
Onychodactylus intermedius, Yoshikawa, Natsuhiko & Matsui, Masafumi, 2014
|
publication ID |
https://doi.org/10.11646/zootaxa.3866.1.3 |
|
publication LSID |
lsid:zoobank.org:pub:6334A96F-7716-4A4D-81AF-A81EAA06B9B7 |
|
DOI |
https://doi.org/10.5281/zenodo.6122899 |
|
persistent identifier |
https://treatment.plazi.org/id/51D86D4A-92DD-4010-809C-7BAD2178AC0D |
|
taxon LSID |
lsid:zoobank.org:act:51D86D4A-92DD-4010-809C-7BAD2178AC0D |
|
treatment provided by |
Plazi |
|
scientific name |
Onychodactylus intermedius |
| status |
sp. nov. |
Onychodactylus intermedius View in CoL sp. nov.
(Japanese name: Bandai-hakone-sanshou-uwo) (English name: Bandai clawed salamander) ( Figs. 4 View FIGURE 4 & 5 View FIGURE 5 )
Synonymy: Onychodactylus japonicus: Okada 1891 , p. 65 (part); Tago 1907, p. 239 (part); Stejneger 1907, p. 42 (part); Dunn 1923, p. 506 (part); Tago 1931, p. 200 (part); Okada 1937, p. 185 (part); Sato 1937, p. 44 (part); Sato 1943, p. 288 (part); Nakamura & Uéno 1963, p. 14 (part); Poyarkov et al. 2012, p. 36 (part).
Onychodactylus japonicus (Subclade II-A): Yoshikawa et al. 2008, p. 249.
Onychodactylus japonicus View in CoL (S-Tohoku group): Yoshikawa et al. 2010a, p. 33; Yoshikawa et al. 2012, p. 229.
Onychodactylus View in CoL sp. S-Tohoku group: Yoshikawa & Matsui 2013, p. 9; Yoshikawa et al. 2013, p. 441.
Etymology. The specific epithet “ intermedius ” is derived from the fact that the range of the new species occupies geographically intermediate position between southern O. japonicus and northern O. nipponoborealis . The Japanese and English names are after Mt. Bandai, a beautiful, symbolic volcano located northwest of the type locality. The new species is also distributed in Ura-bandai area, northern environs of the mountain.
Holotype. KUHE 47455, an adult male from the headstream of a small branch of Osawa River ( 140o11'E, 37o26'N, 790 m a.s.l.) in Koriyama-shi, Fukushima Prefecture, Japan, collected by N. Yoshikawa on 4 May 2013.
Paratypes. A total of 12 specimens: two males ( KUHE 47456–47457) and one female ( KUHE 47460) from the type locality, collected on 4 May 2013 by N. Yoshikawa; one male ( KUHE 48299) and three females ( KUHE 48300–48302) collected on 8 November 2013 by S. Ihara, N. Yoshikawa, and F. Endo from environs of Mt. Adatara, Otama-mura, Fukushima Prefecture; one male ( KUHE 37885 on 8 June 2006 by Y. Misawa) and one female ( KUHE 43309 on 15 June 2009 by Y. Misawa and N. Yoshikawa), all from environs of Nagai Dam, Nagaishi, Yamagata Prefecture; two males ( KUHE 44772, 44774) and one female ( KUHE 44773) from Budo, Murakami-shi, Niigata Prefecture collected on 12 May 2011 by Y. Misawa.
Referred specimens. See APPENDIX 1.
Diagnosis. The new species is a member of the genus Onychodactylus diagnosed by the following characters: lungs absent; black horny claws present on tips of fingers and toes of breeding adults and larvae; vomerine teeth in two short, transverse, arch-shaped series; larvae with skin folds on posterior edges of limbs; breeding males with dermal flaps on posterior edge of hindlimb; black tubercles and asperities on palm and sole in breeding males and on sole in breeding females; breeding in flowing water under the ground; eggs large, pigmentless, small in number.
