
Guimaraesiella (Dicrurobates) lurida Gustafsson, 2020
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4885.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:081203D8-39FF-41C3-A79A-BB63F47AB3B1 |
|
DOI |
https://doi.org/10.5281/zenodo.4332150 |
|
persistent identifier |
https://treatment.plazi.org/id/F85A70C6-AD91-4314-9CD1-4815A6E78A72 |
|
taxon LSID |
lsid:zoobank.org:act:F85A70C6-AD91-4314-9CD1-4815A6E78A72 |
|
treatment provided by |
Plazi |
|
scientific name |
Guimaraesiella (Dicrurobates) lurida Gustafsson |
| status |
new species |
Guimaraesiella (Dicrurobates) lurida Gustafsson & Bush, new species
( Figs 29–35 View FIGURES 29–30 View FIGURES 31–35 )
urn:lsid:zoobank.org:act:F85A70C6-AD91-4314-9CD1-4815A6E78A72
Type host. Dicrurus leucophaeus Vieillot, 1817 —ashy drongo.
Type locality. Chiang Saen , Chieng Rai Province, Thailand .
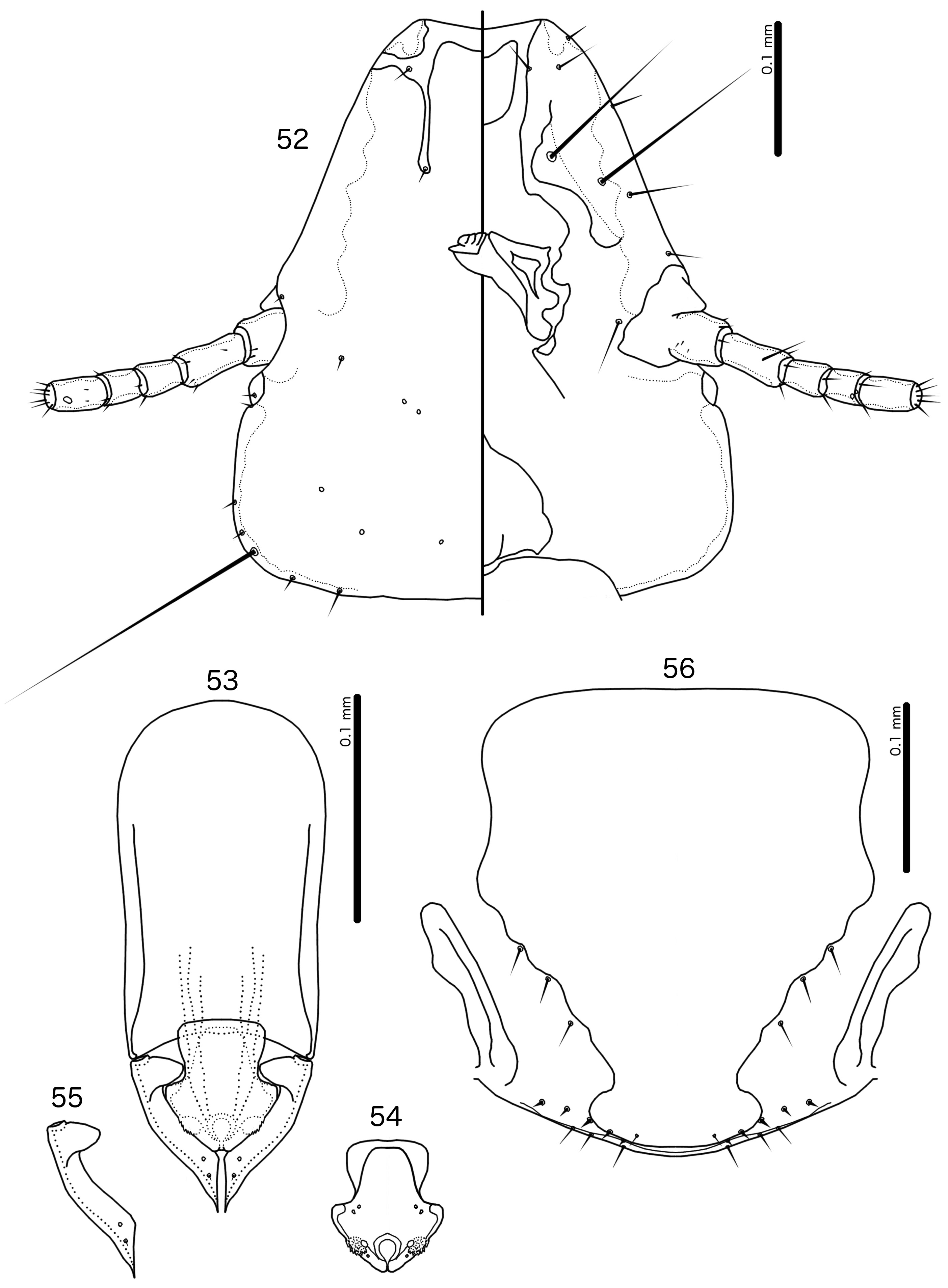
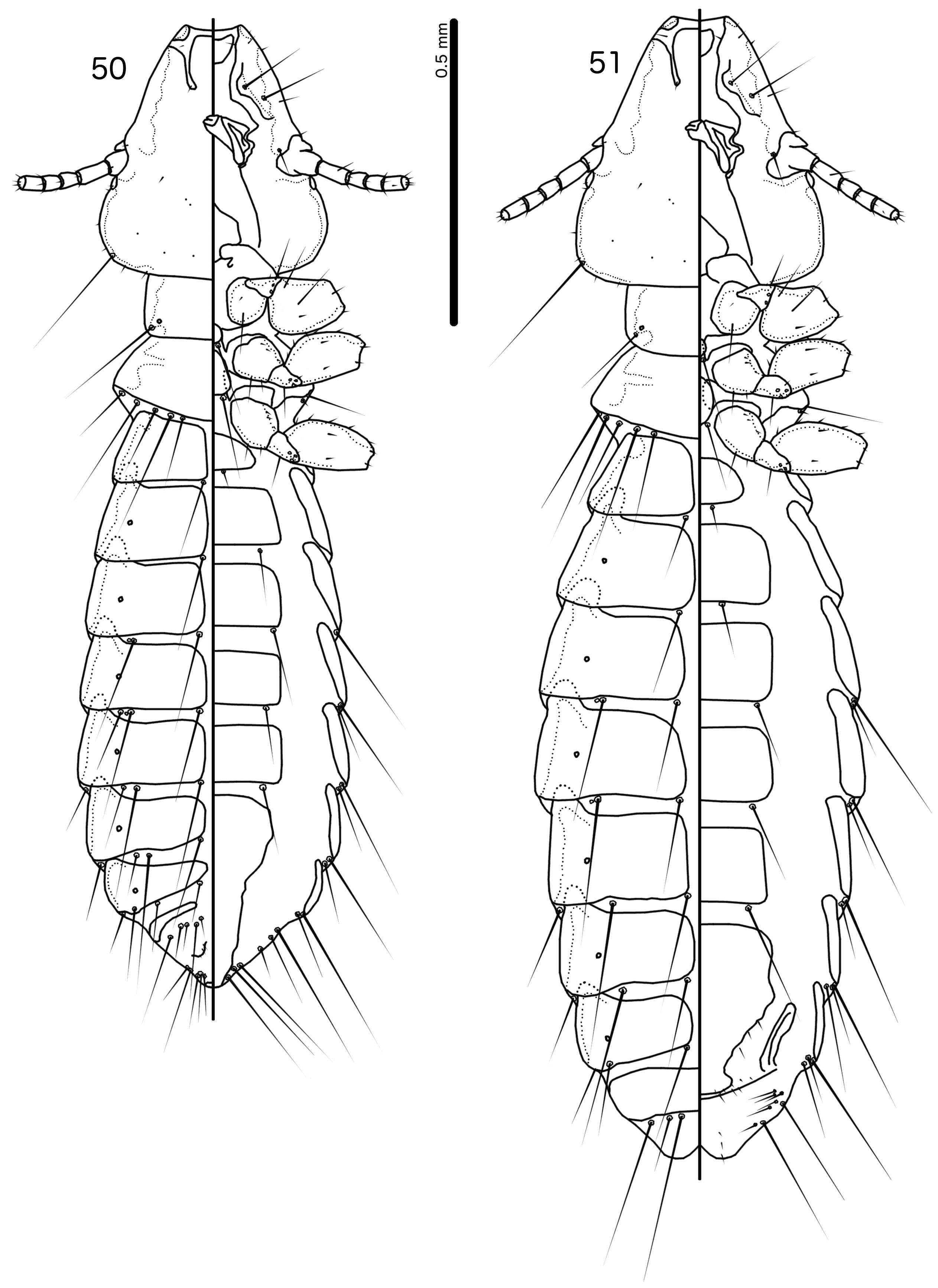
Diagnosis. Guimaraesiella (Dicrurobates) lurida is morphologically closest to Guimaraesiella (Di.) regis n. sp. (see below), but these two species can be separated by the following characters: (1) the dorsal preantennal suture reaches lateral margin of head in Guimaraesiella (Di.) regis ( Fig. 52 View FIGURES 52–56 ), but not in Guimaraesiella (Di.) lurida ( Fig. 31 View FIGURES 31–35 ); (2) the ventral anterior plate is wider than long in Guimaraesiella (Di.) lurida ( Fig. 31 View FIGURES 31–35 ), but longer than wide in Guimaraesiella (Di.) regis ( Fig. 52 View FIGURES 52–56 ); (3) aps is present on female tergopleurite VIII in Guimaraesiella (Di.) lurida ( Fig. 30 View FIGURES 29–30 ), but absent in Guimaraesiella (Di.) regis ( Fig. 51 View FIGURES 50–51 ); (4) the anterior margin of the proximal mesosome is concave in Guimaraesiella (Di.) lurida ( Fig. 33 View FIGURES 31–35 ), but is straight in Guimaraesiella (Di.) regis ( Fig. 54 View FIGURES 52–56 ); (5) the ventral sclerite tapers markedly anteriorly, with the anterior end about a third as wide as the proximal mesosome at its widest point in Guimaraesiella (Di.) lurida ( Fig. 33 View FIGURES 31–35 ), but it tapers less markedly, with the narrowest point about half as wide as the proximal mesosome in Guimaraesiella (Di.) regis ( Fig. 54 View FIGURES 52–56 ).
For differences between Guimaraesiella (Di.) lurida and the other morphologically close species— Guimaraesiella (Di.) nana n. sp. and Guimaraesiella (Di.) luzonica n. sp. — see under the Diagnosis of each of these species.
Description. Both sexes. Head shape and chaetotaxy as in Fig. 31 View FIGURES 31–35 . Lateral margins of preantennal area straight to slightly convex, frons broadly flattened; marginal carina broad, irregular, narrowing anteriorly; dorsal preantennal suture reaches dsms and extends toward but does not reach ads, and does not reach lateral margins of head; ventral anterior plate broad, but short, roughly, semi-circular; coni broad but short, temples rounded; gular plate rhombic with anterior and lateral points ( Fig. 31 View FIGURES 31–35 ). Thoracic and abdominal segments as in Figs 29–30 View FIGURES 29–30 .
Male. Thoracic and abdominal chaetotaxy as in Fig. 29 View FIGURES 29–30 ; aps absent on tergopleurite IV, but present on tergopleurites V–VII. Genitalia as in Figs 32–34 View FIGURES 31–35 : basal apodeme slightly trapezoidal, narrowing anteriorly with rounded anterior end and slight or no constriction at mid-length ( Fig. 32 View FIGURES 31–35 ). Proximal mesosome broad, widening anteriorly, with concave proximal margin; ventral sclerite narrowing markedly in anterior end; mesosomal lobes roughly triangular, with sinuous lateral margins; rugose nodi small; 2 ames sensilla on each side near antero-lateral corners of mesosomal lobes; 2 pmes sensilla on each side of gonopore, near rugose nodi; gonopore oval, with slender marginal thickening ( Fig. 33 View FIGURES 31–35 ). Parameral heads roughly triangular and blades broad, tapering distally ( Figs 32, 34 View FIGURES 31–35 ). Measurements: Ex Dicrurus leucophaeus (n = 7): TL = 1.52–1.65; HL = 0.37–0.40; HW = 0.31–0.34; PRW = 0.20–0.22; PTW = 0.30–0.32; AW = 0.43–0.50.
Female. Thoracic and abdominal chaetotaxy as in Fig. 35 View FIGURES 31–35 ; psps present on tergopleurite VIII. Subgenital plate roughly rectangular, with concave lateral margins in distal half; lateral submarginal bulges broad, rounded; vulval margin rounded, with 3–4 short, slender vms on each side, and 4–5 short, thorn-like vss on each side; 3–6 short, slender vos on each side; distal 1 vos anterior to vss ( Fig. 35 View FIGURES 31–35 ). Measurements: Ex Dicrurus leucophaeus (n = 11): TL = 1.67–1.89 (1.79); HL = 0.41–0.43 (0.42); HW = 0.35–0.38 (0.36); PRW = 0.21–0.24 (0.23); PTW = 0.33–0.36 (0.34); AW = 0.45–0.52 (0.49).
Etymology. The species epithet derives from “ luridus ” Latin for “wan, pale”, referring to the comparatively pale tergal and sternal plates of the species.
Type material. Ex Dicrurus leucophaeus : Holotype ♂, Chiang Saen, Chieng Rai Province, Thailand, 25 Jan. 1965, H.E. McClure, SE-1890 ( NHML). Paratypes: 1♀, same data as holotype ( NHML); 2♂, 2♀, Pang Nam Un , Bun Yun, Nan Province, Thailand, 18 Jan. 1953, R. E. Elbel & H.G. Deignan, RE-2089, RT-B-12206 ( NHML); 1♂, 1♀, Chiang Saeh Kao , Chiang Rai Province, Thailand, 17 Feb. 1953, R. E. Elbel & H.G. Deignan, RE-2287, RT-B-17791 ( NHML); 1♂, 1♀, same data ( BPBM); 1♂, 1♀, Pang Nam Un , Bun Yun, Nan Province, Thailand, 26 Jan. 1953, R. E. Elbel & H.G. Deignan, RE-2209, RT-B-17733 ( BPBM); 1♂, 1♀, Hin Laem , Tha Khanun, Kanchanaburi Province, Thailand, 13 Nov. 1952, R. E. Elbel & H.G. Deignan, RE-1500, RT-B-13044 ( BPBM); 4♀, Chiang Saen Kao , Chiang Rai Province, Thailand, 17 Feb. 1953, R.E. Elbel & H.G. Deignan, RE-2287, RT-B-17791 ( USNM) .
Remarks. Guimaraesiella lurida was represented in the phylogeny of Bush et al. (2016) by one specimen from the type host and by one specimen from Geokichla citrina . Although both of these host species are known to participate in mixed-species foraging flocks ( Sridhar & Sankar 2008) with opportunities for louse exchanges in the wild, all other lice from G. citrina we have examined belong to other groups within Guimaraesiella . Therefore, we do not regard G. citrina as a natural and regular host of Guimaraesiella lurida until further specimens from this host confirm this host-louse association.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.