
Rhopalomyia capitata Felt 1908
|
publication ID |
https://doi.org/10.5281/zenodo.188745 |
|
DOI |
https://doi.org/10.5281/zenodo.6217047 |
|
persistent identifier |
https://treatment.plazi.org/id/074287C9-FFC8-E31B-FF01-FBBC5B313F2C |
|
treatment provided by |
Plazi |
|
scientific name |
Rhopalomyia capitata Felt 1908 |
| status |
|
Rhopalomyia capitata Felt 1908 View in CoL
Rhopalomyia capitata Felt 1908: 363 View in CoL
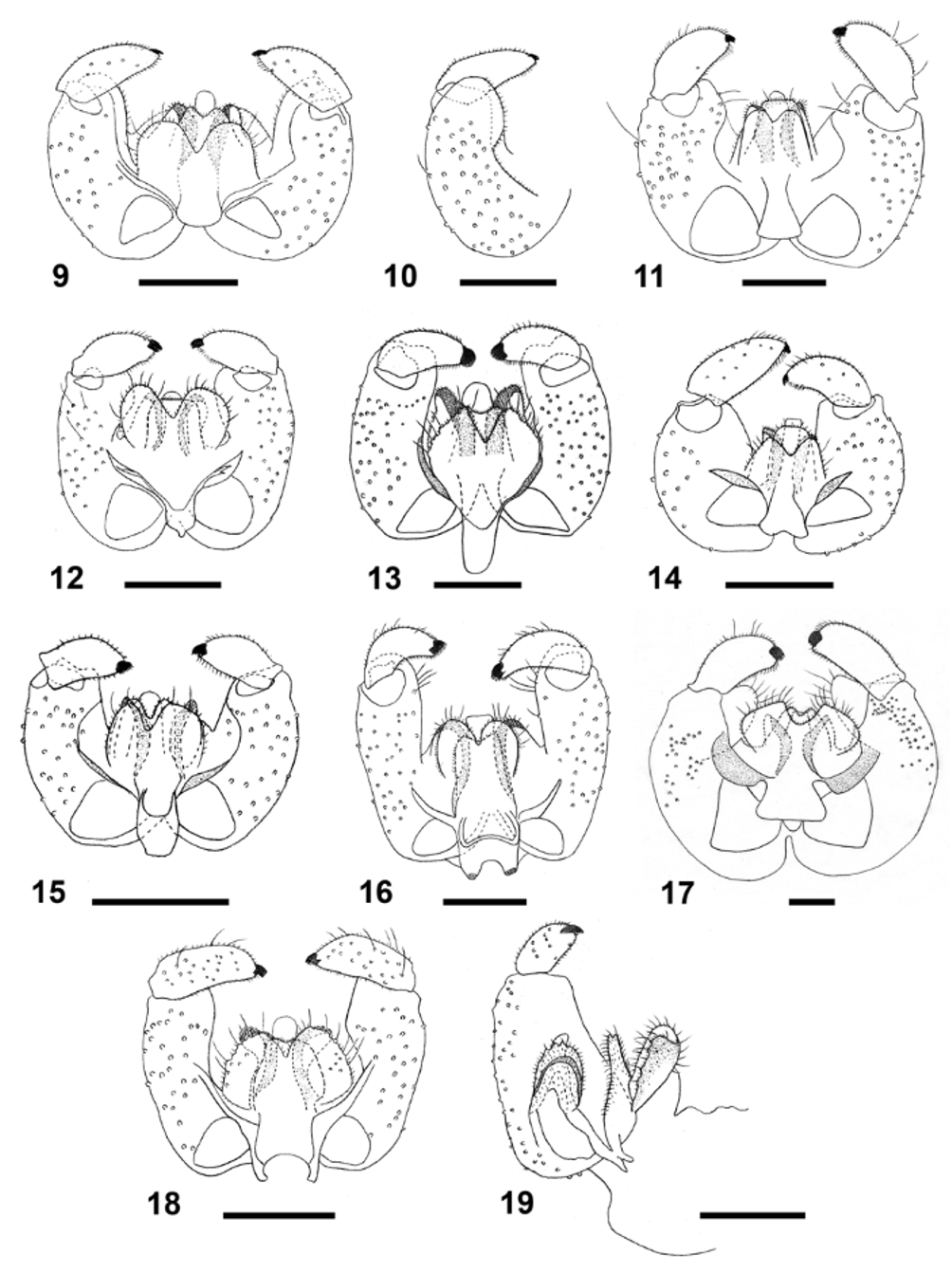
Adult: Antenna with 18 flagellomeres in male, 17–18 or 18–19 flagellomeres in female (spring and summer generations, respectively); necks of male flagellomeres III–VII 0.78–0.89 times as long as nodes, female flagellomeres without necks. Palpus 2-segmented, second segment at least 1.5 times longer than first, setose and setulose. Wing length 2.9–3.5 mm in male, 3.2–3.4 mm in female. Legs densely covered by brownish scales; empodia considerably longer than claws. Male terminalia ( Fig. 13 View FIGURES 9 – 19 ): gonocoxite cylindrical, setose and setulose, with prominent, setose mediobasal lobe as long as aedeagus; gonocoxal apodeme narrow, undivided; gonostylus evenly curved in posterior margin, straight in anterior margin, setose and setulose throughout, with relatively large, brush-like tooth; aedeagus very wide, truncate; hypoproct M-shaped, with shallow notch, setulose; cerci deeply separated by a triangular notch, strongly setose and setulose. Female abdomen (Fig. 29): tergite 7 trapezoid, less sclerotized along lateral and anterior margins, with two anterior trichoid sensilla, several rows of setae posteriorly, and groups of setae mesolaterally; tergite 8 wide Y-shaped, with two anterior trichoid sensilla; ovipositor 5.0–5.6 times as long as tergite 7.
Pupa ( Figs. 42–43 View FIGURES 40 – 47 ): Light orange. Antennal bases tapering into tiny pointed tips, their bases wide V-shaped in frontal view. Frons with tiny pointed projection at mid width, which is absent in pupae of spring generation, and with tapered posterolateral projections; posterior edge widely rounded. Prothoracic spiracle divided apically into two lobes. Abdominal segments with tiny barbs throughout.
Type material: Rhopalomyia capitata Felt. Syntypes: 1 female, 1 male, USA, W. Nyack, NY, 23/IX/1907, E.P. Felt, ex. S. gigantea (as S. serotina ), Felt # a1750, deposited in Felt Collection.
Other material examined (all from S. gigantea ): 2 males, 2 females, USA, NY, West Dryden, 15/IX/1987, M.V. McEvoy; 3 exuviae, USA, PA, Bucknell University Chillisquaque Creek Natural Area, 15/V/2005, N. Dorchin; 3 males, 3 females, USA, PA, Lewisburg, Furnace Rd., 22/V/2005, N. Dorchin; 7 pupae, USA, PA, Bucknell University Chillisquaque Creek Natural Area, 9/IX/2006, C. Blair; 1 male, USA, PA, Bucknell University Chillisquaque Creek Natural Area, 12/IX/2006, N. Dorchin; 1 pupa, USA, PA, Lewisburg, Furnace Rd., 23/V/2007, N. Dorchin; 2 pupae, USA, PA, Montour Environmental Preserve, 25/V/ 2007, M. Wise.
Host: Solidago gigantea and occasionally S. canadensis and S. leavenworthii . Felt (1915) mentioned S. serotina and S. canadensis as the host plants of R. capitata . Solidago canadensis was the name used by Felt for S. altissima and is an erroneous record for R. capitata , whereas S. serotina was synonymized under S. gigantea ( Semple and Cook 2006) .
Gall and biology: The species is bivoltine and galls of the two generations are overall similar in structure. The spring-generation galls, described here for the first time, appear in early to mid May in young S. gigantea sprouts, and are therefore found very close to the ground ( Fig. 74 View FIGURES 70 – 77 ). The larvae that induce these galls hatch in the fall of the previous year and overwinter as first instars in rhizomes without causing any deformation. This was discovered when several rhizome sections that were dug, cut and planted in our research greenhouse in very early spring developed galled sprouts without having been exposed to any adults. The galls are composed of many short and narrow leaves among which 1–8 white, conical chambers are situated ( Fig. 77 View FIGURES 70 – 77 ), each containing a single orange larva. The white chambers appear in the gall only when larvae are second instars. The central complex of short leaves is about 3 cm in diameter and surrounded by 5–10 much longer and wider leaves, giving the gall a star-like shape and an overall diameter of up to 10 cm ( Fig. 74 View FIGURES 70 – 77 ). At young stages of development, the longer, surrounding leaves may sheath tightly the center of the gall and these loosen at a later stage. The spring generation galls of R. capitata can easily be mistaken for Dasineura folliculi galls that develop on the same host at the same time ( Dorchin et al. 2007). However, D. folliculi galls never contain multiple short leaves at their center, and instead are composed of several similar-sized leaves that are wider and thicker at their base, and are accompanied by yellowish spots on leaves below and around the gall.
Pupae developing in the spring generation galls emerge from mid to late May and induce the much slower-developing summer-generation galls ( Fig. 75 View FIGURES 70 – 77 ), which become apparent around mid June and vary considerably in size. These galls are composed of several wide leaves that surround dozens of densely packed, smaller and shorter leaves, giving the gall a shape of a little sunflower that appears flatter than the similar gall of R. solidaginis on S. altissima (see Fig. 71 View FIGURES 70 – 77 ). Occasionally, the outer leaves merge to form a continuous sheath around the central part of the gall. Summer galls reach their final size when the larvae are still tiny first instars that are found at the base of the rosette leaves. Six to twenty white, conical chambers, similar to those found in the spring galls, appear in the gall only when the larvae molt into third instars. Larvae are usually found deep at the bottom of the chamber, facing down. The chambers are 4–6 mm high and 1–2 mm wide, and are situated among the short leaves in the center of the gall rather than being individually surrounded by a group of longer leaves, as in the rosette gall of R. solidaginis . Pupation takes place in late August to mid September and adults emerge in September and early October.
During summer, some shoot tips can support both R. capitata and D. folliculi larvae at the same time, and the resulting gall shows morphological attributes of both species, namely, feeding spots on outer leaves ( Dasineura ) and a group of many shortened leaves at its center ( Rhopalomyia ). Larvae of R. capitata are heavily attacked by polyembryonic endoparasitoids, and parasitism levels can reach 90% in certain localities and dates. Ectoparasitoids are also found in the galls and feed both on larvae and on pupae of the gall inducer.
Remarks: Rhopalomyia capitata and its close relative R. solidaginis are two of several species that have been described from similar rosette galls on goldenrods, a fact that caused confusion with regard to the identity and validity of these species ( Felt 1915, Gagné 1989). Although their summer-generation galls are superficially similar, R. capitata and R. solidaginis are consistently found on different hosts (the former on S. gigantea , the latter on S. altissima ), their adults show clear morphological differences, and recent molecular analyses confirm that they constitute distinct species ( Stireman et al. 2005, Dorchin et al., in prep.). Fontes et al. (1994) recorded the galls of R. capitata from S. leavenworthii and S. gigantea , but attributed them erroneously to S. solidaginis . Stireman et al. (2005) later showed that specimens recovered from galls on S. leavenworthii nested within the R. capitata clade. Both R. capitata and R. solidaginis are relatively large but R. solidaginis is usually larger and adults of its summer generation have more numerous antennal flagellomeres. Females of both species have short ovipositors relative to other Rhopalomyia species from goldenrods, but can be distinguished from each other by the typical shape of the 8th tergite in R. solidaginis , whose arms are narrow at their bases and widen anteriorly (Fig. 38). The male gonocoxite in R. capitata is much more slender and less robust than the unmistakably inflated gonocoxite of R. solidaginis ; the gonocoxal apodeme is longer and more slender in R. capitata , and its gonostylus is not as stout.
Lastly, despite the superficial similarity of their summer galls, the spring generation galls of R. capitata and R. solidaginis are strikingly different morphologically: those of R. solidaginis contain 1–2 chambers in barely modified shoot tips ( Figs. 70, 72 View FIGURES 70 – 77 ), whereas those of R. capitata form complex multi-chambered rosettes ( Fig. 74 View FIGURES 70 – 77 ).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Rhopalomyia capitata Felt 1908
| Dorchin, Netta, Mcevoy, Miles V., Dowling, Todd A., Abrahamson, Warren G. & Moore, Joseph G. 2009 |
Rhopalomyia capitata
| Felt 1908: 363 |