
Axiokebuita Pocklington & Fournier, 1987
|
publication ID |
https://doi.org/ 10.5281/zenodo.171399 |
|
DOI |
https://doi.org/10.5281/zenodo.5623794 |
|
persistent identifier |
https://treatment.plazi.org/id/063DFA6E-6707-FF93-1100-FA15FC5F76AA |
|
treatment provided by |
Plazi |
|
scientific name |
Axiokebuita Pocklington & Fournier, 1987 |
| status |
|
Axiokebuita Pocklington & Fournier, 1987 View in CoL ( Figs. 1 View FIGURE 1 , 2 View FIGURE 2 )
Axiokebuita Pocklington & Fournier, 1987: 108 View in CoL .
Type species. Kebuita minuta Hartman, 1967 , by original designation (Pocklington & Fournier: 108).
Material examined. ANTARCTIC: holotype for Axiokebuita minuta ( USNM 55553, incl. 1 slide with parapodium), South Orkney Islands, 61°26’– 61°24’S, 41°55’W, 593–598 m, coll. USNS Eltanin, 13 Apr 1964; 1 spm ( USNM 56626), Scotia Sea, 62°25’S, 56°30’– 56°32’W, 300 m, coll. USNS Eltanin, 13 March 1964; 2 spms ( USNM 56625, incl. 2 slides with parapodia; Hartman’s original description recorded a single spm from this locality, but the vial actually includes 2 spms), Antarctic Peninsula, 64°54’S, 68°21’– 68°18’W, 412 m, coll. USNS Eltanin, 21 Oct 1962; 1 paratype for Axiokebuita millsi ( USNM 60574), Ross Sea, 73°59’S, 170°41’E – 73°58’E, 170°58’S, 608 m, coll. USNS Eltanin, 13 January 1968. NORWAY, TRONDHEIMSFJORD: 2 spms (used for DNA sequencing), Rødberg, 63°28.32’N, 10°00.02’E, 230–280 m, Lophelia reef, triangular dredge, 19 Jan 2002; 12 spms (2 preserved in formaldehyde, SMNH 75817; 8 used for SEM; 2 used for DNA sequencing), Rødberg, 63°28.36’N, 10° 00.04E, 180–250 m, Lophelia reef, triangular dredge, coll. FP, 21 Feb 2003; 10 specimens (6 preserved in formaldehyde, SMNH 75819; 2 used for SEM; 1 preserved in 95% ethanol, SMNH 75820; 2 used for DNA sequencing), same locality data, 26 Feb 2003.
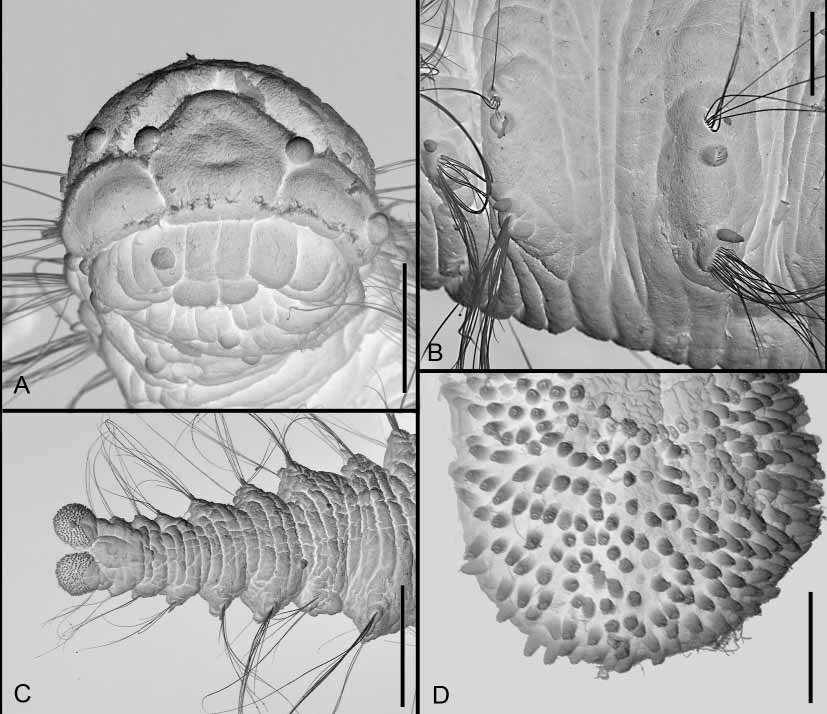
Description of specimens from Trondheim. Body elongate, slightly tapered at both ends ( Fig. 1 View FIGURE 1 ). Specimens 5–6 mm in length for 22–25 segments. Live specimens opaque, unpigmented, white to pinkish in color. Prostomium pentagonal, with straight frontal margin, bearing two distinct laterally directed and well delineated palps (see remarks below); eyes lacking ( Fig. 2 View FIGURE 2 A). Nuchal organs not visible on figures but may occur as ciliated bands bordering posterior part of prostomium. Fully everted proboscis not seen; visible basal part with longitudinal folds ( Fig. 2 View FIGURE 2 A). Peristomium complete ring, dorsally incised. Segment 1 similar to following segments. Each segment with four annullae. Notopodia and neuropodia joined, on a vertically oriented parapodial extension ( Fig. 2 View FIGURE 2 B). Notopodia with small knob ventral to chaetal bundle, neuropodia with digitate, pointed dorsal cirrus dorsal to chaetal bundle. Notochaetae less numerous than neurochaetae; both noto and neurochaetae long, fine capillaries. Intestine strongly spiraling in middle part of body. Pygidium with two padshaped lobes, densely covered by digitate papillae ( Fig. 2 View FIGURE 2 C, D); each papilla terminating in 4–5 pores.
Distribution of Axiokebuita . South Orkney Islands, Antarctic Peninsula, Scotia Sea, Ross Sea, east Canada, western Norway.
Remarks. As seen from studies of live specimens, the papillae covering the pygidial lobes ( Fig. 2 View FIGURE 2 C, D) are used to adhere to surfaces. Although they have not been noted previously in the literature, examination of types and other specimens shows them to be present in both A. minuta and A. millsi . Unless they have been overlooked in other, closely related scalibregmatids, they constitute an apomorphy for Axiokebuita .
We are unable to state whether the Norwegian specimens should be referred to A. minuta , to A. millsi , or to an undescribed species. Axiokebuita minuta , according to Pocklington & Fournier (1987), should lack notopodial postchaetal lamellae, (here referred to as notopodial knobs due to their shape and position, which is below, rather than behind the chaetae), it should have neuropodial lamellae (here referred to as neuropodial cirri due to their cirriform shape) in posterior segments only, and it should have a simple pygidial ring. However, reexamination of Hartman’s type did not confirm these differences; the neuropodial cirri are not restricted to posterior segments but occur minimally also on anterior segments (median segments are too damaged to allow observation), and the pygidium is bilobed and actually have the same papillation as shown in Fig. 2 View FIGURE 2 C and D, not previously observed in the literature. As for the notopodial lobes these may, or may not, be presentthe type is too poorly preserved to permit unequivocal assessment of this feature. This situation is further complicated by the fact that the description of A. millsi was based on material both from east Canada and from the Antarctic. Without access to further specimens in good condition from the type locality of A. minuta , it is difficult to state if the separation between the two can be corroborated by other features. For this reason, we do not allocate our Norwegian specimens further than to Axiokebuita .

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Family |
Axiokebuita Pocklington & Fournier, 1987
| Persson, Jenny & Pleijel, Fredrik 2005 |
Axiokebuita
| Pocklington 1987: 108 |