
Orobothriurus huascaran, Ochoa & Ojanguren Affilastro & Mattoni & Prendini, 2011
|
publication ID |
https://doi.org/ 10.1206/359.1 |
|
publication LSID |
lsid:zoobank.org:pub:870DFDFC-45BF-49CF-8C3F-2527DB611CFC |
|
DOI |
https://doi.org/10.5281/zenodo.4618600 |
|
persistent identifier |
https://treatment.plazi.org/id/6905D1CB-ECAA-4A2D-B3C1-AF3EA985BCDA |
|
taxon LSID |
lsid:zoobank.org:act:6905D1CB-ECAA-4A2D-B3C1-AF3EA985BCDA |
|
treatment provided by |
Felipe |
|
scientific name |
Orobothriurus huascaran |
| status |
sp. nov. |
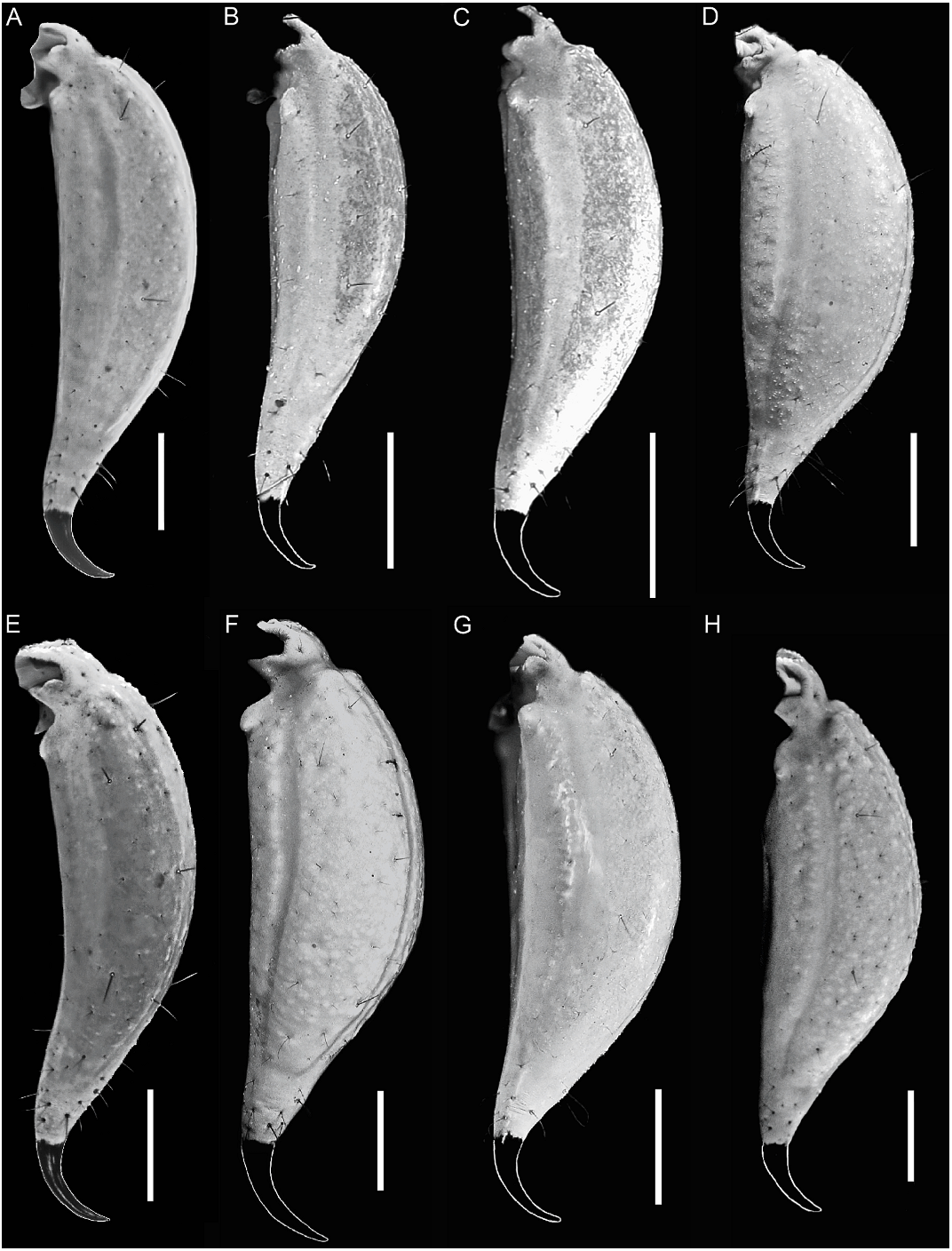
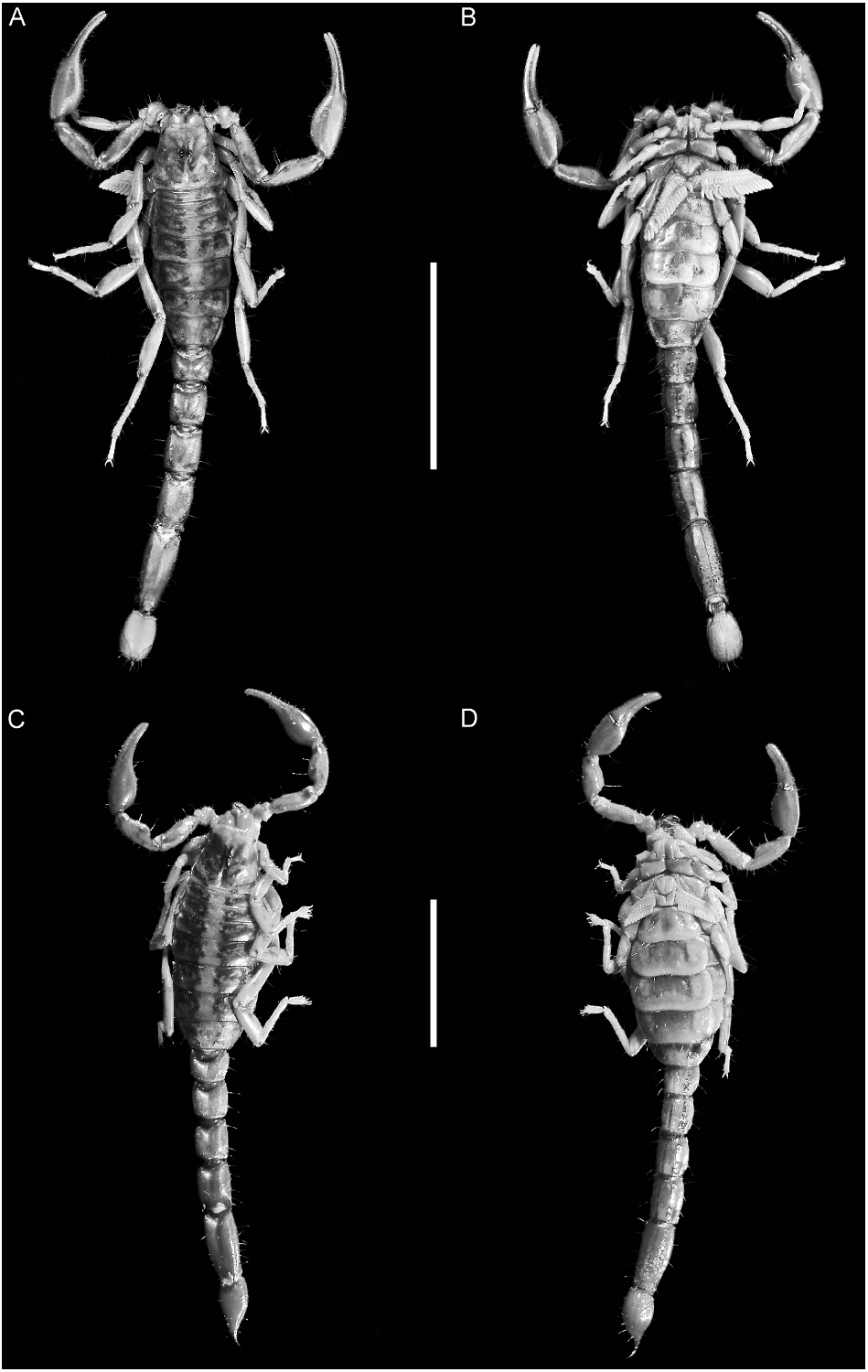
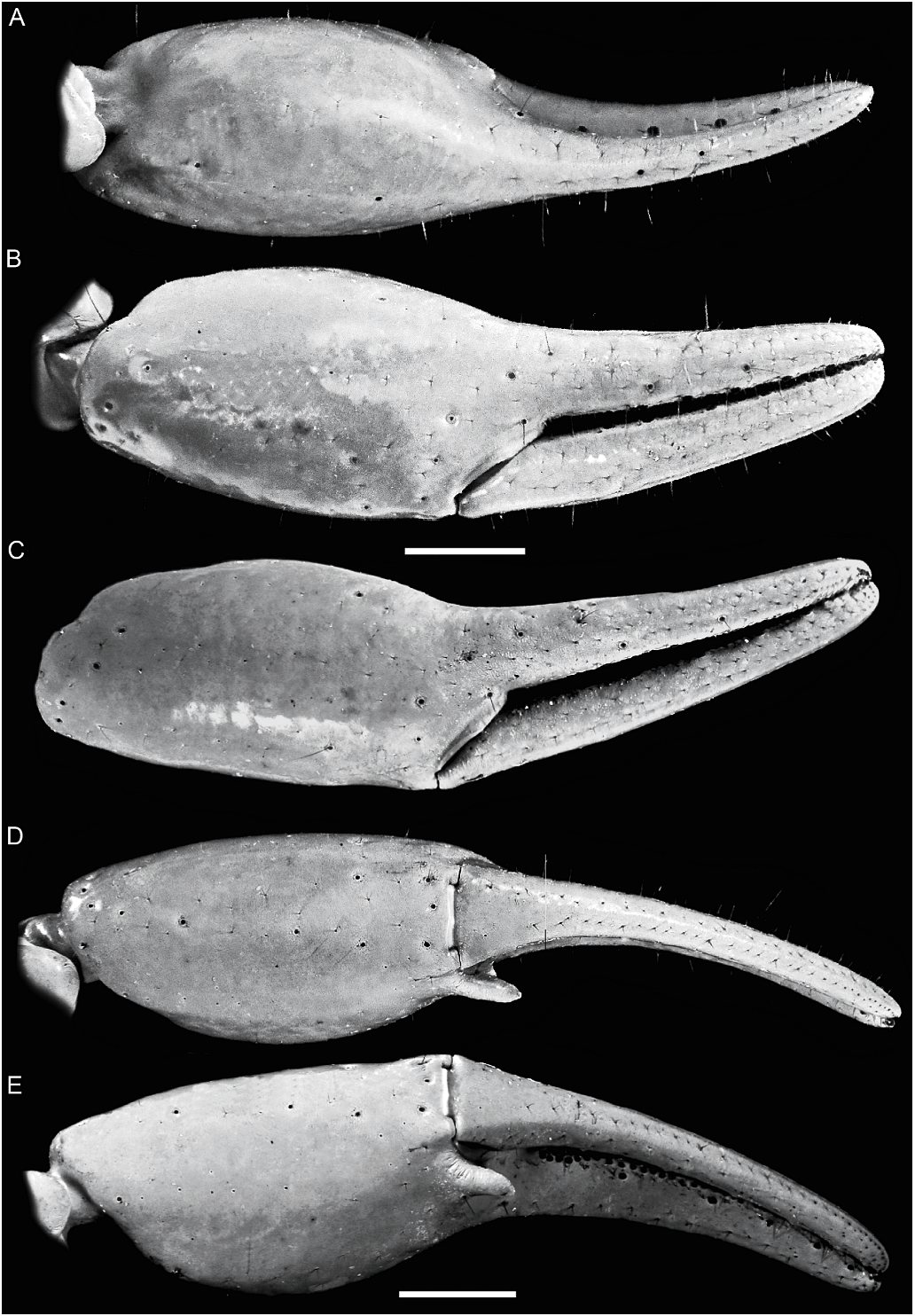
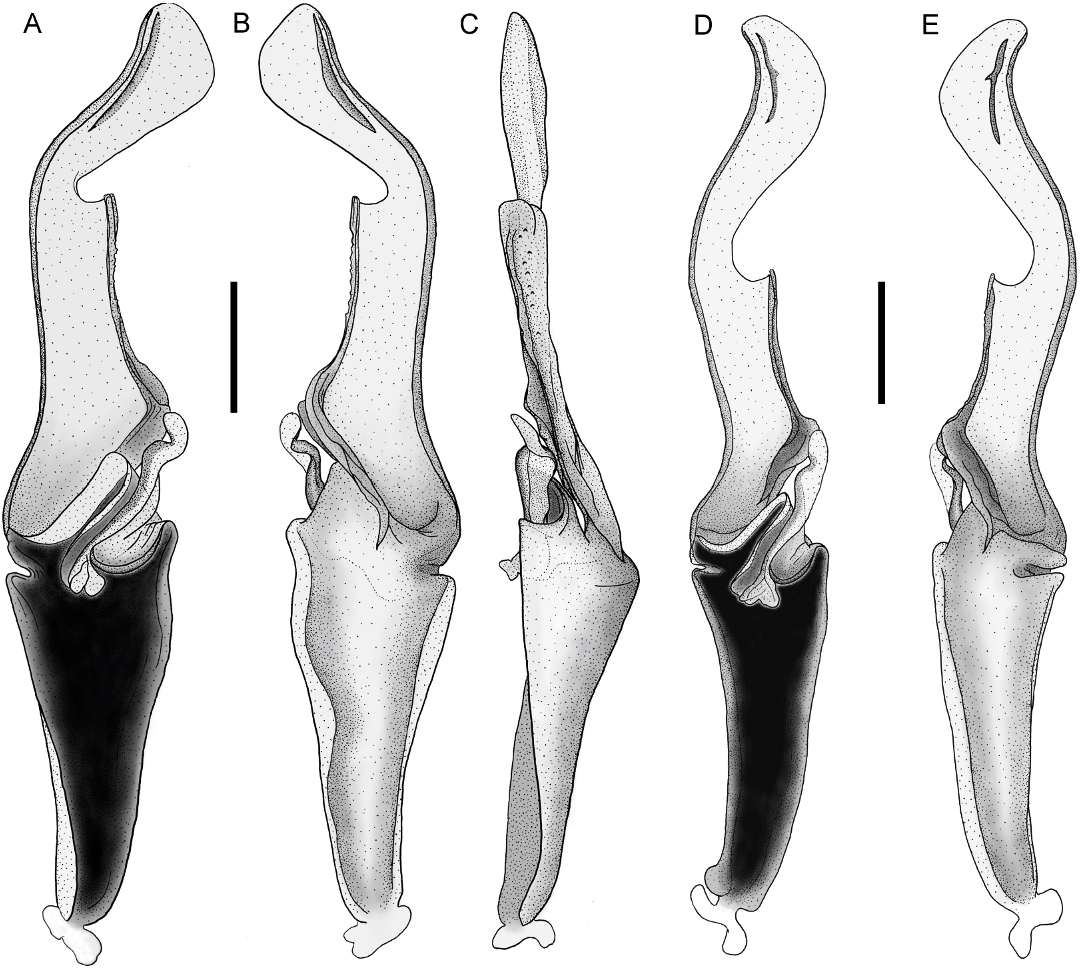
Orobothriurus huascaran View in CoL , n. sp. Figures 4D View Fig , 11C View Fig , 12A View Fig , 14D View Fig , 19D View Fig , 21F View Fig , 22D View Fig , 23D View Fig , 24C View Fig , 37–39 View Fig View Fig View Fig , 40 View Fig A–C, 52; table 3
Orobothriurus crassimanus: Polis, 1990: 252 ; Lourenço, 1997: 588 (part); Coddington and Colwell, 2001: 207; Lourenço, 2002: 400; 2003: 227; Lourenço and Qi, 2006: 290. [misidentification].
Orobothriurus paessleri: Lourenço and Dastych, 2001: 54 View in CoL (part) [misidentification: 1 ♀, from Quebrada Yanganuco].
Orobothriurus sp. 2 : Ochoa, 2004a: 50, 73, figs. 1, 2, 14, 21, table 1. View TABLE 1
TYPE MATERIAL: PERU: Ancash Department: Huaraz Province: Holotype ♂ ( MHNC), Cordillera Blanca, Huascarán National Park ,
Ishinca , 09 ° 229490S 77 ° 279390W, 4100 m, 19.vi.1998, J.A. Ochoa. Paratypes: Huaraz Province : Cordillera Blanca , Huascarán National Park, Ishinca, from 09 ° 229490S 77 ° 279 390W, 4050 m, to 09 ° 229280S 77 ° 249360W, 4910 m, 18–19.vi.1998, J.A. Ochoa, 3 ♂, 3 ♀, 2 juv. ♂ ( MHNC) ; Quebrada Querococha , Catac [09 ° 329S 77 ° 249W], 3900 m, vi.1981, W.R. Lourenço, 1 ♂, 2 ♀ ( MHNG) ; Querococha , 3850 m, 16.iii.1994, M. Etonti, 1 ♂ ( MRSN) ; Pastoruri [09 ° 519S 77 ° 129W], 4900 m, 18.iii.1994, M. Etonti, 1 ♂ ( MRSN). Huaylas Province : Laguna Parón [08 ° 599430S 77 ° 409410W, 4160 m], 16.v.2010, R. Pinto da Rocha and D. Silva, 2 ♂ ( MZSP), 1 ♂, 1 ♀ ( MUSM), 2 juv. ( AMNH [ LP 10709 , 10713 ]). Yungay Province: Llanganuco, 4000 m, 2.i.1976, O. F. Francke, 1 ♂, 5 ♀ (MACN- Ar 17889) ; same locality, 3600–3850 m, 15.iii.1994, M. Etonti, 2 ♀ ( MRSN) ; same locality, 3810 m, vi.1997, F. Pribik, 1 ♀, 1 juv. ( FKPC) ; Llanganuco , Huascarán National Park, 09 ° 049420S 77 ° 389570W, 3800–4000 m, 28–29.ix.1998, J.A. Ochoa, Polylepis forest, 1 ♂, 3 ♀ paratypes ( MHNC), 1 ♂, 1 ♀ ( AMNH), 1 ♂, 1 ♀ ( LBRE) ; Chinancocha , Llanganuco, 09 ° 049180S 77 ° 389380W, 3844 m, 15.v.2010, R. Pinto da Rocha and D. Silva, 1 ♀ ( MZSP) .
ADDITIONAL MATERIAL: PERU: Ancash Department: Carhuaz Province: Huascarán National Park , Aquilpo , Porganillo , 09 ° 199 020S 77 ° 279320W, 3730 m, 15.vi.1998, J.A. Ochoa, 1 ♂, 3 ♀, 2 juv. ♂, 1 juv. ♀ ( MHNC). Huaraz Province : Cordillera Blanca, Huascarán National Park , Ishinca ravine, 09 ° 229490S 77 ° 279390W, 4100 m, 19.vi.1998, J.A. Ochoa, 1 ♂, 3 juv. ( MHNC). Huaylas Province : Laguna Parón [08 ° 599580S 77 ° 419090W, 4180 m], ii.1981, W.R. Lourenço, 1 juv. ♀ ( MHNG). Recuay Province : Huama, 4200 m, 22.viii.1972, P. Brignoli, 6 ♀ [mislabeled ‘‘2 ♂, 4 ♀ ’’], 14 first instar juv. ( MHNG). Yungay Province : Llanganuco, 4300 m, ii.1981, W.R. Lourenço, 3 ♀ ( MHNG), 4400–5500 m [see discussion on the altitude record in Scorpiones ], ii.1981, W.R. Lourenço, 2 ♀ ( MHNG) ; Llanganuco, Huascarán National Park , 09 ° 049420S 77 ° 389570W, 3800–4000 m, 28–29.ix.1998, J.A. Ochoa, Polylepis forest, 1 ♂, 2 ♀ ( MHNC) .
ETYMOLOGY: The specific name is a noun in apposition derived from the Huascarán National Park, Ancash Department, central Peru, where the type locality of this species is situated. This area belongs to the Cordillera Blanca, the world’s highest tropical mountain range.
DIAGNOSIS: Orobothriurus huascaran may be distinguished from other species of the genus by the following combination of characters. The distal border of the apex of the hemispermatophore is slightly rounded, the internal lateral projection of the frontal crest vestigial, and the external lateral projection well developed in O. huascaran (fig. 40A–C) compared to other species in which the distal border of the apex is subtriangular, and both lateral projections of the frontal crest are well developed. Pedipalp chela trichobothrium Db is situated slightly closer to Dt than to Eb 3 in O.
huascaran (fig., 39C), but equidistant between Dt and Eb 3 in other species. The VM and VL carinae are restricted to the posterior two-thirds of metasomal segment V in O. huascaran (fig. 21F), absent in O. wawita and complete in all other species. Orobothriurus huascaran is similar to four other Peruvian species, O. atiquipa , O. curvidigitus , O. paessleri , and O. quewerukana , in several respects. Pedipalp chela trichobothrium Et 3 is situated proximal to Est, and Esb situated between Eb 2 and Eb 3 in all these species, except O atiquita , in which Esb is situated dorsal to Eb 2. The hemispermatophore of these species exhibits an elongated frontal crest, with a strongly curved, S-shaped apex and a basal lobe with a well-developed terminal process. Orobothriurus huascaran may be distinguished by the presence of VM and VSM carinae on sternite VII and metasomal segment I (fig. 19D), which are absent in O. atiquipa , O. curvidigitus , O. paessleri , and O. quewerukana (figs. 18A, 19A, C). Additionally, the pedipalp chela movable finger of the male is straight in O. huascaran (fig. 39C) and curved in O. curvidigitus , O. paessleri , and O. quewerukana (fig. 26B, C, 43C).
DESCRIPTION: Based on holotype ♂ and paratypes. Measurements of holotype ♂ and paratype ♀ recorded in table 3.
Total length: ♂, 26.43–30.79 mm (n 5 9, mean 5 27.97 mm); ♀, 30.05–33.21 mm (n 5 10, mean 5 31.65 mm).
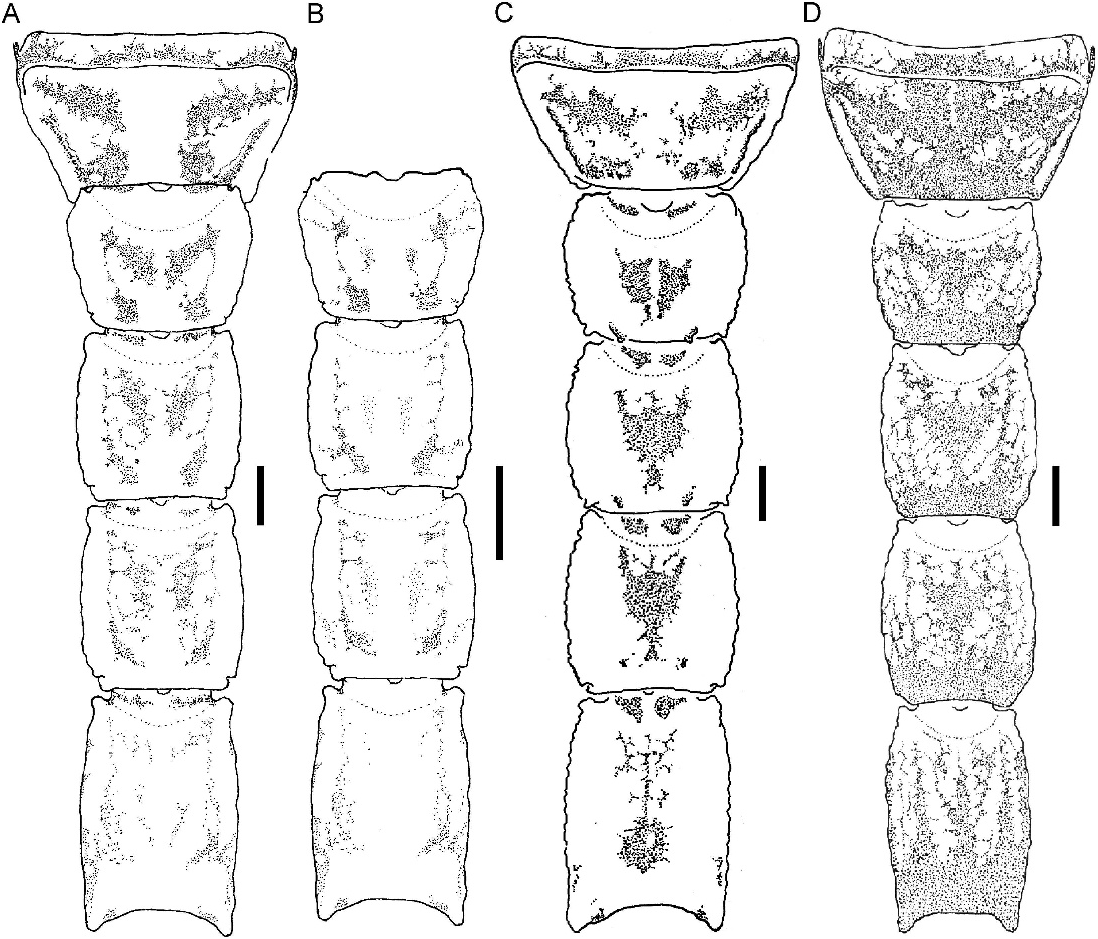
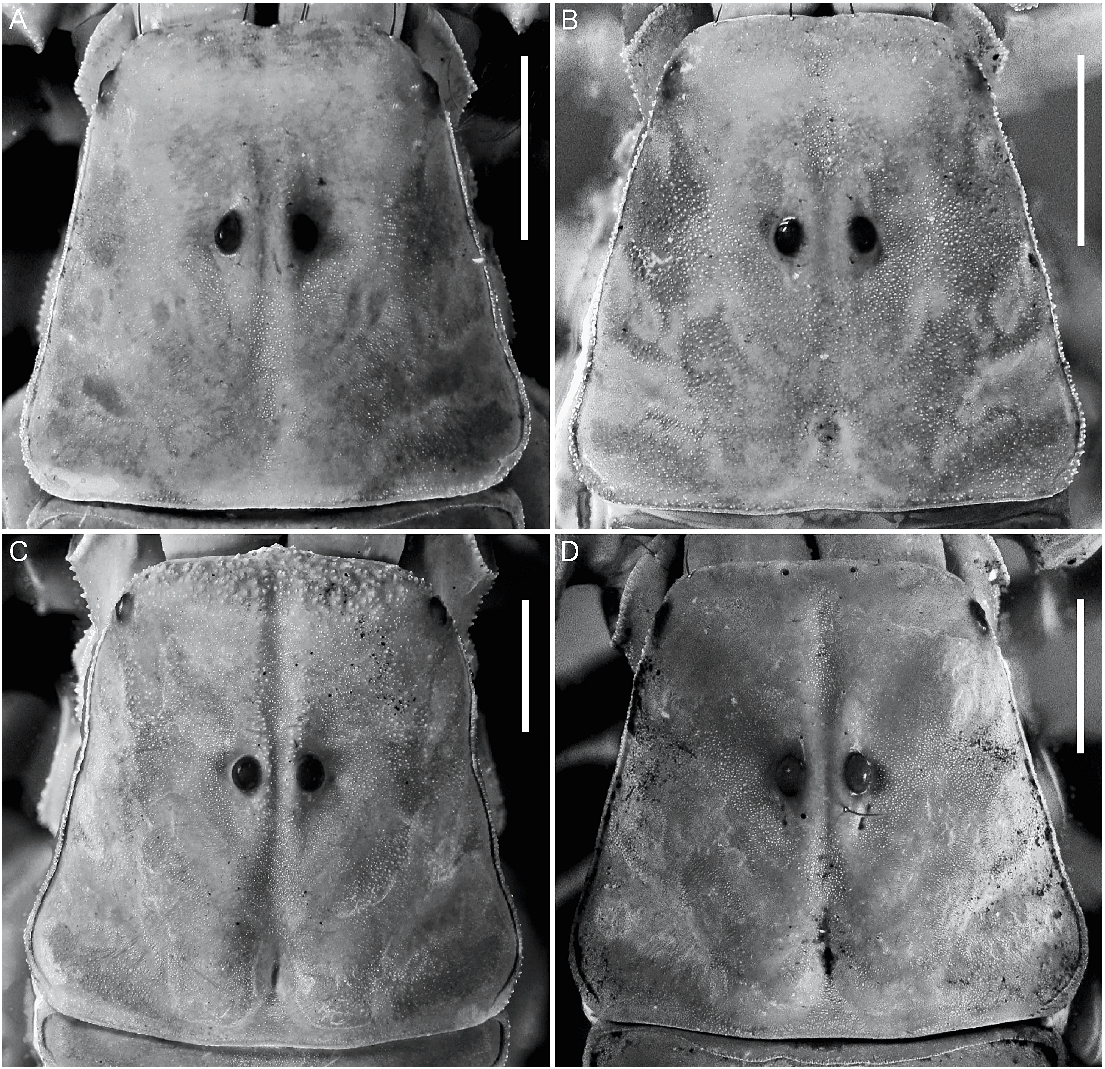
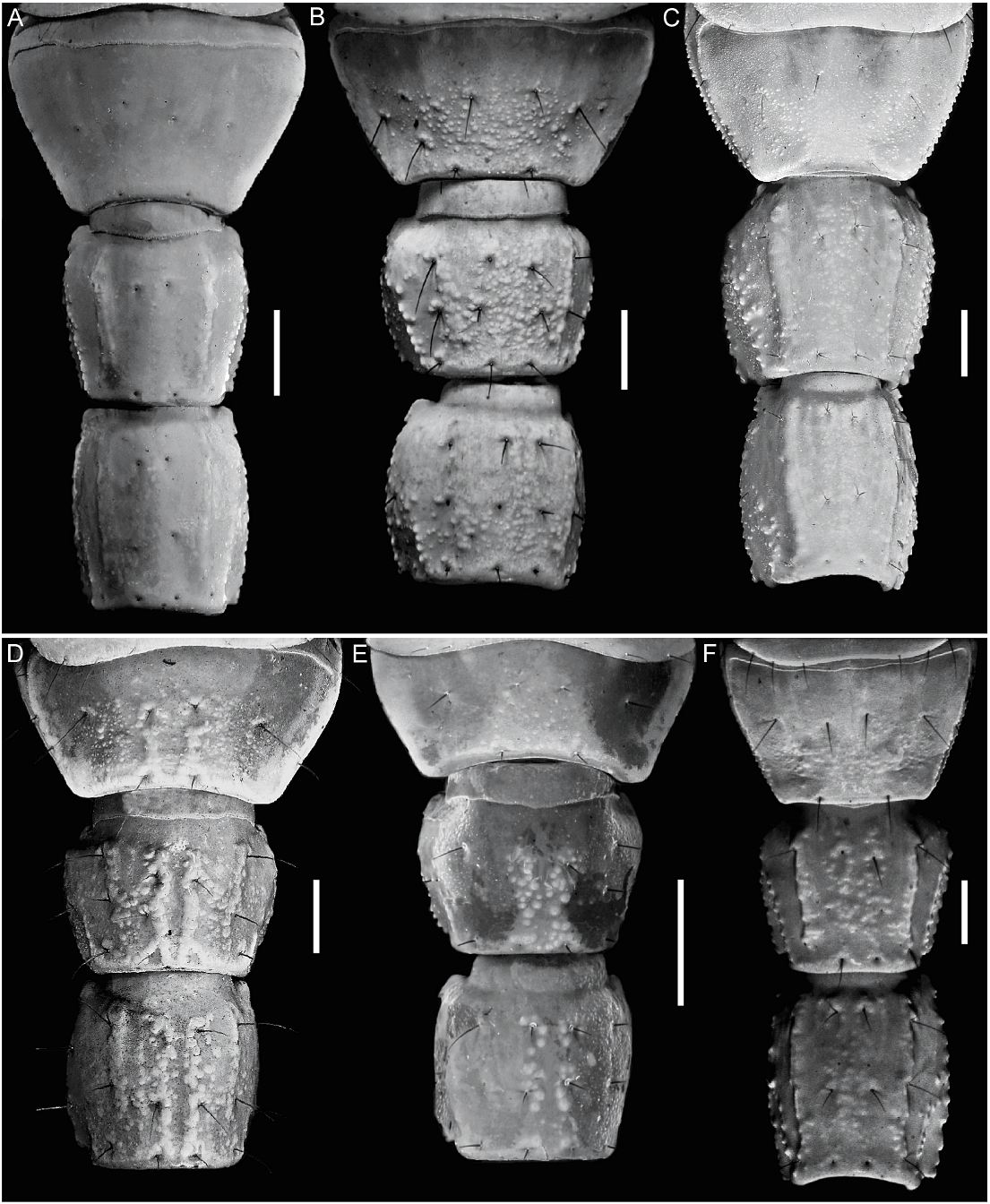
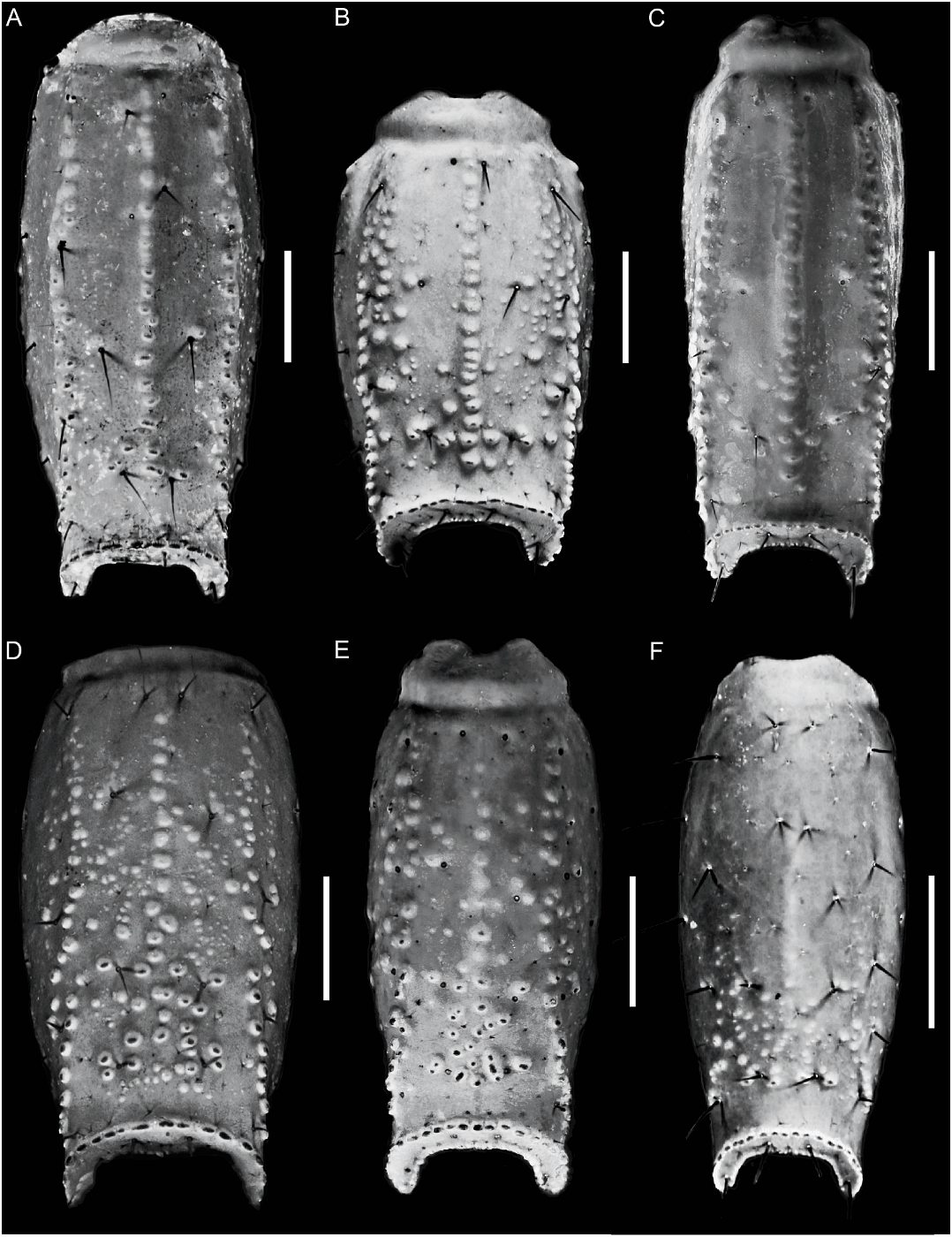
Color: General color yellowish with dark brown spots (fig. 4D). Carapace densely pigmented, especially laterally and posteriorly; anterior margin with narrow stripe (fig. 37A, C); median lateral surfaces densely pigmented; posterolateral surfaces with dense reticulate pigmentation; median ocular tubercle and lateral ocelli dark brown to black; posterior half of anteromedian longitudinal sulcus, median ocular tubercle and anterior half of posteromedian longitudinal sulcus densely pigmented; posterolateral sulci unpigmented. Chelicerae, dorsal surfaces with fine reticulate pigmentation, contiguous distally near base of movable finger; movable finger external surface pigmented. Pedipalp coxa and trochanter faintly pigmented; femur densely pigmented along margins with small unpigmented areas in proximal half; patella densely and irregularly pigmented; chela manus with faint longitudinal stripes along carinae, contiguous at base of movable finger. Legs, femur, and patella densely pigmented prolaterally; tibia and basitarsus faintly pigmented. Tergites I–VI each with two dark spots sublaterally along entire margin and pretergites, delimiting broad, unpigmented median stripe; VII with similar pattern and additional reticulate pigmentation (fig. 12A). Sternum, genital opercula and pectines unpigmented. Sternites III–VI unpigmented, VII with two dark stripes sublaterally (fig. 11C). Metasomal segments I–III, dorsal surfaces each with two subtriangular spots medially, separated by narrow unpigmented line on I, contiguous or separated on II, contiguous on III, becoming broader posteriorly and often connected to median pigmentation by fine reticulate pigmentation, with additional pigmentation along DL carinae; lateral surfaces densely pigmented, especially in anterior half, posterior half with reticulate pigmentation (figs. 11C, 12A); ventral surface with narrow VM stripe, not contiguous with lateral pigmentation, and with two dark VL stripes, becoming broader in posterior half and extending to LIM carinae. Segment IV as for segment III, but with dorsal surface more densely reticulate. Segment V, dorsal surface faintly pigmented along DL carinae; lateral surfaces with dense reticulate pigmentation; ventral surface with three dark stripes, two VL and one VM, becoming slightly broader in posterior half but not contiguous with lateral pigmentation (fig. 11C). Telson vesicle with narrow VM and two broad VL stripes, separated by two narrow unpigmented stripes; aculeus sclerotized, dark reddish-brown.
Chelicerae: Movable finger with two subdistal teeth.
Carapace: Surfaces smooth in anterior third, finely granular elsewhere. Anterior margin linear, without median notch (fig. 14D). Anteromedian longitudinal sulcus complete, well developed, more so in ♂; median ocular and posteromedian longitudinal sulci well developed; posterolateral sulci obsolete. Median ocular tubercle raised, situated anteromedially; median ocelli two ocular diameters apart.
Pedipalps: Femur, DI and VI carinae complete, finely granular; DE carina obsolete, smooth (fig. 38A); internal surface finely and sparsely granular medially; other intercarinal surfaces smooth. Patella, DI and VI carinae obsolete, finely granular (♂) or smooth (♀) (fig. 38B–D); internal surface with two prominent granules distally, the larger adjacent to trichobothrium i; other intercarinal surfaces smooth. Chela manus slender, fingers relatively elongated (fig. 39); length/width ratio: ♂, 3.46–4.14 (n 5 11, mean 5 3.76), ♀, 3.37–4.14 (n 5 20, mean 5 3.74); D, DS, DMA, DI, and VM carinae obsolete; intercarinal surfaces smooth; internal surface with acuminate apophysis (♂) or low bulge (♀) near articulation of movable finger (fig. 39A, D); fingers, dentate margins each with median denticle row and 4–5 pairs of internal and external accessory denticles.
Trichobothria: Femur with 3 trichobothria, patella with 19, chela with 27 (figs. 38, 39). Chela trichobothrium Et 3 situated proximal to Est; Esb situated between Eb 2 and Eb 3 (fig. 39).
Tergites: Tergites I–VI, surfaces finely granular. Tergite VII tetracarinate, paired DL carinae restricted to posterior two-thirds of segment, paired DSM carinae to posterior half; surfaces more coarsely and densely granular in posterior half.
Legs: Femur and patella, prolateral surfaces finely granular, retrolateral surfaces smooth. Femur, ventral carinae weakly developed; other carinae absent. Patella acarinate. Telotarsi, pro- and retroventral rows of spiniform macrosetae with following counts on leg I, 1/1; II, 2/2; III and IV, 3/3.
Pectines: Pectinal tooth count: ♂, 17–20 (n 5 30, mode 5 18); ♀, 14–17 (n 5 56, mode 5 15).
Sternites: Sternites III–VI, surfaces smooth; spiracles small, narrow. Sternite VII, surface smooth in anterior half, granular in posterior half; VL and VSM carinae well developed (♀, fig. 19D) or obsolete (indistinct from granulation) to absent (♂).
Metasoma: Segment I, DL carinae complete, more developed in ♀ ; usually one pair of DL macrosetae; ML carinae complete; two pairs of ML setae; LIM carinae obsolete, entirely smooth or with few granules in posterior third; VL and VSM carinae complete, obsolete, smooth (♂) or granular (♀) ; three pairs of VL and two or three pairs of VSM macrosetae (fig. 19D). Segments II and III, DL carinae complete; one pair of DL macrosetae; ML carinae restricted to posterior half (♂) or complete but less developed than on segment I (♀) ; two pairs of ML macrosetae; LIM carinae reduced to one or two granules posteriorly (♀) or absent (♂) on segment II, absent on III ; VL carinae as for segment I, but less developed (♀) or absent (♂) on II, obsolete (♀) or absent (♂) on III ; VSM carinae as for segment I, but less developed on II, obsolete (♀) or absent (♂) on III ; three pairs of VL and VSM macrosetae. Segment IV, DL carinae complete; one or (usually) two pairs of DL macrosetae; ML carinae reduced to few small granules posteriorly; LIM carinae absent; VL and VSM carinae absent; three pairs of VL and four pairs of VSM macrosetae. Segment V, length/ width ratio: ♂, 2.07–2.40 (n 5 5, mean 5 2.17), ♀, 1.71–2.02 (n 5 7, mean 5 1.87) ; DL carinae reduced to three or four prominent granules anteriorly; lateral intercarinal surfaces smooth (♂) or finely and sparsely granular medially near VL carinae (♀) ; VL carinae usually restricted to posterior twothirds (but complete, weakly granular in anterior third in some ♀), with terminal granules more developed ; VSM and VM carinae usually obscured by surface granulation in posterior two-thirds (fig. 21F), but VM carinae faintly evident in some ♀ (fig. 21D) ; four pairs of VL and VSM macrosetae; two pairs of macrosetae along posterior margin; ventral intercarinal surfaces densely granular in posterior two-thirds, more so in ♀, granules more acute in ♂.
Telson: Length/height ratio: ♂, 2.86–3.52 (n 5 11, mean 5 3.24); ♀, 2.61–3.39 (n 5 19, mean 5 3.04). Vesicle slightly elongated (♂, fig. 23D) or globose (♀, fig. 24C); dorsal surface smooth, flat, gland not apparent (♂); ventral surface smooth (♂) or granular in anterior third (♀). Aculeus short and curved.
Hemispermatophore: Apex shorter than frontal crest, distal border slightly rounded;
distal crest curved like ventral margin. Frontal crest elongated; basal part oblique; distal part parallel to ventral margin of lamina, lateral projections slightly undulated and larger than basal part, ental lateral projection vestigial, ectal lateral projection complete. Basal lobe, terminal process extending to constriction of frontal crest (fig. 40A–C).
DISTRIBUTION: Orobothriurus huascaran is endemic to the Ancash Department of central Peru (fig. 52). All known localities of this species occur in the Cordillera Blanca mountain range of the western Andes, the world’s highest tropical mountain range (fig. 3F). This species has been collected from 3730 m (Porganillo, trekking route to Aquilpo mountain) to 4910 m (Ishinca ravine). It is present above 4050 m in the Ishinca area, but more abundant at 4100 m; only four specimens were found above 4500 m. The record of this species from Laguna Parón is currently the northernmost record for the genus Orobothriurus . The record from the Ishinca ravine is the world’s highest record for a scorpion.
ECOLOGY: The area inhabited by O. huascaran represents typical Puna vegetation with shrub steppe, grasses and small patches of Polylepis spp. forest (‘‘queñua’’ in Quechua), situated at 3800–4200 m (fig. 3F). All specimens were found under stones during the day. No other scorpion species have been recorded in the area.
REMARKS: The female specimen from Quebrada Yanganuco, listed under O. paessleri by Lourenço and Dastych (2001: 54), is probably conspecific with O. huascaran , n. sp., based on the known distribution of this species.





| MHNC |
Museo de Historia Natural de Concepcion (Chile) |
| MHNG |
Museum d'Histoire Naturelle |
| MRSN |
Museo Regionale di Scienze Naturali |
| MZSP |
Sao Paulo, Museu de Zoologia da Universidade de Sao Paulo |
| AMNH |
American Museum of Natural History |
| VSM |
Det Kgl. Norske Videnskabers Selskab Museet |
| ML |
Musee de Lectoure |
| LIM |
Severoceské muzeum |
| VM |
Okresní vlastivedné muzeum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Orobothriurus huascaran
| Ochoa, José A., Ojanguren Affilastro, Andres A., Mattoni, Camilo I. & Prendini, Lorenzo 2011 |
Orobothriurus sp. 2
| Ochoa, J. A. 2004: 50 |
Orobothriurus paessleri: Lourenço and Dastych, 2001: 54
| Lourenco, W. R. & H. Dastych 2001: 54 |
Orobothriurus crassimanus:
| Lourenco, W. R. & J. Qi 2006: 290 |
| Lourenco, W. R. 2003: 227 |
| Lourenco, W. R. 2002: 400 |
| Coddington, J. A. & R. K. Colwell 2001: 207 |
| Lourenco, W. R. 1997: 588 |
| Polis, G. A. 1990: 252 |