
Mycale (Oxymycale) klausjanusorum, Van Soest, 2018
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4466.1.6 |
|
publication LSID |
lsid:zoobank.org:pub:39B8D898-27B4-4BCE-A800-E729652C619B |
|
DOI |
https://doi.org/10.5281/zenodo.5951176 |
|
persistent identifier |
https://treatment.plazi.org/id/04138630-FF9C-FF88-4AB7-C8DEFD2D0E6F |
|
treatment provided by |
Plazi |
|
scientific name |
Mycale (Oxymycale) klausjanusorum |
| status |
sp. nov. |
Mycale (Oxymycale) klausjanusorum View in CoL sp. nov.
Figs 1a–d View FIGURE 1 , 2a–c View FIGURE 2 , 3a–f View FIGURE 3
Material examined. Holotype ZMA Por. 0 9452, Indonesia, Flores Sea, SW Salayar, E of Bahuluang, 6.485°S 120.45°E, depth 295–300 m, rectangular dredge, coll. J. van der Land, RV Tyro Snellius II Expedition, stat. 217, 12 October 1984.
Paratypes (detached fragments) ZMA Por. 0 9459, same data as holotype.
Description. ( Fig. 1a View FIGURE 1 ) Semiglobular massive encrustations on calcareous rubble, whitish with pale orange patches (in alcohol). Size of holotype 2 x 1 x 1 cm, of paratype detached fragments the largest of which is 3.5 x 1 x 0.5 cm. Surface markedly hispid. Consistency compressible, firm.
Skeleton. ( Figs 1b–d View FIGURE 1 , 2a View FIGURE 2 ) The choanosomal skeleton is strongly plumose, with thick spicule tracts present in the interior region (visible in Fig. 1d View FIGURE 1 ) about 500–600 µm in diameter, which towards the surface divide into thinner tracts of about 200 µm ( Figs 1b View FIGURE 1 , 2a View FIGURE 2 ). The subectosomal tracts partially carry a tangential ectosomal skeleton of 80 µm in thickness (visible in Figs 1b–c View FIGURE 1 , 2a View FIGURE 2 ) consisting predominantly of oxeas, but the tracts are also fanning out into bouquets of individual megascleres ( Figs 1b–c View FIGURE 1 , 2a View FIGURE 2 ), that protrude beyond the tangential ectosomal skeleton. The bouquets consist of longer style-like megascleres, differing in length and shape to the oxea-like megascleres from the tangential ectosomal skeleton. The choanosomal tracts appear to consist of a mixture of the two megasclere types. This bouquet-type of surface skeleton is unusual for Mycale species. Microscleres are confined to the choanosomal and tangential ectosomal skeleton, and are absent in the surface bouquets. Anisochelae I are predominantly arranged in rosettes ( Fig. 2a View FIGURE 2 ), with diameters of 300-400 µm, anisochelae II are individually distributed, relatively scarce, anisochelae III are crowded in irregular groups among the spicule tracts. Sigmas I are evenly and densely distributed, sigmas II are rather rare, and are individually distributed. Trichodragmas are densely distributed, the larger category most abundant, the smaller quite scarce.
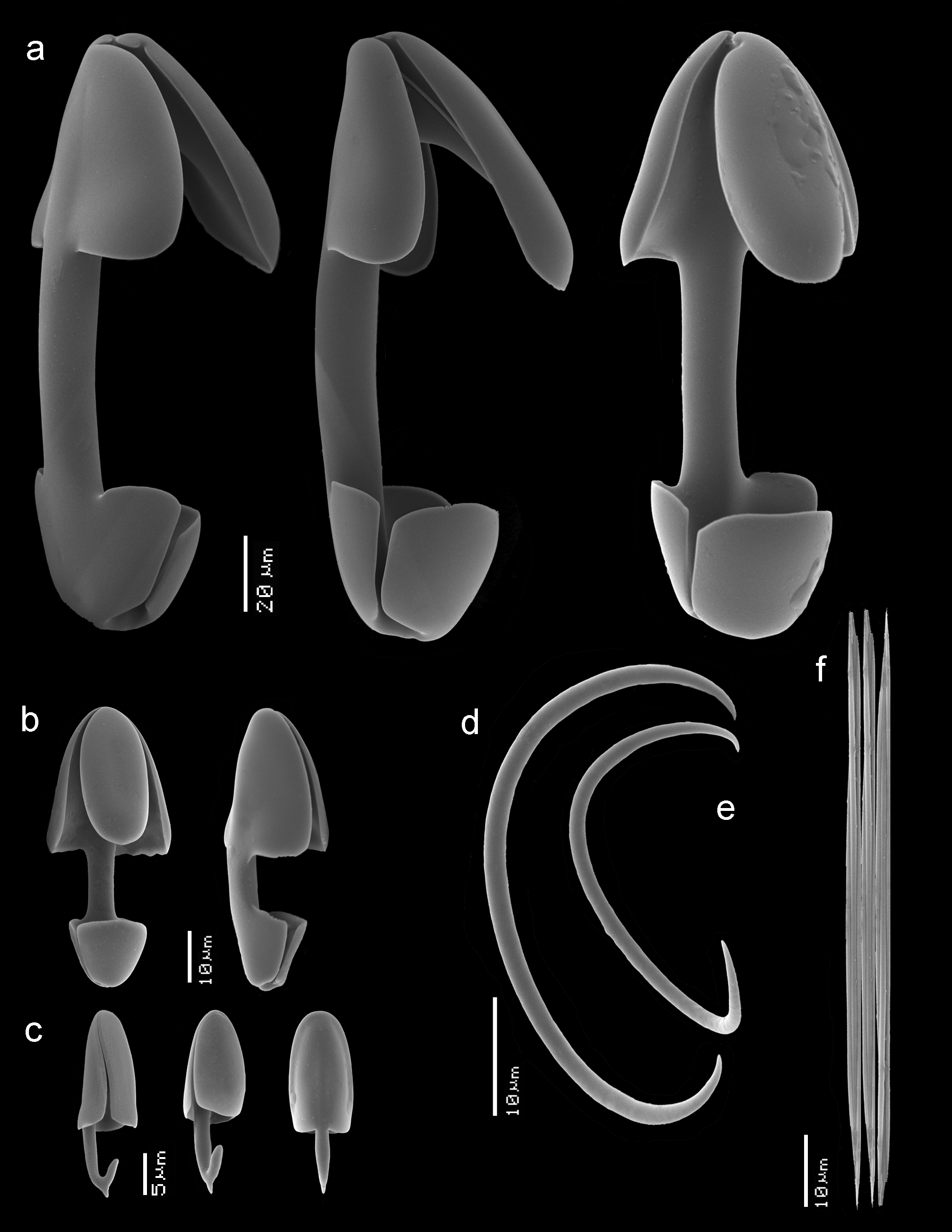
Spicules. ( Figs 2b–c View FIGURE 2 , 3a–f View FIGURE 3 ) Oxeote and stylote anisoxeas, three categories of anisochelae, two categories of sigmas, trichodragmas.
Oxeote megascleres ( Figs 2b,b View FIGURE 2 1 View FIGURE 1 ), at lower magnification optically symmetrical ( Figs 2b View FIGURE 2 ), but magnified ( Figs 2b View FIGURE 2 1 View FIGURE 1 ) they are slightly asymmetric with opposite more gradually pointed and more mucronated blunt endings, 522– 656 –732 x 14 – 16.8 –20 µm.
Stylote megascleres ( Figs 2c View FIGURE 2 ), with opposite gradually pointed and bluntly rounded end ( Figs 2c View FIGURE 2 1 View FIGURE 1 ), often with slightly narrow neck, 992– 1082 – 1175 x 19 – 20.7 –22 µm.
Anisochela I ( Figs 3a View FIGURE 3 ), robust, with flaring upper alae, naked shaft about one third of the total spicule length, 157– 172.6 –189 µm.
Anisochela II ( Figs 3b View FIGURE 3 ), rather similar in shape to anisochela I but with upper alae less flaring and shaft occupying only a quarter of the total spicule length, 33– 44.9 –57 µm.
Anisochela III ( Figs 3c View FIGURE 3 ), with upper alae close to the shaft and occupying more than half the length of the spicule, lower part reduced to a single spurred ala, 21– 24.0 –29 µm.
Sigma I ( Fig. 3d View FIGURE 3 ), predominantly strongly curved with thin sharp apices, 37– 44.7 –54 µm.
Sigma II ( Fig. 3e View FIGURE 3 ), similar in shape to the larger sigmas and close to overlapping in size, 16– 20.3 –27 µm.
Trichodragmas ( Fig. 3f View FIGURE 3 ), straight, possibly divisible in two size categories, but overlapping, the larger, very common, 78– 85.8 –107 x 14 – 20.6 –28 µm, the smaller, rare (only a few (n=4) were observed in the light microscopic slides, not shown), 24– 35.5 – 54 x 9 – 10.6 –12 µm.
Occurrence and ecology. Indonesia, between the islands of Salayar and Bahuluang, Flores Sea, on gravelly bottom at 295–300 m depth.
Etymology. It is my pleasure to name the new species combinedly to Klaus Rützler and Jean (‘Janus’) Vacelet in recognition of their life-long dedication to the science of sponges. Both were supportive and formative during my early sponge activities and over the years showed much valued friendship.
Remarks. The holotype and the paratype fragments are possibly part of a compound specimen, of which fragments became detached during the dredging operation. The uncertainty is the reason for separately registering two samples, distinguished as holotype and paratype.
I assembled the main features of all Mycale (Oxymycale) species in Tables 1a and 1b [one species, Oxymycale strongylata Pulitzer-Finali, 1986 is excluded because Hajdu & Desqeyroux-Faúndez (1994, p. 578) found it to be a junior synonym of Mycale (Mycale) laevis ( Carter, 1882) ]. From this table it can be concluded that the new species differs from all other Mycale (Oxymycale) in (1) having two distinct types of megascleres differing also in length, (2) in the greater length of the megascleres (up to 1175 µm, whereas the greatest length of the other species is 850 µm), (3) the ectosomal bouquets of single megascleres protruding beyond the tangential ectosomal skeleton, (4) the larger anisochelae I (up to 189 µm, whereas the largest anisochelae of the other species reach only up to 105–110 µm).
The closest species to the new species appears to be the one Sim & Kim (1988) (p. 25) described from South Korea as Oxymcale paradoxa De Laubenfels, 1935 . It shares the presence of oxea-like and style-like megascleres and overall habitus, but differs in spicule sizes (the two megasclere forms are not different in size, the larger anisochelae are only up to 115 µm). Details on the skeleton and the surface are not given. It is not conspecific with the California species, which is regarded as chimeric and not a Mycale species by Hajdu (1994). Apparently it concerns a Topsentia ( Halichondriidae ) or a Myrmekioderma (Heteroxyidae) contaminated with spicules of Mycale (Carmia) bellabellensis ( Lambe, 1905) . It is likely that a new name for the Korean specimen will have to be proposed.
Two other Oxymycale species have been described from South Korea, M. (O.) koreana ( Sim, 1982) and M. (O.) rhoi ( Sim & Lee, 1998) . The first is described as erect, and with skeleton Carmia -like without a proper ectosomal skeleton, megascleres are oxeas only, up to 539 µm, and anisochelae up to 88 µm, clearly unlike the new species. The latter Korean species is described as having a tangential ectosomal skeleton, oxea megascleres only, up to 660 µm, and anisochelae up to 110 µm, and thus similar to the new species. Nevertheless, the overall suite of characters is sufficiently distinct.
From the type species, Arctic M. (O.) intermedia ( Schmidt, 1874) (p. 433, fig. 10) as redescribed by Van Soest & Hajdu 2002 (based mostly on Thiele’s 1903b description of a slide from the Weltner collection, and on Koltun’s 1959 descriptions of 5 specimens, including the junior synonym Oxymycale wagini Koltun, 1952 ) the new species differs in the weak development of the skeleton of Schmidt’s species of which the ectosomal part is Carmia -like in having thin choanosomal tracts and no mention of a tangential spicule arrangement, and in the apparent lack of sigmas and trichodragmas. Megascleres and anisochelae I are about half as large as those of M. (O.) klausjanusorum sp. nov. Like the latter, M. (O.) intermedia is reported from depths down to 325 m. The species is not well-known.
Further species assigned to Mycale (Oxymycale) are progressively more unlike the new species: South African M. (O.) stephensae Samaai & Gibbons, 2005 lacks trichodragmas. Antarctic M. (O.) acerata Kirkpatrick, 1907 and M. (O.) tylotornota Koltun, 1964 lack sigmas, Western Pacific M. (O.) strongylophora ( De Laubenfels, 1954) has strongyles for megascleres. Western Pacific M. (O.) stecarmia ( De Laubenfels, 1954) has very small oxeas (up to 154 µm) for megascleres, the anisochelae are only up to 42 µm, it may possess toxas (not considered proper), together making it a very unlikely relative of the new species. Finally, Florida M. (O.) renieroides ( Schmidt, 1870) cannot be properly assessed as a Mycale according to Hajdu & Rützler (1998) after examination of a slide in the Natural History Museum made by Schmidt. The latter species is apparently still represented by type material, both dried and in alcohol, in the collection of the Museum of Comparative Zoology, Harvard, USA.
Similarities with the new species are evident in Mycale (Mycale) longistyla Koltun, 1958 from 132–270 m depth in the North Pacific, where the megascleres protrude far beyond the tangential ectosomal skeleton. These megascleres consist of three categories, large styles of up to 3.5 mm x 32 µm, intermediate styles of 1144– 1768 x 23 –32 µm, and small strongyles of 416–530 x 9–11 µm, differentiated in a way reminding of the megascleres of the new species, but upper sizes are clearly in excess of the latter. Assignment of this species to the subgenus Mycale (Oxymycale) may be contemplated (but see below).
Comparison is also warranted with several species not currently assigned to the subgenus Mycale (Oxymycale) showing skeletal similarities with the new species.
Mycale (Mycale) topsenti Burton, 1959 View in CoL from 1046 m depth off the coast of Oman shares the structure of the surface skeleton with the new species, with ‘diffuse brushes’ of megascleres projecting beyond the tangential ectosomal skeleton. Megascleres are clearly different in consisting of a single size of subtylostyles of 900 x 18 µm, and among the microscleres the largest anisochelae are up to 88 µm. The skeletal properties described for New Caledonian Mycale (Mycale) myriasclera Lévi & Lévi, 1983 View in CoL , from 300 m depth, include also megascleres protruding beyond the tangential ectosomal skeleton, but a clear difference is the absence of sigmas in the New Caledonian species.
Several Mycale View in CoL species with anisochelae I of the same size or larger as those of the new species reported from the Indo-West Pacific region were also reviewed, e.g. Mycale (Mycale) grandis Gray, 1867 View in CoL , Mycale (Mycale) anisochela Lévi, 1963 View in CoL . However, these species differ clearly in the structure of the surface skeleton, the shape of the megascleres and the additional microscleres.


| ZMA |
Universiteit van Amsterdam, Zoologisch Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Mycale (Oxymycale) klausjanusorum
| Van Soest, Rob W. M. 2018 |
Mycale (Mycale) myriasclera Lévi & Lévi, 1983
| Levi & Levi 1983 |
Mycale (Mycale) anisochela Lévi, 1963
| Levi 1963 |
Mycale (Mycale) topsenti
| Burton 1959 |
Mycale
| Gray 1867 |
Mycale (Mycale) grandis
| Gray 1867 |