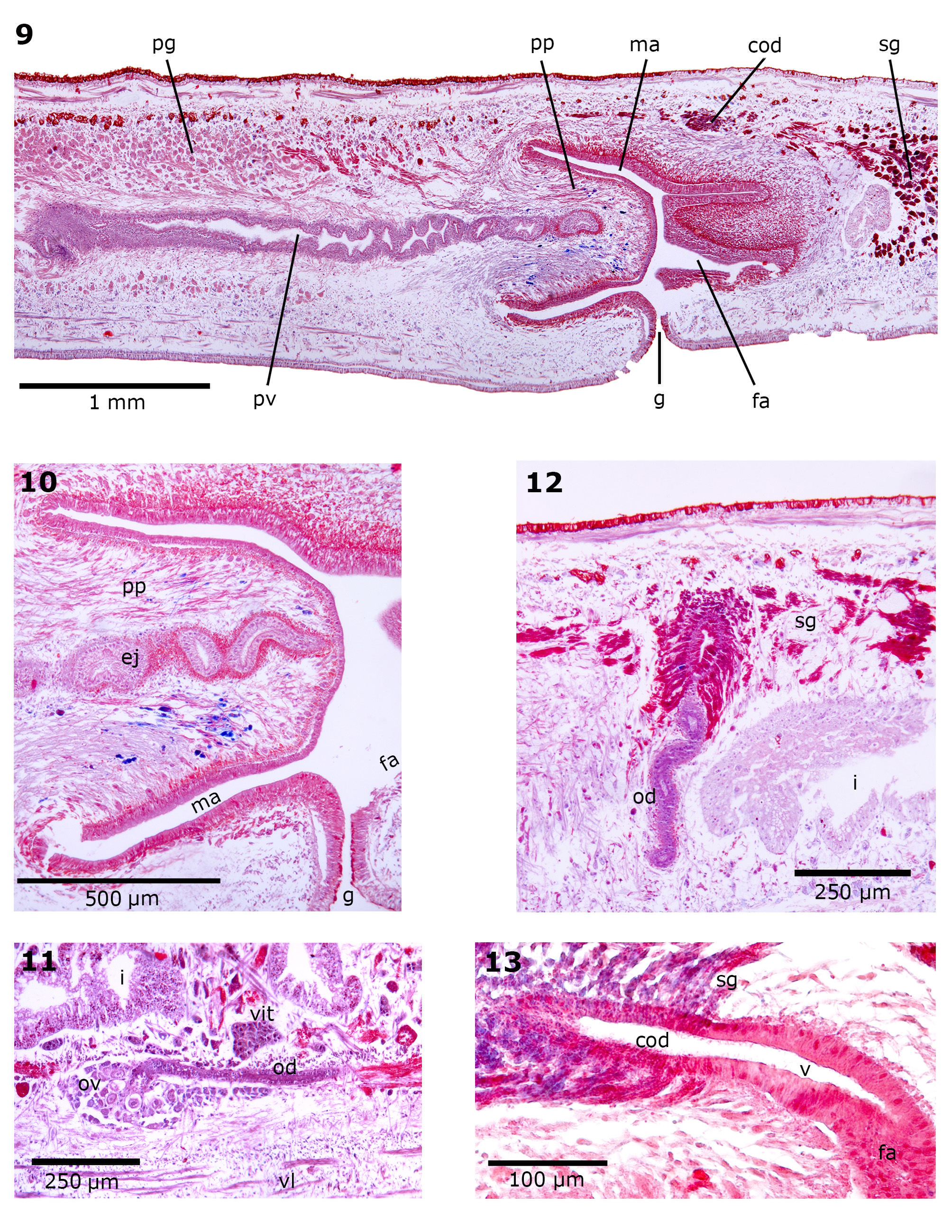
TABLE 1. Measurements (mm) of the type-specimens of Supramontana argentina sp. nov. All the measurements were obtained from fixed specimens. DG, distance of gonopore from anterior end; DM, distance of mouth from anterior end; DMG, distance between mouth and gonopore. The numbers given in parentheses represent the position relative to body length (%).
| Holotype MLP 6699 | Paratype MLP 6698 | Paratype MLP 6476 Paratype MLP 6475 | |
|---|---|---|---|
| Length | 39.5 | 17 | 37 45 |
| Maximum width | 2.9 | 1.6 | 2.1 2.4 |
| Height | 1.45 | 0.62 | 1.2 1.77 |
| DM | 23.7 (60) | 11 (65) | 20 (54) 27 (60) |
| DG | 32.8 (83) | 14 (82) | 28 (76) 36 (80) |
| DMG | 9.1 | 3 | 8 9 |
| Creeping sole (%) | 85 | 90 | 90 90 |
TABLE 2. Thickness of cutaneous (CM) and parenchymatic (PM) musculatures (µm), and CMI and PMI indices at prepharyngeal region of type-specimens of Supramontana argentina sp. nov.
| CM dorsal | Holotype MLP 6699 | Paratype MLP 6698 | Paratype Paratype MLP 6476 MLP 6475 |
|---|---|---|---|
| Circular Diagonal Longitudinal | 2.5 10 65 | 2.5 10 45 | 5 5 12.5 15 56 65 |
| Total CM ventral Circular | 77.5 2.5 | 57.5 2.5 | 73.5 85 5 5 |
| Diagonal Longitudinal (external) Longitudinal (internal) | 15 75 100 | 10 25 50–75 | 12.5 10 50 50 100 90 |
| Total CMI (%) PM dorsal | 192.5 19 50 | 87.5–112.5 18–21 20 | 167.5 155 20 14 35 37.5–50 |
| PM supra-intestinal PM sub-intestinal PM sub-neural | 15–35 20–40 5–10 | 10 12.5 10 | 35 50–62.5 25 25 15–25 50 |
| PMI (%) | 6–9 | 8 | 9–10 9–11 |
TABLE 3. Measurements (mm) of reproductive organs of type-specimens of Supramontana argentina sp. nov. LCGD, length of common glandular ovovitelline duct; LFA, length of female atrium; LMA, length of male atrium; LPP, length of penis papilla; LPV, length of prostatic vesicle; LV, length of vagina; T: BH, ratio of the height of testes to the height of the body. The numbers given in parentheses represent the position relative to body length (%).
| Holotype MLP 6699 | Paratype MLP 6698 | Paratype MLP 6476 | Paratype MLP 6475 | |
|---|---|---|---|---|
| Anteriormost testes | 9 (23) | 3.9 (23) | 6.5 (18) | – |
| Posteriormost testes | 22.7 (57) | 10.4 (61) | 17.9 (48) | 26 (58) |
| T:BH (%) | 15–22 | 15–18 | 9–23 | 12 |
| LPV | 2.25 | 0.9 | 1.8 | 2.2 |
| LPP | 0.8 | 0.5 | 0.38 | 0.48 |
| LMA | 0.85 | 0.6 | 0.7 | 0.6 |
| Location of ovaries | 8.1 (20) | 3.6 (21) | 6 (16) | – |
| LCGD | 0.4 | 0.3 | 0.6 | 0.3 |
| LV | 0.3 | 0.2 | 0.25 | 0.25 |
| LFA | 0.6 | 0.5 | 1.1 | 0.9 |
| MLP |
Museo de La Plata |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.