It is further assigned to a member of the O. japonicus species complex by the genetic evidence and presence of a distinct dorsal stripe. Dorsal stripe or marking yellow, ochre, brown, or reddish brown on grayish brown or purplish brown background; body size small to moderate with mean SVL (±1SD) of 63.0± 4.9 mm and 65.5± 4.1 mm in males and females, respectively; tail longer than SVL in males, but equal to or only slightly longer in females; snout relatively short; usually lacking wedge-shaped dark marking on chest; ventrum purplish brown, purplish gray or light gray, with white dots; presacral vertebrae including atlas usually 18; costal grooves 12 or 13; right and left vomerine tooth series usually continuous without gap, series curving distinctly; vomerine tooth series tending to have anteriorly directed short sub-branch at inner meeting; relatively large number of vomerine teeth, about 16 on one side of series.
Description of holotype. An adult male with SVL 61.0 mm ( Figs. 4 View FIGURE 4 A–B & 5); body moderately slender; skin smooth; head oval and depressed, longer than wide; neck slightly narrower than head; snout rounded, projecting beyond lower jaw; nostril close to eye; eye large, slightly shorter than snout, prominently protruded; gular fold posteroventrally to head; parotoid gland developed, oval, ca. 1.9 times longer than wide, posterior end at the level of gular fold; postorbital groove obvious; vomerine teeth in two transverse, distinctly curved arch-shaped series without gap in between ( Fig. 6 View FIGURE 6 A); 14 and 12 teeth on right and left series, respectively; inner end of each vomerine tooth series curving to form short sub-branch directed anteriorly; series anteriorly scattered with dark pigments; forelimb thin; relative length of fingers I<IV<II<III; hindlimb slightly longer and more robust than forelimb; relative length of toes I<V<II<III<IV; adpressed limbs separated by 0.5 costal folds; no characteristics showing breeding condition; dermal flap on posterior edge of hindlimb not developed; tips of fingers and toes rounded without black horny claws; no black tubercles and asperities on palm and sole; trunk elongated and cylindrical; distinct middorsal groove from level of parotoid gland to cloaca; well-developed 12 costal grooves on both sides of trunk; presacral vertebrae including atlas numbering 18; base of tail including around cloaca not swollen; cloaca longitudinal slit, length 9.8% SVL, with anterior two-fifths of its edge slightly swollen, forming inverse narrow Vshaped precloacal skin fold; precloacal skin fold as wide as one third of cloaca length; no precloacal black spine series; tail long, 126.6% length of SVL, cylindrical at base, increasingly compressed toward tip; tail slightly lower toward tip, laterally compressed in posterior half and extremely at tip; tip of tail slightly pointed in lateral view; tail highest (8.7% SVL) at proximal one-fifth.
Measurements (in mm) of the holotype. SVL 61.0, HL 14.4, HW 8.3, TAL 77.2, AGD 30.8, FLL (L) 15.7, FLL (R) 16.1, HLL (L) 19.5, HLL (R) 18.4, UEL (L) 4.0, UEL (R) 4.1, IOD 2.7, END (L) 1.8, END (R) 1.9, ICD 5.0, IND 3.9, SL (L) 4.3, SL (R) 4.4, CW 7.6, BTAH 5.1, BTAW 5.7, MTAH 4.6, MTAW 3.8.
Color: In life, gray-brown with distinct, wide ochre dorsal stripe from snout to tip of tail ( Figs. 4 View FIGURE 4 & 5 View FIGURE 5 ); dorsal stripe wavy but sharply defined from background; on tail, dorsal stripe narrowed toward tip, and brightest at base; side of body gray-brown with silvery white dots, gradually fading toward ventrum; ventrum grayish with silvery white dots; upper iris golden slightly mottled with brown, lower iris dark-brown. In alcohol, color and pattern fading generally, and dorsal ochre coloration bleached.
Variation. The following description of variation is based on the maximum number of 17 adult males and 10 adult females obtained from Yamagata, Niigata, and Fukushima Prefectures. Morphometric data are summarized in Table 1 together with those of seven congeneric species.
There was no significant difference between males (63.0± 4.9 mm, n = 17) and females (65.5± 4.1 mm, n = 10) in SVL (t = 1.321, p = 0.198: Table 1). In values relative to SVL, males had longer tail and shorter trunk than females (Table 1). Males also had more robust hindlimb and posteriorly more projecting distal end of fibula than females.
A small number of adult specimens we could collect in the breeding season showed following characteristics: black claws on fingers and toes, swollen parotoid gland, black precloacal spine series, and laterally compressed tail in both sexes; dermal skin fold on posterior edge of hindlimb, black tubercles and asperities on palm and sole, swollen cloaca, and inverse V-shaped precloacal skin fold in males; inverse narrow U-shaped precloacal skin fold in females.
Number of costal grooves varied from 12 to 13 (Table 2) and was not sexually different (t = 1.782 and 1.576, p = 0.087 and 0.127 for left and right sides, respectively). Number of presacral vertebrae including atlas did not differ between sexes (t = 0.031, p = 0.975), and varied from 18 to 19 (mode = 18). There was no sexual difference in number of vomerine teeth (11 to 22, mean = 15.9± 2.4 in males and 8 to 25, 15.5± 4.3 in females: t = 0.250, p = 0.807). Anteriorly directed, short sub-branch of vomerine tooth series at inner meeting occurred in 63.0% of specimens, and did not differ sexually ( p = 0.810: Table 3).
Color pattern in life was variable among individuals. Dorsal stripe varied from straight, uneven, to broken into continuous or discontinuous series of flecks. Color of dorsal stripe varied from yellow, ochre, brown to reddishbrown, while background color varied from gray-brown, dark-brown, to purplish-brown. Border of dorsal stripe and background color was generally sharply defined, but was often obscured. Color on lateral to ventral sides was gray, purplish-gray, or purplish-brown with varying amount of silvery white dots.
Larva: Larvae found in the open streams had SVL of 16.3–38.5 mm and TAL of 12.5–40.2 mm, resulting in total length (TL) of 28.8–78.6 mm ( Fig. 7 View FIGURE 7 ). Head rectangular and blunt at snout; three pairs of short external gills; labial fold well developed at posterior half of upper jaw; caudal fin low but well-developed dorsally and ventrally; dorsal fin relatively higher than ventral fin; origin of dorsal fin at level of hindlimb to cloaca; ventral fin originating from around posterior four-fifths to two-thirds of tail; tail tip moderately rounded; skin fold on posterior edge of limb; dark asperities on surfaces of palm and sole; digits with acute and curved black claws.
Two color morphs were recognized in larvae based on dorsal color. One was the unstriped type, possessing varying amounts of dark-brown mottling on light-brown or yellowish-gray background. Dark-brown mottling varied from very weak (nearly uniformly yellowish-gray: Fig. 7 View FIGURE 7 F) to strong (nearly uniformly dark-brown: Fig. 7 View FIGURE 7 A–B), although most specimens were moderately mottled ( Fig. 7 View FIGURE 7 C–D). Another is the striped type, possessing wide and distinct reddish-brown dorsal stripe extending from head to tail tip on dark-brown background ( Fig. 7 View FIGURE 7 E). The striped type larvae were less frequently found than did the unstriped type in most populations. The color of the ventrum is slightly transparent, whitish, or grayish and sometimes slightly pigmented.
Comparisons. Morphological data for comparison with the other congeneric species are summarized in Tables 1–4. Some other data are cited from Yoshikawa et al. (2013) and Poyarkov et al. (2012).
Onychodactylus intermedius differs from the continental Onychodactylus species ( O. fischeri , O. koreanus , O. zhangyapingi , and O. zhaoermii ) in having a distinct wide dorsal stripe (vs. spotted, mottled, or indistinctly striped dorsum in continental species).
Onychodactylus intermedius has significantly smaller SVL in both sexes than O. fischeri . In characters relative to SVL, O. intermedius significantly differs from O. fischeri in having a shorter tail in males, shorter trunk, longer forelimb and hindlimb, longer and wider head, larger IND, IOD, CW, and ICD in both sexes (Table 1). Onychodactylus intermedius is also significantly different from O. fischeri in having smaller number of presacral vertebrae and costal grooves (Table 2).
From O. koreanus , O. intermedius differs in having shorter tail, longer trunk, and wider chest, all relative to SVL (Table 1), and smaller number of presacral vertebrae in both sexes; smaller SVL, shorter hindlimb relative to SVL, and fewer costal grooves in males (Table 2); and shorter head relative to SVL in females.
Morphological comparisons of O. intermedius with the other species in the O. japonicus species complex produced the following results (Tables 1, 2, 3 & 4). Onychodactylus intermedius is morphologically very similar to O. japonicus , but differing from it by: smaller SVL in males; relatively shorter tail in both sexes (121.8% of SVL in males and 101.9% in females vs. 125.0% and 106.7% in O. japonicus ); larger number of vomerine teeth in males; vomerine tooth series tending to have anteriorly curved short sub-branch at inner meeting. In coloration, O. intermedius is variable and looks similar to O. japonicus from eastern Japan, but differs from O. japonicus from western Japan and from the type locality, in which the stripe is distinct, sharply defined red or orange. A pair of dark marking on chest appears less frequently in O. intermedius (3.7% of specimens examined) than in O. japonicus (68.6%).
TAB LE 2. Variation in the number of presacral vertebrae (PSVN) and costal grooves (CGN) on each side of the body in eight O n ych od ac t yl u s species. Bold numbers indicate modal values.
PSVN CGN(L) CGN(R)
Species Sex n 17 18 19 20 21 22 11 12 13 14 15 16 17 11 12 13 14 15 16 17 O . intermedius sp. nov.
male 17 11 6 10 7 11 6
female 10 8 2 4 6 5 5 O. fuscus sp. nov.
male 6 3 3 1 5 1 5
female 5 4 1 4 1 4 1 O. japonicus
(eastern pops.) male 60 8 41 11 7 41 12 8 42 10
female 66 3 57 6 1 52 13 1 48 17 (combined) male 78 8 53 17 7 50 20 1 8 50 19 1
female 75 3 58 14 1 52 22 1 48 26 O. kinneburi
male 36 32 4 2 32 2 1 34 1
female 33 1 30 2 28 5 2 27 4 O. nipponoborealis
male 20 1 18 1 5 14 1 6 13 1
female 26 1 12 13 3 19 4 3 19 4 O. tsukubaensis
male 6 5 1 1 5 1 5
female 10 1 9 1 9 2 8
male 8 5 3 5 3 5 3
female 2 2 1 1 1 1 O . fischeri
male 9 1 7 1 4 3 1 5 2 1
female 3 2 1 2 1 2 1 From O. kinneburi , O. intermedius significantly differs in having smaller SVL in both sexes, and relatively wider chest in females (13.6% of SVL vs. 12.2% in O.kinneburi ). Onychodactylus intermedius also differs from O. kinneburi in having significantly smaller numbers of presacral vertebrae in both sexes (mode = 18 vs. 19 in O. kinneburi ) and costal grooves (mode = 12 in males and 13 in females vs. 13 in both sexes in O. kinneburi ). In O. intermedius , vomerine tooth series is distinctly curved (vs. shallowly curved in O. kinneburi ), forming anteriorly curved sub-branch at inner meeting (vs. tending to lack short sub-branch in O. kinneburi ). Number of vomerine teeth in male O. intermedius is significantly larger than O. kinneburi . In coloration, O. intermedius with ochre or brownish dorsal stripe with dark-brown background, and often with silvery white dots on ventrolateral side, can be easily distinguished from O. kinneburi with sharply defined yellowish-orange or yellow dorsal stripe or continuous series of blotches on uniformly black background, and usually lacking silvery white dots on body.
From O. nipponoborealis , O. intermedius differs in having a narrower head in males (14.5% of SVL vs. 15.1% in O. nipponoborealis ), as well as a shorter snout (7.1% of SVL in males and 7.3% in females vs. 7.9% and 7.8%, in O. nipponoborealis ), and wider intercanthal space (59.6% of HW in males and 59.3% in females vs. 56.9% and 56.8% in O. nipponoborealis ) in both sexes. In coloration, O. intermedius is similar to O. nipponoborealis , and is difficult to distinguish it based only on coloration. Dark marking on chest, usually absent in O. intermedius (present only in 3.7% of specimens examined), is more frequently present in O. nipponoborealis (65.2%: Table 4).
TABLE 3. Variation in the number of vomerine tooth series (means±1SD, followed by ranges in parenthesis), gap between right and left sides of the vomerine tooth series, and short sub-branch of vomerine tooth series at the inner meeting (number followed by percentage in parenthesis) in eight Onychodactylus species.
Gap Short sub-branch of VTS
Species Sex n VTN(L) VTN(R) VTN (average) present absent L+R L or R absent
O. intermedius sp. nov.
male 17 16.1±2.9 15.6±1.9 15.9±2.4 4 13 11 - 6 (11–22) (12–18) (11–22) (23.5) (76.5) (64.7) - (35.3) female 10 16.1±4.6 14.9±4.1 15.5±4.3 4 6 4 2 4 (10–25) (8–21) (8–25) (40.0) (60.0) (40.0) (20.0) (40.0) Sum 27 16.1±3.5 15.4±2.8 15.7±3.2 8 19 15 2 10 (10–25) (8–21) (8–25) (29.6) (70.4) (55.6) (7.4) (37.0)
O. fuscus sp. nov.
male 6 12.8±1.5 12.0±1.9 12.4±1.7 6 - - - 6 (11–15) (9–15) (9–15) (100) - - - (100) female 5 14.8±3.1 15.2±3.3 15.0±3.0 4 1 3 - 2 (11–18) (12–19) (11–19) (80.0) (20.0) (60.0) - (40.0) Sum 11 13.7±2.5 13.5±3.0 13.6±2.7 10 1 3 - 8 (11–18) (9–19) (9–19) (90.9) (9.1) (27.3) - (72.7)
(eastern pops.) male 57 11.6±3.6 11.8±3.9 11.7±3.7 17 40 7 4 46 (5–21) (7–26) (5–26) (29.8) (70.2) (12.3) (7.0) (80.7) female 61 13.9±4.1 14.4±4.2 14.1±4.1 15 46 10 4 47 (5–22) (4–25) (4–25) (24.6) (75.4) (16.4) (6.6) (77.0) Sum 118 13.2±4.3 13.1±4.2 13.0±4.1 32 86 17 8 93 (5–23) (4–26) (4–26) (27.1) (72.9) (14.4) (6.8) (78.8)
(combined) male 75 12.7±4.4 12.7±4.0 12.7±4.2 30 45 13 6 56 (5–23) (7–26) (5–26) (40.0) (60.0) (17.3) (8.0) (74.7) female 70 14.2±4.1 14.7±4.2 14.5±4.1 22 48 13 6 51 (5–22) (4–25) (4–25) (31.4) (68.6) (18.6) (8.6) (72.8) Sum 145 13.4±4.3 13.7±4.2 13.6±4.2 52 93 26 12 107 (5–23) (4–26) (4–26) (35.9) (64.1) (17.9) (8.3) (73.8)
male 36 13.6±3.3 13.5±3.2 13.6±3.2 12 24 2 7 27 (9–22) (9–20) (9–22) (33.3) (66.7) (5.6) (19.4) (75.0) female 33 15.6±3.7 16.4±3.9 16.0±3.8 13 20 8 10 15 (9–23) (8–25) (8–25) (39.4) (60.6) (24.2) (30.3) (45.5) Sum 69 14.6±3.6 14.9±3.8 14.7±3.7 25 44 10 17 42 (9–23) (8–25) (8–25) (36.2) (63.8) (14.5) (24.6) (60.9)
male 20 15.0±5.0 15.2±4.4 15.1±5.8 6 14 9 2 9 (4–24) (5–23) (4–24) (30.0) (70.0) (45.0) (10.0) (45.0) female 26 15.4±3.8 15.0±3.6 15.2±3.6 9 17 14 5 7 (8–23) (9–20) (8–23) (34.6) (65.4) (53.8) (19.2) (27.0) Sum 46 15.2±4.3 15.1±3.9 15.2±4.2 15 31 23 7 16 (4–24) (5–23) (4–24) (32.6) (67.4) (50.0) (15.2) (34.8)
male 6 16.0±3.2 15.5±3.0 15.8±3.0 1 5 - 1 5 (12–20) (12–19) (12–20) (16.7) (83.3) - (16.7) (83.3) female 10 18.2±2.2 17.1±2.5 17.8±2.3 - 10 7 - 3 (14–22) (12–20) (12–22) - (100) (70.0) - (30.0) Sum 16 17.4±2.7 16.7±2.8 17.0±2.7 1 15 7 1 8 (12–22) (12–20) (12–22) (6.3) (93.7) (43.7) (6.3) (50.0)
male 8 18.1±3.7 18.8±2.1 18.4±2.9 1 7 4 2 2 (15–26) (15–22) (15–26) (12.5) (87.5) (50.0) (25.0) (25.0) female 2 17.0±2.8 16.5±3.5 16.8±2.6 - 2 1 - 1 (15–19) (14–19) (14–19) - (100) (50.0) - (50.0) Sum 10 17.9±3.4 18.3±2.4 18.1±2.9 1 9 5 2 3 (15–26) (14–22) (14–26) (10.0) (90.0) (50.0) (20.0) (30.0)
male 9 14.9±3.2 14.0±3.2 14.4±3.1 4 5 4 - 5 (10–20) (9–20) (9–20) (44.4) (55.6) (44.4) - (55.6) female 3 13.3±1.5 13.0±2.6 13.2±1.9 1 2 1 - 2 (12–15) (11–16) (11–16) (33.3) (66.7) (33.3) - (66.7) Sum 12 14.5±2.9 13.8±3.0 14.1±2.9 5 7 5 - 7 (10–20) (9–20) (9–20) (41.7) (58.3) (41.7) - (58.3) In comparison with O. tsukubaensis , O. intermedius is significantly smaller in male SVL. Onychodactylus intermedius also differs significantly from it in having longer tail (121.8% of SVL in males and 101.9% in females vs. 102.3% and 90.4% in O. tsukubaensis ), narrower head (14.5% of SVL in males and 14.8% in females vs. 15.4% and 15.6% in O. tsukubaensis ), shorter snout (7.1% of SVL in males and 7.3% in females vs. 7.7% and 7.9% in O. tsukubaensis ), and wider interorbital space (31.8% of HW in males and 32.2% in females vs. 28.3% and 27.8% in O. tsukubaensis ). Number of costal grooves were significantly larger in O. intermedius than O. tsukubaensis in both sexes (mode = 12 in males and 13 in females vs. 12 in both sexes). Onychodactylus intermedius is similar to O. tsukubaensis in coloration and is difficult to be distinguished from it based only on coloration.
In the larval stage, O. intermedius looks very similar to O. japonicus and O. nipponoborealis and is difficult to distinguish from them. Onychodactylus intermedius larva differs from O. tsukubaensis in having longer tail ( Fig. 8 View FIGURE 8 ), and in lacking silvery white mottlings on body and light-yellow dorsocaudal stripe ( Fig. 7 View FIGURE 7 ). Larval O. intermedius differs from O. kinneburi in lacking orangish dorsal markings.
Karyotype. No karyotypic information is available for O. intermedius .
Genetic characteristics. Onychodactylus intermedius sp. nov. corresponds to Subclade II-A or S-Tohoku group in the previous studies ( Yoshikawa et al. 2008, 2010a, 2012), and detailed phylogenetic position and genetic relationships have been discussed. However, this study revealed that Subclade II-C, which is newly reported here and described below, is the sister taxon of O. intermedius ( Fig. 2 View FIGURE 2 ). The mean uncorrected p-distance for cytb gene between O. intermedius and closely related species O. tsukubaensis (Subclade II-B) was 5.33% (4.90–5.70%), and Subclade II-C was 4.15% (3.59–4.73%). Within the species, the distance varied up to 1.67%, and no distinct geographic genetic structure was found. Onychodactylus intermedius is parapatric with O. japonicus and O. nipponoborealis , but they are nearly reproductively isolated, with only a few hybrids found around species boundaries ( Yoshikawa et al. 2012).
Fecundity and natural history. Breeding season of O. intermedius sp. nov. is not yet confirmed. However, our field observations suggest that the new species breeds from late spring to early summer (summer breeding) and/or late autumn to early winter (winter breeding), because adults with breeding characters were collected in both seasons, although the season seems to depend on the locality: a clawed male, KUHE44772, was collected in early May from Murakami-shi, Niigata Pref.; a clawed female, KUHE38310, with mature eggs was collected in late October from Nagai-shi, Yamagata Pref.; and clawed females, KUHE 48301–48302, with mature eggs were collected in early November from Otama-mura, Fukushima Pref. (these were induced to oviposit in the laboratory, see below). It is also possible that some populations breed biannually as known in O. japonicus in Ishikawa Prefecture ( Akita, 1989).
The breeding site is presumed to be the underground flowing water near headstreams, as is the case in other Onychodactylus species. We obtained two pairs of egg sacs from females from Otama-mura, Fukushima Prefecture in the laboratory on 24 December 2013 by gonadotropin induction ( Fig. 9 View FIGURE 9 A). Egg sacs were attached on the stone and strongly adhered by short gelatinous stalks. The outer gelatinous layer was transparent, elastic, and strong. Egg sacs were cylindrical and spindle-shaped, in which eggs were arranged in a single row and two rows, respectively. Very weak, longitudinal grooves could be seen on the surface of the egg sacs. The clutch size was nine (six and three eggs in each sac) and 22 (10 and 12 eggs), with the mean ±1SD of 15.8±5.3 (n = 4), and diameters of 31 eggs ranged from 4.5–5.7 mm (5.2±0.4). The eggs were pigmentless and entirely yellowish-white.
Larval life history of O. intermedius is poorly known, but the larvae found in open streams ranged 28.8–78.6 mm in TL. Larvae inhabit the streams, and may grow up to metamorphose after two or more years, as known in other congeners (Yoshikawa & Matsui 2013; Yoshikawa et al. 2013). The season of metamorphosis is unknown, but the size at metamorphosis is 33.2–45.1 mm in SVL and 61.7–86.7 mm in TL or larger based on the metamorphosed juveniles found in the wild. Metamorphosed juveniles were often found in the forest floor around the stream.
Ecology of adults in the non-breeding season is not well known, but they inhabit cool and humid forest-floors near streams in well-forested mountains ( Fig. 9 View FIGURE 9 B), like the other Onychodactylus species. A case of predation by a snake Amphiesma vibakari vibakari was observed in the type locality on 11 June 2013 (N. Yoshikawa & T. Nakano, unpublished).
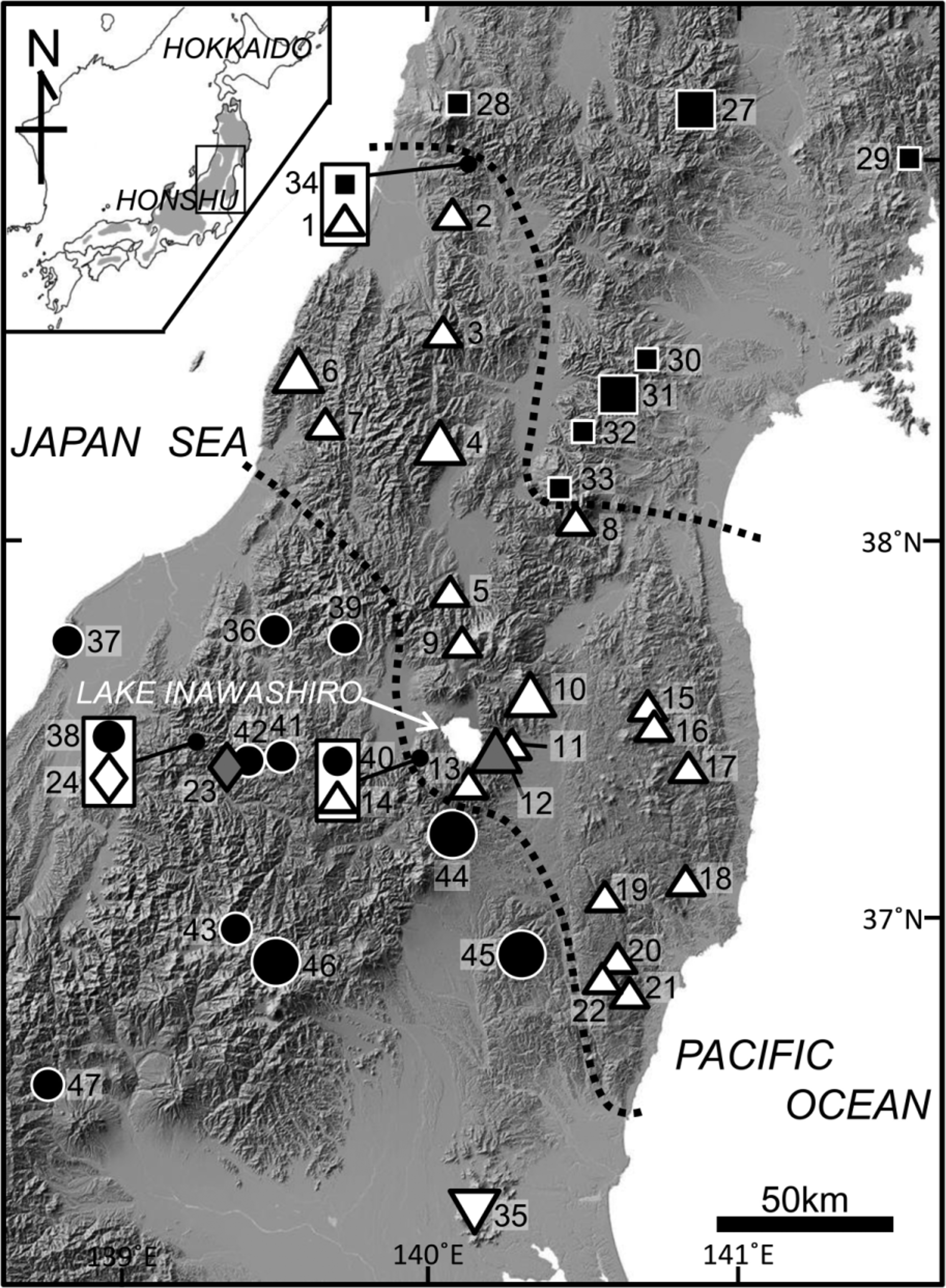
Range. Onychodactylus intermedius sp. nov. is known from northern to southeastern Yamagata, southern Miyagi, and northeastern half of Fukushima Prefectures of southern Tohoku District, northern Niigata Prefecture of Hokuriku District, and northeastern Ibaraki Prefecture of Kanto District, all in eastern Honshu. As mentioned above, the new species borders two congeners, O. nipponoborealis and O. japonicus . Distributional pattern of the new species is complex and the species boundaries ( Fig. 1 View FIGURE 1 ) mostly do not correspond to potential geographic barriers such as rivers or lowlands.
Conservation. At present, the new species is abundant and the most common salamander species in the known distributional range. The new species is listed as O. japonicus on the Red List of Ibaraki Prefecture as Vulnerable (Ibaraki Prefecture 2000), and the Red lists of Miyagi, Niigata, and Yamagata Prefectures as Near Threatened (Miyagi Prefecture 2013; Niigata Prefecture 2001; Yamagata Prefecture 2003), but not listed in the Red List of Fukushima Prefecture (Fukushima Prefecture 2002). Because this species strongly depends on stability of streams and forests, careful managements and monitoring on its habitats and populations are needed to maintain its population size, especially in peripheral areas, such as the southeastern and eastern parts of its range. We propose that the new species to be designated as Near Threatened in the Red lists of IUCN and Ministry of the Environment of Japan.







| KUHE |
Kyoto University, Graduate School of Human and Environmental Studies |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |