
Hemitaeniochromis brachyrhynchus, Oliver, Michael K., 2012
|
publication ID |
https://doi.org/10.5281/zenodo.213531 |
|
DOI |
https://doi.org/10.5281/zenodo.5612544 |
|
persistent identifier |
https://treatment.plazi.org/id/0B478787-BE4B-0E10-63B6-FF23FB904C93 |
|
treatment provided by |
Plazi |
|
scientific name |
Hemitaeniochromis brachyrhynchus |
| status |
sp. nov. |
Hemitaeniochromis brachyrhynchus View in CoL , new species
Figures 1–4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 , Tables 1 View TABLE 1 & 2 View TABLE 2
Hemitaeniochromis View in CoL sp. ' insignis big eye', Snoeks & Hanssens 2004: 284 & Fig. 52.
Holotype. YPM 25201, adult male, 123.6 mm SL. Malaŵi: Lake Malaŵi: Thumbi Island West, N side, approximately 14° 0 0′ 57″S, 34° 48′ 34″ E, trammel nets and experimental gillnets set in series perpendicular to shore over rocks possibly with sand in 17–38 m depth, 3–4 Jul. 1980; M.K. Oliver, K.R. McKaye, and T.D. Kocher. Field no. MKO 80–49.
Paratype. MRAC 99-41-P-1746, sex undetermined, 81.5 mm SL. Malaŵi: Lake Malaŵi: Nkhata Bay, south bay, 11° 36. 22′ S, 34° 18.16′ E, purchased from indigenous fishermen, 27 Sept. 1997; SADC /GEF. Field no. 97/20/ 114/7.
Diagnosis. A species of Hemitaeniochromis readily distinguished from its only formal congener, H. urotaenia , by its narrow lacrimal bone whose width, at the midpoint of the bone, is one-third (33.5–34.5%) of the orbit length (vs. nearly equal to the orbit length in H. urotaenia ); by its snout being shorter than the orbit length (vs. longer in H. urotaenia ); by its much larger eyes, the orbit length being about 36–38% of head length (vs. ~ 22–28% in H. urotaenia ); by its shorter lower jaw which is about 39–41% of head length (vs. ~ 45–51% in H. urotaenia ); and by its melanin pattern, which is composed of spots and line segments that are less well-defined than those of H. urotaenia (compare Figs. 1 View FIGURE 1 and 5 View FIGURE 5 ). The narrow lacrimal and large eyes also distinguish H. brachyrhynchus from all other species that have sometimes been placed in Hemitaeniochromis (see Discussion). Indeed, these two features in combination are probably unique to H. brachyrhynchus among all known Lake Malaŵi cichlids.
Description. Morphometric and meristic data are given in Tables 1 View TABLE 1 and 2 View TABLE 2 .
Body moderately elongate, dorsal profile evenly rounded from nuchal region of head to end of spinous dorsal fin, more convex than ventral body profile ( Fig. 1 View FIGURE 1 ). Head profile concave above snout. Jaws rather short, gape steeply inclined, premaxilla nearly vertical when mouth widely opened ( Fig. 2 View FIGURE 2 a); lower jaw apparently slightly projecting and inclined at ~50° to lateral midline when mouth is closed (in paratype; mouth of holotype cannot be fully closed due to stiffness of preservation). Lips slightly thickened but not lobate. Snout distinctly shorter than orbit, 1.28–1.33 in orbit length. Eyes large, orbit length ~36–38% HL. Lacrimal bone narrow, its width at mid-bone only about one-third orbit length, its orbital and labial margins nonparallel, converging nearer each other ventrally than at dorsal end of bone. Caudal fin emarginate.
Dental arcade of each jaw broad, semicircular in outline anteriorly ( Fig. 2 View FIGURE 2 c). Outer jaw teeth stout, the shafts conical; those of both jaws implanted with interspaces about as wide as the teeth in holotype (upper jaw, Fig. 2 View FIGURE 2 c, d; lower jaw, Fig. 3 View FIGURE 3 d), spaced more closely with interspaces about half as wide as teeth in paratype ( Fig. 3 View FIGURE 3 b). Paratype and holotype display what may be ontogenetic changes in crown morphology. Paratype anteriorly has 7 or 8 (upper jaw) and 9–12 (lower jaw) very unequally bicuspid teeth on each hemijaw, the cusps nearly conical and acute ( Fig. 3 View FIGURE 3 b); and smaller, unicuspid teeth, buried to the crowns in thickened mucosa, on posterior half of dental arcade. In contrast, holotype has uniformly unicuspid teeth with nearly conical crowns, although a few have crowns with a lateral “shoulder” ( Fig. 2 View FIGURE 2 d) in location of minor cusp of paratype’s teeth. Crowns of outer upper jaw teeth are slightly incurved in both specimens, as is usual in Lake Malaŵi haplochromines. However, in both specimens, the anterior teeth of the lower jaw have the crowns angled slightly forward and outward relative to their shafts ( Fig. 3 View FIGURE 3 b, d), an unusual condition. Inner teeth are tricuspid with acute median cusp in paratype, acutely unicuspid in holotype.
Lower pharyngeal bone (examined only in holotype; Fig. 2 View FIGURE 2 b) Y-shaped, delicate; all teeth laterally compressed, cuspidate, none enlarged (except those of posterior row near midline) nor submolariform. Median suture straight, not sinuous.
Gill rakers 10–12 on ceratobranchial, simple, unbranched, grading in length from longer posteriorly to short anteriorly; lightly pigmented with scattered melanophores.
Scales ctenoid. Lateral line discontinuous, the upper section kinked downward caudally so that the two sections are separated by a single scale without a canal or pore ( holotype, bilaterally) or by two scales, of which the upper has a pore but no canal and is situated below and behind the last true lateral line scale (with canal) of the upper segment, and the lower has neither pore nor canal ( paratype, bilaterally). Squamation extending onto caudal fin between fin rays, in paratype covering basal 60% of upper and lower lobes and basal 40% along middle rays, in holotype covering 90% of entire fin. Bases of soft dorsal and anal fins partly covered by 1–3 irregular rows of small scales.
Holotype Paratype YPM 25201 MRAC 99-41-P-1746 Holotype Paratype YPM 25201 MRAC 99-41-P-1746
Scales:
Coloration in life. Unknown.
Coloration in preservative ( Fig. 1 View FIGURE 1 ). Dorsum, nuchal region, head above eye, and upper surface of snout dark brown; remainder of head and body with nearly uniform tan or light brown ground color, becoming paler on hyoid area and branchiostegal membrane and silvery tan on belly. Lacrimal brown, lacking distinct stripe; operculum with dark spot; no other distinct head markings. Seven faint vertical bars below dorsal-fin base, darkest on upper body; traces of two further bars on caudal peduncle. Four to five indistinct dorsal midline spots below dorsal-fin base. Supralateral stripe situated just above upper lateral line, extending discontinuously between subdorsal bars 1–4, darkest at bar intersections and between bars 2 and 3. Midlateral stripe just above level of lower lateral line, partly discontinuous, originating at subdorsal bar 2 and extending to end of caudal peduncle where it forms a more or less discrete small precaudal spot; this stripe darkest at bar intersections and most nearly continuous above anal fin. Dorsal fin lacking a submarginal stripe; soft dorsal marbled with darker and lighter brown markings. Caudalfin membrane light to medium brown, without evident maculae. Anal fin nearly uniform brownish; holotype with hints of ~3 elongate paler spots with darker margins on posterior half of fin. Pectoral hyaline; pelvic brownish on anterior half, unpigmented on posterior 2 or 3 rays.
Habitat and distribution. Almost nothing is known. Two large collecting efforts ( MKO and colleagues, Jun.–Aug. 1980, 133 stations; SADC/GEF, 1990s, see Duponchelle & Ribbink 2000), each obtaining thousands of fishes from varied habitats, localities, and depths with a range of techniques, collected only a single H. brachyrhynchus specimen apiece. The two specimens are from localities 270 km apart ( Fig. 4 View FIGURE 4 ), indicating that H. brachyrhynchus is widely distributed. The depths of capture of the two specimens are uncertain. The holotype was caught in an unspecified part of a set of demersal trammel nets and experimental gillnets, of various mesh sizes, totaling 183 m in length, which had been set in series overnight on the bottom (over rocks possibly with areas of sand) where the depth extended from 17– 38 m. The stomach of the holotype was everted when the specimen was brought to the surface, suggesting that it came from relatively deep water. The paratype was purchased at Nkhata Bay from artisanal fishermen; its labels do not record the method of capture, but I speculate that it might have been angled on a longline from a dugout canoe as this is a popular indigenous fishing technique at Nkhata Bay. This species does not seem to be represented in the most complete photographic atlas of Malaŵi cichlids from all habitats ( Konings 2007). [An entity labeled Hemitaeniochromis sp. ‘ spilopterus blue’ by Konings is, to judge by the single male and female he illustrates, reminiscent of H. brachyrhynchus but appears to have a distinctly wider lacrimal ~55% of orbit length (vs. 33.5–34.5% in H. brachyrhynchus ) and has the midlateral stripe (clearly visible in the female) continuously pigmented, not broken into spots, although it is slightly darker where crossed by the vertical bars as in other striped species. I would consider it a Protomelas .] I am not aware of any H. brachyrhynchus specimens resulting from the extensive experimental trawling program in the southern lake; no specimens were illustrated or described in Turner’s (1996) book on offshore cichlids. The scant available information thus suggests that H. brachyrhynchus is a widely distributed but rare species, apparently occurring around rocks in rather deep water. Although it has not been possible to examine stomach contents, the distinctive lower-jaw teeth, with their outwardly and anteriorly angled crowns, are similar to those of other haplochromine cichlids, of several lineages, that have a paedophagous diet (see Discussion).
Conservation status. Unknown. Hemitaeniochromis brachyrhynchus may normally inhabit deeper water than has generally been sampled over rocky substrates. The widely separated localities of the two type specimens at least demonstrate that the species is not restricted to a single circumscribed area.
Etymology. Latinized from the Greek adjective βραχύσ, short, and noun ρύγχοσ, snout, in reference to the abbreviated preocular region of the head; brachyrhynchus is a noun in apposition. No indigenous names have been recorded.
Discussion. Hemitaeniochromis was proposed for a single species, H. urotaenia ( Fig. 5 View FIGURE 5 ). The genus was diagnosed “…by having a boldly marked melanin pattern consisting of a mid-lateral band confined to the posterior half of the flank but continued forwards as a series of spots and a supralateral band broken into a series of spots. Mouth large, lower jaw 2.0 to 2.2 in head length, with well developed mental region. Outer teeth unicuspid, slightly recurved, separated by spaces approximately equal to tooth diameter” ( Eccles & Trewavas 1989: 71). In couplet 57 of their key to genera, these authors noted further that Hemitaeniochromis (together with Tyrannochromis , which has a very similar melanic pattern but, because of its beaklike jaws and laterally compressed head, is not likely to be confused with Hemitaeniochromis ) are “[l]arge-mouthed predators” with “… teeth all unicuspid in fish over 100 mm SL, sometimes very unequally bicuspid in smaller individuals.” Although not mentioned in the diagnosis, it is also noteworthy that the interrupted midlateral stripe begins about 5 scales behind the operculum, instead of immediately behind it as in the plesiomorphic haplochromine pattern. However, this apomorphic condition is not unique to Hemitaeniochromis , being found also in some species currently assigned to Protomelas , in Tyrannochromis macrostoma ( Regan, 1922) , and in some individuals of Lethrinops lethrinus (Günther, 1894) .
Hemitaeniochromis brachyrhynchus fits into this original diagnosis of Hemitaeniochromis well in some respects, including the apomorphic discontinuous midlateral stripe beginning well behind the operculum and the spaced, unicuspid adult jaw teeth. The exceptional characters are the smaller mouth (lower jaw 2.45–2.54 in head), the less well-defined midlateral stripe (compare Figs. 1 View FIGURE 1 and 5 View FIGURE 5 ), the outwardly curved crowns of the anterior lower jaw teeth, and the more closely spaced jaw teeth of the paratype (but not the holotype, a larger specimen). At present, without stronger phylogenetic evidence, Hemitaeniochromis appears to be the most appropriate genus for this new species, as also indicated by Snoeks & Hanssens (2004). However, except for the specialized midlateral stripe, no convincing synapomorphy shared with H. urotaenia is evident from external examination of the H. brachyrhynchus type specimens.
The unusual forward and outward curvature of the anterior tooth crowns in the lower jaw of H. brachyrhynchus is also seen in a few other haplochromine cichlids. Similar dentition characterizes the known paedophages (specialist feeders on eggs and larvae, usually of other cichlids) of the genus Caprichromis and the suspected paedophage Protomelas spilopterus ( Trewavas, 1935) ( Eccles & Trewavas 1989: 69) , both endemic to Lake Malaŵi, as well as Haplochromis obesus (Boulenger, 1906) , H. maxillaris Trewavas, 1928 , and H. melanopterus Trewavas, 1928 of Lake Victoria, which are also known or suspected paedophages ( Greenwood 1959). A heavily built lower jaw, and teeth partly buried in thickened mucosa are also features common to H. brachyrhynchus and all these fishes. It seems reasonable to hypothesize that H. brachyrhynchus is another convergently specialized paedophage, which may explain why its jaws, dentition, and snout anatomy differ so strikingly from those of its congener, H. urotaenia . Stomach contents were not examined in the two known specimens, the holotype having had its stomach everted upon capture, as noted above; the belly of the paratype remains unslit.
The nominal genus most similar to Hemitaeniochromis in melanin pattern and general habitus is Protomelas . Eccles and Trewavas (1989) provide no actual diagnostic characters in their diagnosis of the latter genus, even stating “No synapomorphies have been recognised separating Protomelas from the other Malaŵian genera, which are distinguished by their own synapomorphies.” Thus, Protomelas is a genus of convenience rather than a lineage, a catch-all for numerous more or less similarly marked, relatively plesiomorphic Lake Malaŵi haplochromines. Included in Protomelas by Eccles and Trewavas were species “with moderate sized mouths” and a “[m]elanin pattern based on the plesiomorphic, with the longitudinal bands usually predominating.” Some species, including the type species P. k i r k i i (Günther, 1894) ( Fig. 6 View FIGURE 6 ), have a complete midlateral stripe originating close behind the operculum. In others [e.g., P. insignis ( Trewavas, 1935) and P. spilonotus ( Trewavas, 1935) ] this stripe begins well behind the operculum as in Hemitaeniochromis ; some other Protomelas species are variable in this feature. The jaw dentition of included species is “…usually bicuspid, at least in young, but may be replaced in adults by unicuspids which, however, are not widely spaced” ( Eccles & Trewavas 1989). The chief characters excluding H. brachyrhynchus from this broadly defined Protomelas and instead aligning it with H. urotaenia are the interrupted, spotted midlateral stripe and the widely spaced jaw teeth in the adult holotype.
Two of the species that Trewavas (1935) had originally described in Haplochromis , H. spilopterus and H. insignis , were among the forms assigned by Eccles and Trewavas (1989) to their new genus Protomelas . Both have been referred to Hemitaeniochromis by some recent authors.
The first of these, P. spilopterus , is a moderately common large-eyed cichlid of sandy shores and intermediate (sand/rock) areas, recognizable in the field by its obliquely inclined mouth, heavy lower jaw with dentary more than twice as deep as the premaxilla, and continuous midlateral stripe (exceptionally broken into separate spots) originating close behind the operculum ( Fig. 7 View FIGURE 7 ). Its jaw teeth are unequally bicuspid in young individuals, becoming mostly unicuspid in adults, the lower-jaw teeth having the crowns angled slightly forward. Although this species has not been observed to feed, it is strongly suspected of being a paedophage ( Eccles & Trewavas 1989; Konings 2007). Konings (2007: 344) remarks, “…in my opinion H. spilopterus belongs to [ Hemitaeniochromis ] as it shares all its morphological characteristics. The only discrepancy is that the mid-lateral stripe is continuous in most individuals….”
The other species, P. insignis , is a rather uncommon intermediate-zone inhabitant with a midlateral stripe originating at least an eye diameter behind the operculum. This stripe is most often continuous, but may be divided into semi-discrete spots ( Fig. 8a View FIGURE 8. a ). The outer jaw teeth are predominantly unicuspid in adults, but occasional teeth with bicuspid crowns are retained. Protomelas insignis has been observed to feed by stealing eggs from the nests of other sandy-shore cichlids in nesting arenas, especially where multiple species are nesting ( Lewis et al. 1986: 39). These authors, and Konings (2007), report the same unusual behavior in Otopharynx ovatus , a cichlid with three lateral spots ( Fig. 8 View FIGURE 8. a b). Is this mere coincidence? While re-examining one lot with five P. insignis specimens (YPM 14291) in preparing this paper, I was intrigued to find in it a note I had written many years ago ( 26 Feb. 1981): “NB—are insignis and ovatus a single sp.?—similar behavior; color pattern of latter suggested in that of former. Similar phar[yngeal]s, dentition. Do they differ in anything but color pattern?” Interestingly, Turner (1996: 196) independently developed a similar suspicion that O. ovatus is conspecific with what he discussed and illustrated as Hemitaeniochromis ‘insignis’, a cichlid with midlateral and supralateral stripes (although he referred to the latter species as an “oblique-striped paedophage,” apparently a lapsus since both his text and his photo depict a cichlid with two horizontal stripes, not an oblique stripe). Turner remarks that “In the confusion of an attack on a spawning pair of Copadichromis , one [of] these fishes appeared to change between the spotted and striped pattern.” Thus, if Turner’s observations are correct, what were originally thought to be two separate and differently marked species, not closely related to each other, may be phases of a single species, which may even be capable of rapidly altering its melanin pattern to facilitate aggressive mimicry of various cichlid prey species with either striped or spotted melanic patterns. However, well before both my observations and Turner’s, it was David Eccles who first suspected that P. insignis and O. ovatus might be the same species. This is documented by his note in the jar containing (in 1972, when I examined them) the syntypes of Haplochromis ovatus Trewavas, 1935 , BMNH 1935.6.14.1487- 1489. Eccles’ note reads: “ H. insignis , H. ovatus , H. obtusus appear to be conspecific. D.H. Eccles Oct 1969.” (I had seen this notation, but did not consciously remember having read it, when I made my own note of the possible synonymy of H. insignis and H. ovatus 9 years later in 1981.) Eccles’ same note was repeated in the lot of H. insignis types, BMNH 1935.6.14.839-843; and in that of the H. obtusus holotype, BMNH 1935.6.14.1453. Nevertheless, Eccles & Trewavas (1989) later treated all three as valid species, even assigning them to three different genera: Protomelas insignis , Otopharynx ovatus , and Maravichromis obtusus (the last of these genera synonymized with Mylochromis Regan, 1920 by Derijst & