
Othelosoma duplamaculosum Sluys & Neumann
|
publication ID |
https://doi.org/ 10.5281/zenodo.250240 |
|
publication LSID |
lsid:zoobank.org:pub:B0B97269-AE03-4A41-811A-5F135988055E |
|
DOI |
https://doi.org/10.5281/zenodo.6022394 |
|
persistent identifier |
https://treatment.plazi.org/id/06116D51-FFC2-844B-FF02-FB2EFDD6FC3D |
|
treatment provided by |
Plazi |
|
scientific name |
Othelosoma duplamaculosum Sluys & Neumann |
| status |
sp. nov. |
Othelosoma duplamaculosum Sluys & Neumann , sp. nov.
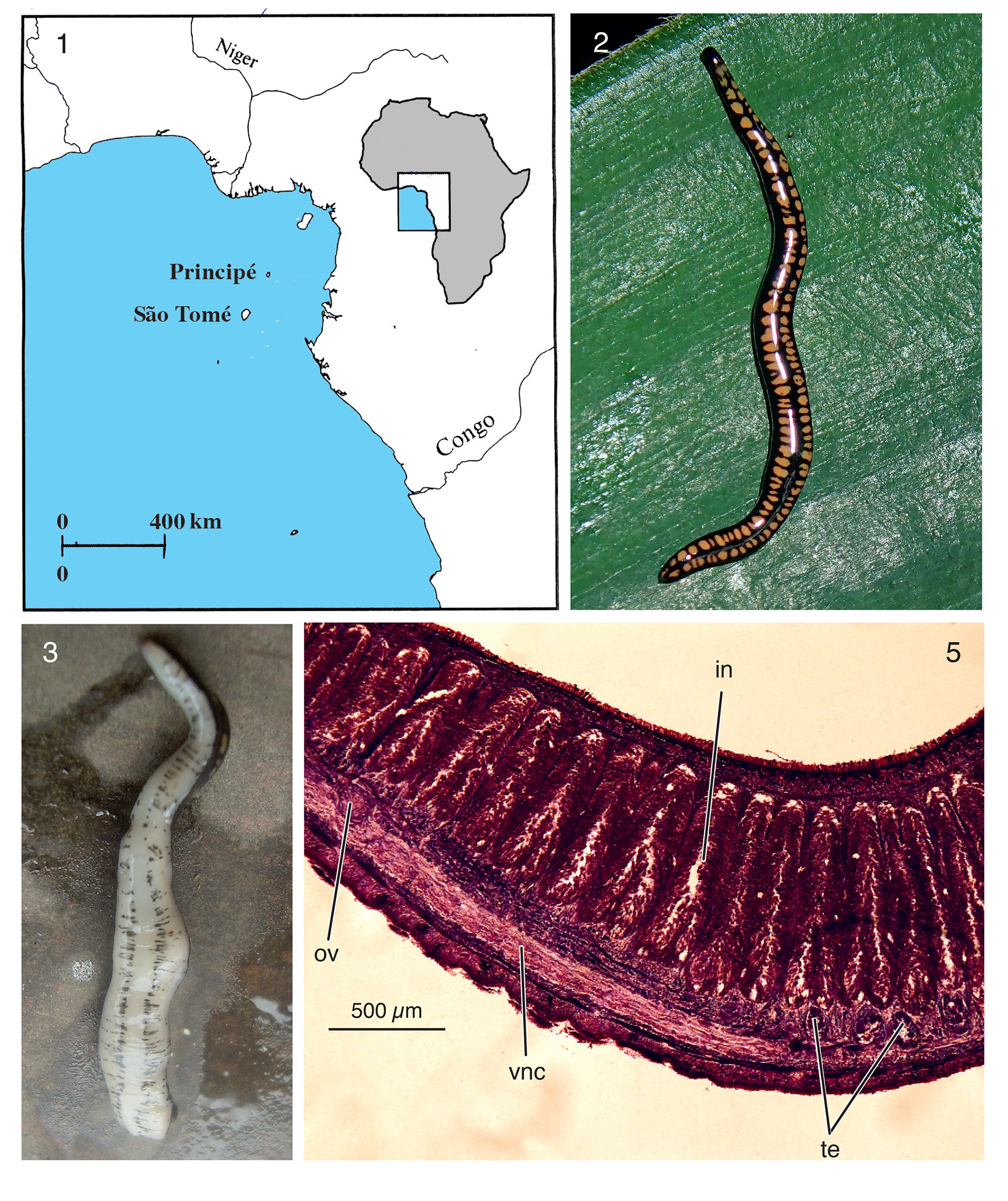
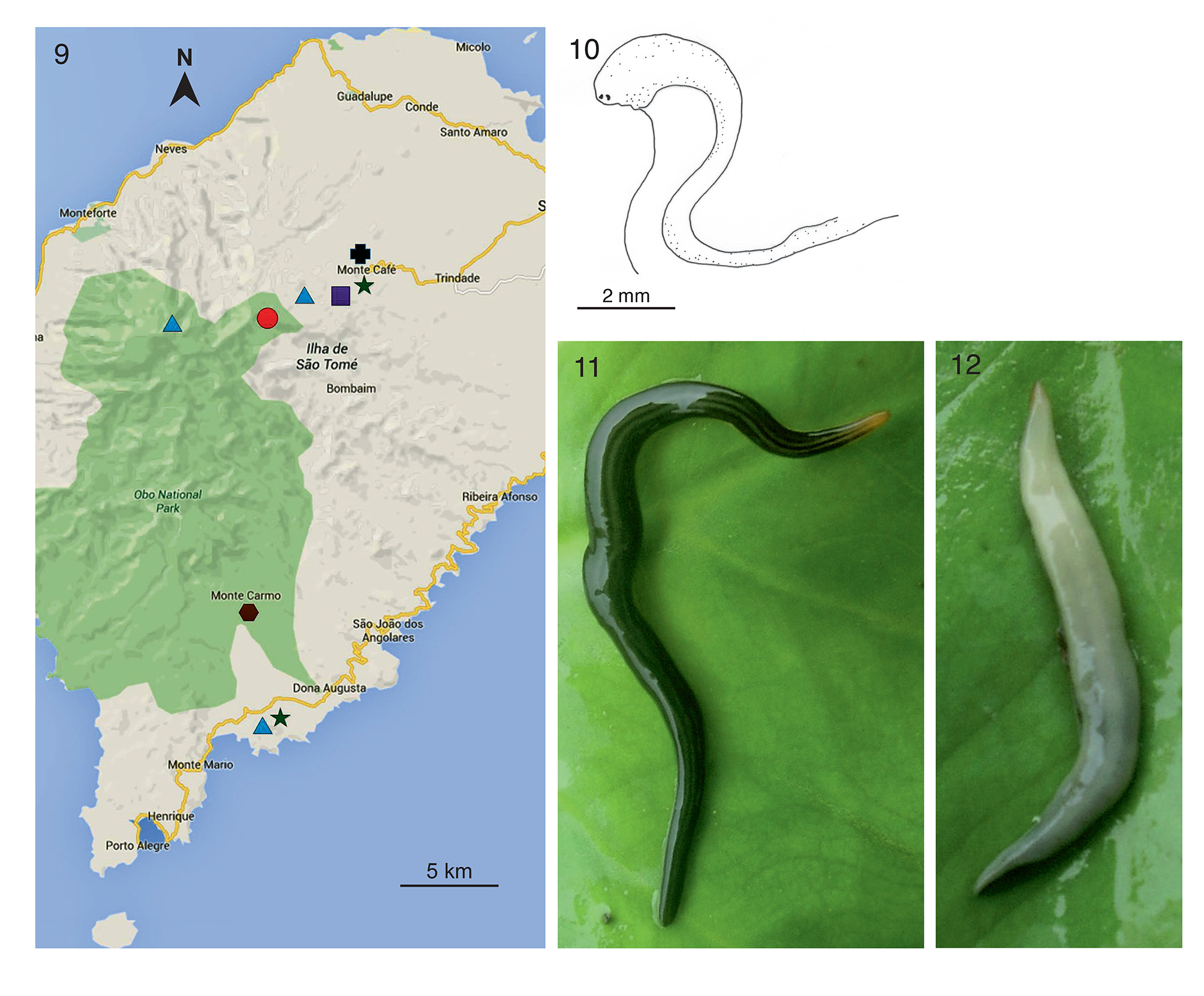
Material examined. Holotype: ZMA V.Pl. 7247.1, São Tomé Island, Bom Sucesso , Monte Café, Mé Zóchi district, 18 November 2014, coll. Leonel Viegas, sagittal sections on 7 slides of the anterior part; sagittal sections on 20 slides of the pharynx and copulatory apparatus; sagittal sections on 11 slides of the tail end.
Other material: ZMA V.Pl. 7248.1, São Tomé Island, Pica do São Tomé, N0.273325 E6.547972, alt. 1725m, 11 April 2012, coll. Miko Nadel, sagittal sections of the front end on 4 slides; transverse sections of the copulatory apparatus on 12 slides; sagittal sections of the pharynx region on 10 slides; sagittal sections of the posterior end on 9 slides. GoogleMaps
ZMA V.Pl. 7249.1, São Tomé Island, Bom Sucesso , March 2015, coll. Leonel Viegas, sagittal sections of the anterior end on 12 slides; horizontal sections of the pharyngeal region on 6 slides; sagittal sections of the copulatory apparatus on 11 slides; sagittal sections of the tail end on 6 slides.
Etymology. The specific epithet is derived from the Latin duplus, twofold, and maculosus, spotted, dappled, and alludes to the fact that this species shows rows of spots on both dorsal and ventral body surface.
Diagnosis. Black Othelosoma species with two rows of irregular yellow-ochre spots on the dorsal surface. Two rows of black specks, forming bands, present on the dirty white ventral body surface. No retractor muscle in the anterior part of the body. Communication of the bursal canal with the vaginal duct located halfway between the atrial and bursal openings of the latter. Vaginal duct extending far into the small, thick-walled copulatory bursa.
Description. Dorsal surface black with rather large, irregular yellow-ochre spots arranged in two rows ( Fig. 2 View FIGURES 1 – 3, 5 ). Ventral surface dirty white and with two rows of irregular black specks, forming two bands; irregular rows of specks also run from each of the bands towards the lateral side of the body ( Fig. 3 View FIGURES 1 – 3, 5 ). A pair of rather large eyes is situated at the very anterior end of the body. The preserved specimen ZMA V.Pl. 7247.1 measured 23mm in length and 2.3mm in width. Narrow creeping sole, covering about one-third of the total width of the body.
Subepidermal musculature consisting of a thin layer of circular muscle, followed by a thin layer of longitudinal muscle. Parenchymal longitudinal muscles well developed, particularly on the ventral side, where longitudinal muscles are present also dorsally to the ventral nerve cords. No retractor muscle in the anterior end of the animal.
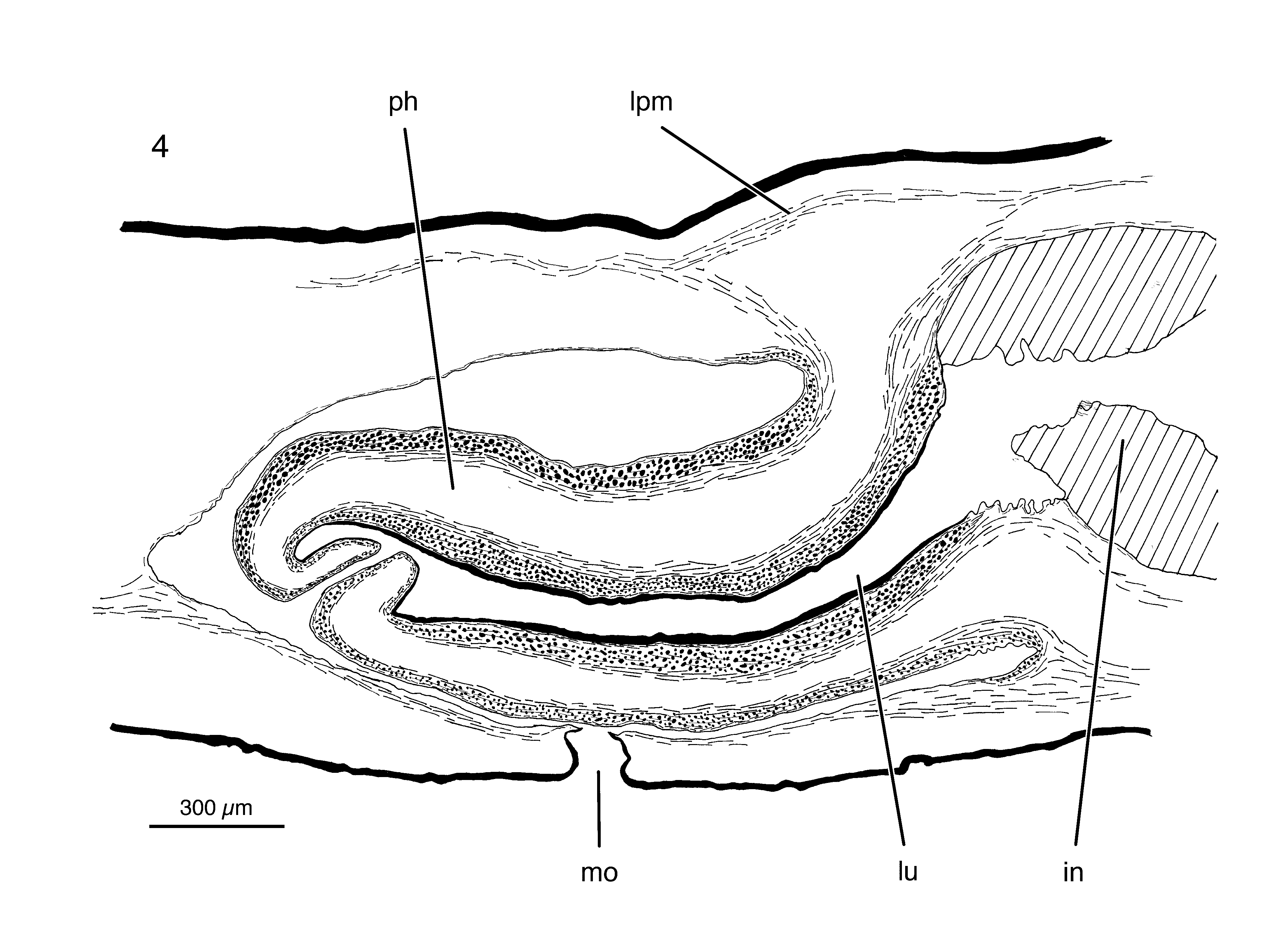
The root of the cylindrical pharynx is placed more or less halfway of the distance between the anterior tip and the posterior end of the specimen. Pharynx length is almost 1/13th of the total body length in the preserved holotype specimen. Mouth located halfway between the anterior and the posterior tip of the holotype specimen. Mouth opening located about halfway between the posterior wall of the pharyngeal cavity and the root of the pharynx ( Fig. 4 View FIGURE 4 ). Pharyngeal pouch musculature composed of few layers of subepithelial longitudinal muscles.
Outer pharynx epithelium underlain by a simple layer of longitudinal muscles, followed by stronger layers of intermingled muscles with both circular and longitudinal fibres. The same configuration of muscle layers is present underneath the inner pharynx epithelium, lining the lumen, albeit that here the zone of muscles is thicker than the one underneath the outer epithelium ( Fig. 4 View FIGURE 4 ).
In the holotype the first testes are located 0.8mm behind the ovaries and 2.3mm behind the centre of the brain. The testes extend backwards to about 700µm posterior to the mouth opening. The testes are located ventrally on either side of the body between the intestinal branches and the sub-intestinal transverse parenchymal muscle layer. The shape of the testes follicles is oblong, the follicles occupying less than 1/7th of the dorsoventral diameter in the prepharyngeal and the pharyngeal part of the body (Fig. 5).
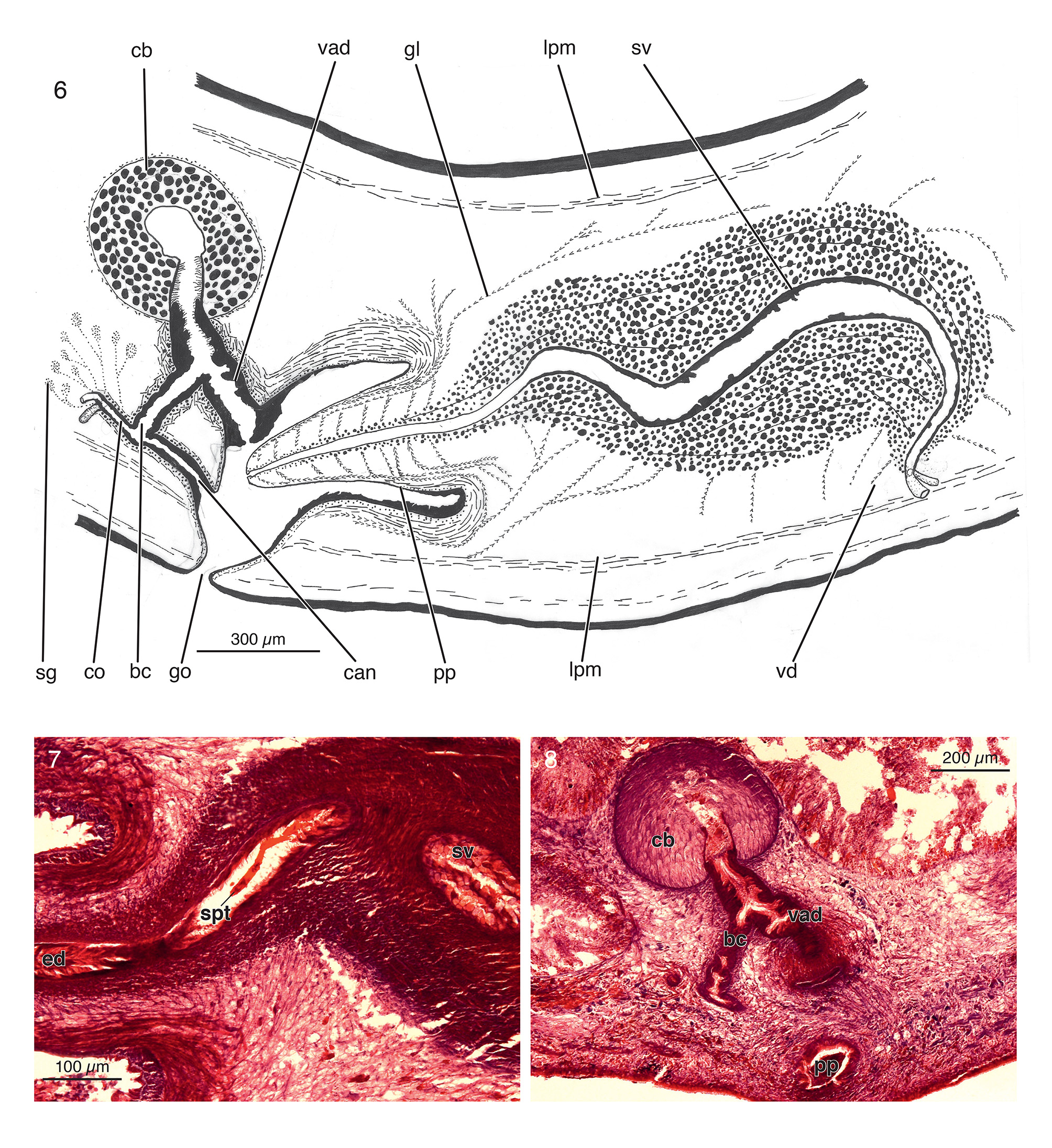
The sperm ducts run amongst the fibers of the parenchymal muscle layer located dorsally to the ventral nerve cords. The distribution of these muscle fibers around the duct is irregular, being more abundant on the dorsal than on the ventral side. The sperm ducts expand very slightly and turn dorsally under the most antero-ventral part of the seminal vesicle. At this point the sperm ducts fuse to form a short common sperm duct, which penetrates the coat of muscles around the seminal vesicle. The ovoid, elongated seminal vesicle has a relatively narrow lumen and runs more or less parallel to the body surface. After having received the opening of the common vas deferens, the Sshaped lumen initially expands somewhat in diameter and, subsequently, gradually narrows to form the ejaculatory duct, which opens at the tip of the penis papilla ( Fig. 6 View FIGURES 6 – 8 ). The latter is a short conical structure with a blunt tip and it is covered with a thin, nucleated epithelium. In the holotype the penis papilla proper, i.e. from its point of insertion to the tip, measures 500µm, while the seminal vesicle measures 1300µm in length. The seminal vesicle is surrounded by a very strong layer of circular muscles. Longitudinal muscles lie interspersed in this layer of circular muscles. The posterior part of the seminal vesicle contains a more than 10µm long part of a sclerotic spermatophore ( Fig. 7 View FIGURES 6 – 8 ), while remnants of a spermatophore are also present at the opening of the ejaculatory duct at the penis tip and in the male atrium.
Penis glands are mainly distributed dorsally and ventrally to the root of the penis papilla. Furthermore, the penis papilla receives secretions from extra-penial glandular elements from the area around the posterior half of the seminal vesicle ( Fig. 6 View FIGURES 6 – 8 ). These erythrophil glands discharge, perpendicularly to the anterior-posterior body axis, into the the ejaculatory duct and the lumen of the seminal vesicle, or prostatic vesicle. Further there are many of these glands around the anterior part of the seminal vesicle discharging secretions into the latter.
Genital atrium lined with a thin, nucleated epithelium, underlain with a subepithelial circular muscle layer, followed by longitudinal muscles.
Gonopore located at two-thirds of the distance between the anterior and posterior end of the specimen. Distance between mouth and gonopore approximately 4.6mm, i.e. 1/5th of the total body length in the preserved holotype specimen.
Ovaries situated above the ventral nerve cords, occupying 1/6th of dorsoventral diameter of the anterior part of the body ( Fig. 7 View FIGURES 6 – 8 ). They are positioned at 1.4mm behind the brain. The oviducts arise from ventral side of ovaries. Oviducts run in posterior direction between the fibers of the parenchymal muscle band. The dispersion of the muscle fibers around the ducts is irregular, more fibres being present to the dorsal side than to the ventral side. Posterior to the gonopore, at the level of the bursa, the oviducts turn dorsally and, subsequently, fuse to form a short common oviduct ( Fig. 6 View FIGURES 6 – 8 ). The latter opens into the bursal canal at a position about halfway between the opening of the canal into atrium and its communication with the bursa. The oviducts receive shell glands before and after they have fused to form a common oviduct. The cell bodies of these erythrophil shell glands are distributed dorsally to the common oviduct, while they also extend quite far into posterior direction.
From the point of the opening of the common oviduct, the bursal canal of the holotype rises obliquely for about 200µm into antero-dorsal direction to open into the vaginal duct (this part corresponds to Beauchamp's canal); in the other direction it runs obliquely towards the atrium (this section corresponds to the canalis anonymus). The communication between the bursal canal and the vaginal duct is at about halfway between the opening of latter into the bursa and its communication with the atrium, which is located about 310µm dorsally to the ventral surface.
The vaginal duct penetrates the ventral surface of the copulatory bursa and extends in the holotype for about 110µm into the latter ( Fig. 8 View FIGURES 6 – 8 ). This extension is mostly composed of a very flat section of the epithelium, bearing well developed cilia, as does the entire vaginal duct ( Fig. 6 View FIGURES 6 – 8 ).
Vaginal duct and bursal canal are lined with a nucleated, ciliated, and pseudostratified epithelium. The musculature around both canals, vaginal duct and Beauchamp's canal, consists of a layer of circular muscles followed by a layer of longitudinal muscles, which are partly diagonally, criss-cross arranged. This zone of muscles is slightly more developed on the vaginal duct than on the Beauchamp's canal. The canalis anonymus section of the bursal canal is surrounded by a thin layer of circular muscles.
The ball-shaped bursa of the holotype measures about 400µm in diameter and occupies approximately 30% of the dorso-ventral diameter of the body in this part of the specimen. The bursa is lined with a thick-walled (100- 150µm), pseudo-stratified epithelium ( Fig. 8 View FIGURES 6 – 8 ). The lumen of the bursa is small, less than 120µm in diameter. The bursal musculature is composed of a weak layer of decussate muscle fibres.
Distribution. The species is known from the type locality, Bom Sucesso, Monte Café, where it was collected on two different occasions, and also from another location on São Tomé Island, viz. Pica do São Tomé. A third locality where the species was observed, but from where no specimens were collected, is Ribeira Peixe, around 25km south of the type locality near Monte Café ( Fig. 9 View FIGURES 9 – 12 ).
Discussion. The unique feature, among the known species of the genus Othelosoma , of irregular spots arranged in two rows on the dorsal as well as on the ventral body surface make it easy to distinguish O. duplamaculosum from all other presently known representatives of the genus Othelosoma . Generally, most Othelosoma species have a striped body. Below we will compare O. duplamaculosum with all other Othelosoma species that have at least one dark dorsal stripe or band.
The combination of the following internal, anatomical features in O. duplamaculosum invites comparison with only three of the known Othelosoma species, viz. O. caffrum (Jameson, 1907) , O. huntum Marcus, 1955 , and O. wauzen Marcus, 1955 : (1) bursal canal fusing with the vaginal duct halfway between the atrial and bursal openings of the latter, and (2) presence of a small, thick-walled bursa that occupies not more than one third of the dorsoventral diameter of the preserved specimen in that part of the body. The bursal canal in these three species fuses with the vaginal duct more or less halfway before the latter enters into the small copulatory bursa ( Marcus 1955a). In other words, the only bursal opening is the opening of the vaginal duct into the bursa, the bursal canal fusing with the vaginal duct well before this point. Furthermore, all three species have a common oviduct, as does O. duplamaculosum .
However O. huntum and O. wauzen differ from O. duplamaculosum in the presence of intra- and extrabulbar vesicles of the male duct ( Marcus 1955a), whereas the lumen of the seminal vesicle of O. duplamaculosum gradually narrows to form the ejaculatory duct without forming intra- or extrabulbar vesicles. Further, in O. wauzen the common atrium is separated from the male atrium by a muscular sphincter ( Marcus 1955a), which is absent in O. duplamaculosum . In contrast to O. duplamaculosum , in which the lumen of the seminal vesicle is S-shaped, in O. huntum the whole muscular seminal vesicle, and its lumen, is loop-shaped ( Marcus 1955a).
In addition to these anatomical differences between the copulatory organs of O. duplamaculosum and O. huntum and O. wauzen mentioned above, they do also differ in another feature. In O. wauzen , and also in O caffrum , there is a ventral retractor muscle, whereas O. huntum shows a dorsal retractor muscle in the anterior body end ( Marcus 1955a). However, Othelosoma duplamaculosum completely lacks retractor muscles in the anterior end of the body.
The course of the muscular seminal vesicle is similar in O. duplamaculosum and O. caffrum , but the shape of the penis papilla is different. While in O. caffrum the vertically orientated penis papilla has a broad basis, which quickly narrows towards the tip ( Marcus 1955a), the more horizontally orientated penis papilla of O. duplamaculosum has the shape of a blunt cone, which diminishes in diameter much more gradually. Furthermore, a muscular sphincter around the vaginal duct is present in O. caffrum ( Marcus 1955a) , whereas such a structure is absent in O. duplamaculosum .
It is noteworthy that in specimen ZMA V.Pl. 7247.1 of O. duplamaculosum the posterior fifth of the seminal vesicle contains a more than 10µm long part of a sclerotic spermatophore, while remnants of a spermatophore are also present at the opening of the ejaculatory duct at the tip of the penis papilla and in the male atrium. To the best of our knowledge this is the first record of a sclerotic spermatophore in an Othelosoma species. In general, such sclerotic spermatophores are rare in terrestrial planarians and thus far have only been recorded for Platydemus victoriae ( Heinzel, 1929) and within the copulatory bursa of Pimea monticola Winsor, 1991 , Microplana aixandrei Vila-Farré et al., 2008 and have been suggested for Microplana hyalina Vila-Farré et al., 2011 ( Winsor 1991; Vila- Farré et al. 2011, and references therein).
| ZMA |
Universiteit van Amsterdam, Zoologisch Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Microplaninae |
|
Genus |