
Dysmicoccus lavandulae Germain, Matile-Ferrero & Williams
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3980.4.7 |
|
publication LSID |
lsid:zoobank.org:pub:F0AE0356-EF0D-4DDC-94AA-5046D7F86F51 |
|
DOI |
https://doi.org/10.5281/zenodo.6098332 |
|
persistent identifier |
https://treatment.plazi.org/id/061D87E7-055F-FFA9-FF2A-F8E039B6FB52 |
|
treatment provided by |
Plazi |
|
scientific name |
Dysmicoccus lavandulae Germain, Matile-Ferrero & Williams |
| status |
sp. nov. |
Dysmicoccus lavandulae Germain, Matile-Ferrero & Williams n. sp.
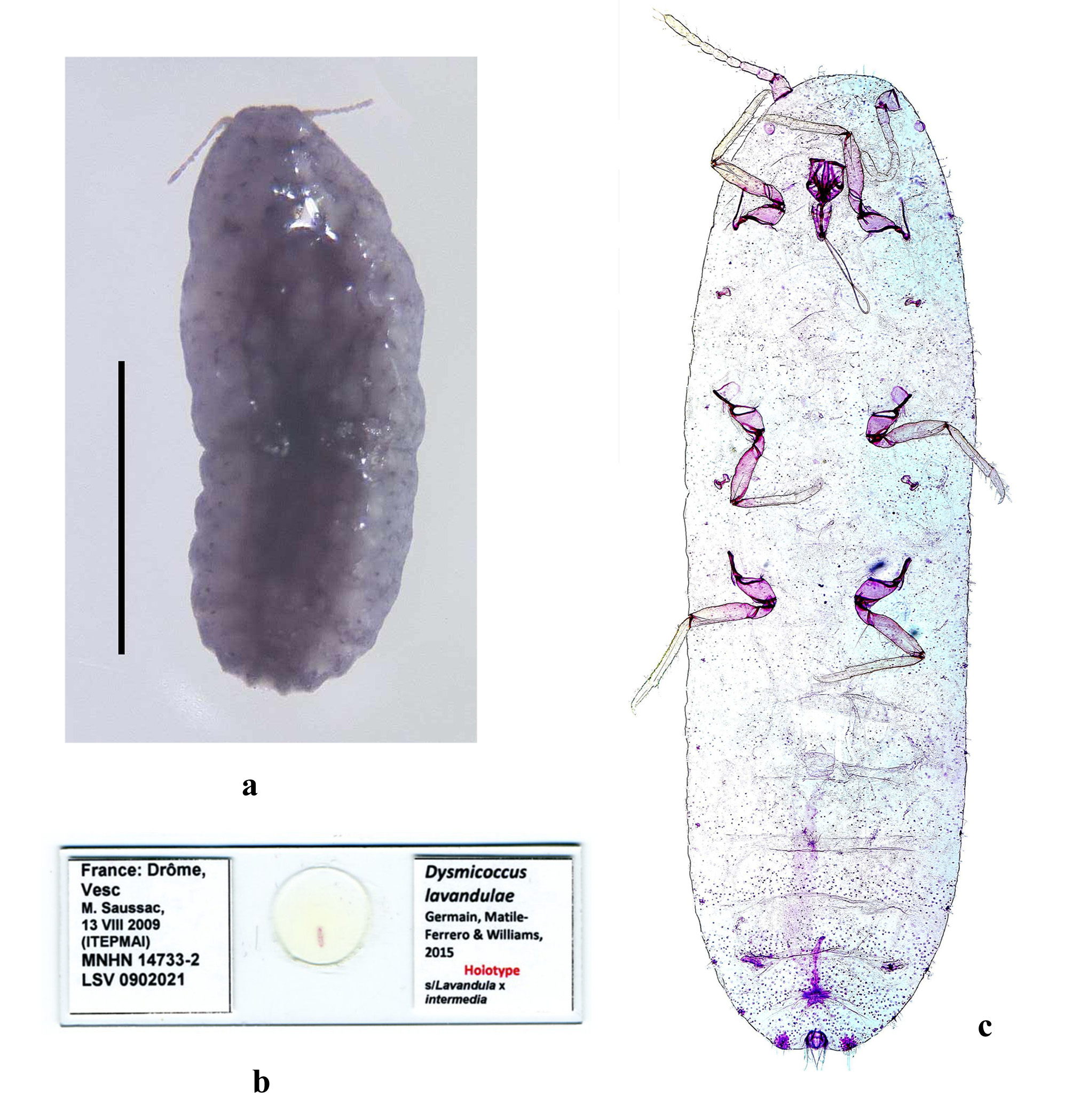
(fig. 2)
Unmounted material. Body of live specimens greyish, devoid of wax with 2 short caudal wax filaments. Purplish blue in alcohol 70° ( Fig. 3 View FIGURE 3 a), becoming immediately brown in cold KOH and then reddish brown 3 hours later.
Slide-mounted adult female ( Fig. 3 View FIGURE 3 b and 3c) elongate oval, 1.44–2.96 mm long; anal lobes moderately developed, each with an apical seta 120–180 µm long. Antennae each 350–450 µm long, with 8 segments. Legs well developed; hind trochanter + femur 220–317 µm long, hind tibia + tarsus 280–410 µm long, claw about 2430 µm long. Ratio of lengths of hind tibia to tarsus 2.7. Ratio of lengths of hind tibia + tarsus to hind trochanter + femur 1.29. Translucent pores absent from hind coxa. Labium 85–125 µm long, shorter than clypeo-labral shield. Circulus present, variable in size and shape, elliptical to quadrate, 30–130 µm wide. Ostioles well developed, with inner edges of lips sclerotized, each lip with 3–5 setae and a few trilocular pores. Anal ring about 67–81 µm wide, with 6 setae, each 95–137 µm long. Cerarii numbering 12 (10–13) definite pairs. Anal lobe cerarii each with 2 conical setae, each 19–24 µm long, about 6 auxiliary setae and a group of trilocular pores. Anterior cerarii each with 2 shorter conical setae and 1 or 2 auxiliary setae.
Dorsal surface with slender setae ranging in length from 10 to 29 µm. Multilocular disc pores absent.
Trilocular pores evenly distributed, each about 4 µm wide. Discoidal pores minute, very sparse, each about 2.6 µm wide. Oral collar tubular ducts of 2 sizes, smaller ducts each 2–3.5 µm in diameter, larger ducts 3.3–5 µm in diameter; smaller ducts much more numerous and mainly forming transverse rows on last abdominal segments; larger ducts sparsely distributed among smaller ducts but most numerous on abdominal submargins.
Ventral surface with slender flagellate setae, these becoming shorter towards margins. Multilocular disc pores, each about 6.4–8.7 µm in diameter, present posterior to vulva and on segments VI and VII but never reaching margins. Trilocular pores as on dorsum, evenly distributed. Minute discoidal pores sparse. Oral collar tubular ducts, of 2 sizes, as on dorsum but less numerous.
Material examined. Holotype: adult female, FRANCE, Drôme, Vesc, on Lavandula x intermedia, M. Saussac, 13.VIII.2009, MNHN 14733-2, LSV 0 902021 ( MNHN).
Paratypes: same data as holotype, 3 adult females on 3 slides (Anses-LSV, 1 slide, BMNH 1 slide; MBK 1 slide); Drôme, Dieulefit, on Lavandula x intermedia, R. Bonnaure, 26.VI.2010, 12 adult females on 5 slides (Anses-LSV: 2 slides with 5 adult females; BMNH 1 slide with 2 adult females; MBK 1 slide with 2 adult females; MNHN 1 slide with 3 adult females); same data, R. Bonnaure, 13.V.2011, 2 slides with 1 adult female and 6 2ndinstar females) (Anses-LSV); same data, R. Bonnaure, 31.V.2011, 5 adult females on 5 slides (Anses-LSV); same data, R. Bonnaure, 31.V.2011, 6 adult females on 6 slides (Anses-LSV 3 slides; BMNH 1 slide; MBK 1 slide; MNHN 1 slide); same data, R. Bonnaure, 8.VI.2011, 2 adult females on 2 slides (Anses-LSV); Dieulefit, Marroux, on L. x intermedia, D. Matile-Ferrero & J.-F. Germain, 16.VI.2011, 1 adult female ( MNHN).
Molecular characterization. After the sequence analysis, the consensus sequences obtained for 28S, ITS2 and COI were of 763 bp, 893 bp and 1045 bp, respectively. Intraspecific variation was observed in COI only: two haplotypes differing by two substitutions were observed in two and three specimens, respectively.
The consensus sequences were submitted to the NCBI database under the accession numbers KR340586 View Materials (28S), KR340587 View Materials (ITS2), KR340584 View Materials ( COI haplotype 1) and KR340585 View Materials ( COI haplotype 2).
The sequences obtained for COI and 28S were compared to sequences already available for other Dysmicoccus and Trionymus species (Kaydan et al. 2015): Dysmicoccus multivorus (the species most closely related to D. lavandulae ), Trionymus aberrans , Trionymus perrisi and Trionymus artemisiarum . Because the PCR primers used in this work and those used by Kaydan et al. (2015) were different, comparisons could only be made on the 380 bp for COI and 306 bp for 28S DNA regions.
The sequence comparison, after alignment ( Fig 4 and 5), reveals the extent of divergence corresponding to interspecific variations in previous studies ( Malausa et al. 2011; Park et al. 2011). At COI, the divergence was>7% between D. lavandulae and D. multivorus and>9% between D. lavandulae and the Trionymus species. At 28S, the divergence was>2% between D. lavandulae and D. multivorus and>8% between D. lavandulae and the other Trionymus species. From these results, it is concluded that the new species clearly has own linage and different from all other related species.
Comments. The new species is placed in Dysmicoccus Ferris and belongs to a group with a reduced number of cerarii and two sizes of oral collar tubular ducts on the dorsum and venter. This group differs from Trionymus Berg , a genus here regarded as restricted to species with only 1–3 pairs of cerarii. In usually possessing 12 (range 10-13) pairs of cerarii, D. lavandulae is similar to D. pietroi Marotta (Marotta, 1992) , a species possessing 13 pairs of cerarii but which also has ventral multilocular disc pores on segment V (absent on this segment in D. lavandulae ), and noticeably more tubular ducts on both the dorsum and venter. Dysmicoccus pietroi was described from Italy, Campania, on unidentified Poaceae and on Cirsium sp. ( Asteraceae ). Dysmicoccus lavandulae is also similar to some species presently included in Trionymus in ScaleNet, a database of the scale insects ( Ben-Dov et al. 2015). Pending further research, we believe that D. lavandulae is also related to D. multivorus (Kiritchenko) , described originally as Pseudococcus multivorus by Kiritchenko (1936), now a widespread species in Europe (and recorded from France by Panis (1999) and neighbouring parts of Asia. However, D. multivorus , as discussed by Ter- Grigorian (1973), Tereznikova (1975) and Danzig (1997), usually possesses only 4–6 pairs of cerarii.
The new species is also similar to Trionymus angustifrons Hall , described from Egypt on Ambrosia maritima (Asteraceae) by Hall (1926), and recorded from south-eastern France, by Goux (1941). Specimens are at hand prepared by Goux (MNHN), labelled Marseille, La Madrague-de-Montredon, on Compositae, 19.II.1939, coll. L. Goux. The species has also been recorded from Saudi Arabia on Rhazya stricta (Apocynaceae) at Rhyadh, and on Tamarix aphylla (Tamaricaceae) from Jizan by Matile-Ferrero (1988). In addition, it has also been recorded from Israel, on Carthamus glauca (Asteraceae) from Rehovot and on Echinops viscosus (Asteraceae) from Qazrin by Ben-Dov (1991). Trionymus lanatus (Balachowsky) , described from France, Port-Cros Island on Kentrophyllum lanatum (Asteraceae) by Balachowsky (1932), also possesses 5 or 6 pairs of cerarii and belongs to the same group. It is hoped to study the status of these species more in the near future.
10 20 30 40 50 60 70
.... | .... | .... | .... | .... | .... | .... | .... | .... | .... | .... | .... | .... | .... | .... | ... Dysmicoccus lavandulae (HaP1) TCCAGAAGTTTATATTCTAATTTTACCAGGATTTGGAGCTATATCCCAAATTATAAATCAAGAAAGGGGAAAAATTGAA Dysmicoccus lavandulae (HaP2) .............................................T....................A........... Dysmicoccus multivorus (MBK047) ...T..................C.............T..A.....T....................T........... Trionymus Perrisii (MBK045) ...T.....A........T...C.C.....T...........G..T...........C........T..T.....A..
90 100 110 120 130 140 150
.... | .... | .... | .... | .... | .... | .... | .... | .... | .... | .... | .... | .... | .... | .... | ... Dysmicoccus lavandulae (HaP1) TTTTCAGTAAAATTAATATAATTTTTGCTATAATTTCAATTGGAATTTTAGGATTCATTGTTTGAGCCCATCATATATT Dysmicoccus lavandulae (HaP2) .............................................................................. Dysmicoccus multivorus (MBK047) ....T......................................T........T..T...................... Trionymus Perrisii (MBK045) ....T...............................................T..T...........T.....C....
170 180 190 200 210 220 230
.... | .... | .... | .... | .... | .... | .... | .... | .... | .... | .... | .... | .... | .... | .... | ... Dysmicoccus lavandulae (HaP1) ACTATTGGTTTAGACATTGATACACAATTATACTTTATATCAGCAACTATAATTATTGCTATTCCAACAAGAATTAAAA Dysmicoccus lavandulae (HaP2) .............................................................................. Dysmicoccus multivorus (MBK047) ..............T.................T........T..T.................C............... Trionymus Perrisii (MBK045) ..............T.....C......C.T..T..C.....C..T..A...........A..C...........C...
250 260 270 280 290 300 310
.... | .... | .... | .... | .... | .... | .... | .... | .... | .... | .... | .... | .... | .... | .... | ... Dysmicoccus lavandulae (HaP1) TTTCAGTTGAATAATAACTTTAAATGGAAAAAAAATCTTAAATTCATCTATTTCATTATGATCAATAGGTTTTATTATT Dysmicoccus lavandulae (HaP2) .............................................................................. Dysmicoccus multivorus (MBK047) ...T................................T..T.....T..A..C..............T........... Trionymus Perrisii (MBK045) ...T..A.............................T........T..A....AT........T..T..A........
330 340 350 360 370 380
.... | .... | .... | .... | .... | .... | .... | .... | .... | .... | .... | .... |
Dysmicoccus lavandulae (HaP1) TATTTACATTAGGAGGATTAACAGGAATTATTTTATCTAATTCAATTATCGATATTAATC
Dysmicoccus lavandulae (HaP2) ............................................................
Dysmicoccus multivorus (MBK047) .....T.T........CC...............................T.....C....
Trionymus Perrisii (MBK045) .......T........C.....T.........C.T..............T..C......T
10 20 30 40 50 60 70
.... | .... | .... | .... | .... | .... | .... | .... | .... | .... | .... | .... | .... | .... | .... | ... Dysmicoccus lavandulae GGTAAACGGACAGAGCCCGTGAATCCGGGCGACGGAATTCAAAATCGACGGCGTTCGCGCGTCGTCGGTTCGATATTTC Dysmicoccus multivorus (MBK047) .......C................T................................................A....T Trionymus aberrans (MBK084) .........................................G.................................... Trionymus Perrisi (MBK045) .........................................G....................................
90 100 110 120 130 140 150
.... | .... | .... | .... | .... | .... | .... | .... | .... | .... | .... | .... | .... | .... | .... | ... Dysmicoccus lavandulae GCCGCCATTTCACATTACGGACGTCGCGACCCGTTCGGTGTCGGTCCGCAGGAGACGCGCGAAAGTTCGTGGGCGCTCG Dysmicoccus multivorus (MBK047) ..........T-.T.............................................................T.. Trionymus aberrans (MBK084) ......GCCAA-TTCA......................C....................................TCGA Trionymus Perrisi (MBK045) ......GCCAA-TTCA......................C..................T.................TCG
170 180 190 200 210 220 230
.... | .... | .... | .... | .... | .... | .... | .... | .... | .... | .... | .... | .... | .... | .... | ... Dysmicoccus lavandulae CGGGTTTCGGCCTTGGCGGGCGTTCGCGAGTGCGCGTTT-TTTTTCTGGCCGACTCGCCGGACGGTAAGCGAAGCGGTG Dysmicoccus multivorus (MBK047) .......................................-...................................... Trionymus aberrans (MBK084) ...........-.C..TC.....................A...................................... Trionymus Perrisi (MBK045) ...........-.C..TC....................CA...........................G..........
250 260 270 280 290 300 310
.... | .... | .... | .... | .... | .... | .... | .... | .... | .... | .... | .... | .... | .... | Dysmicoccus lavandulae TCGCGGCGGCCTC--TCGCTAGCGCCGTCGCGTTACCGGTCTGCGAC-GAATCTTCGGGCCTCTTTCCGA Dysmicoccus multivorus (MBK047) .............--................................-...................... Trionymus aberrans (MBK084) G...........TCG..AGG...........................A...................... Trionymus Perrisi (MBK045) ............TCG.TA.G...........................-...................... We have not been able to discuss the material that Panis (1999) examined but the specimens that he identified as Dysmicoccus multivorus collected on lavandin are normally broadly oval. The species that we have described here is considered to be different as it is usually extremely elongate, possibly because it lives mainly in the flower buds.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |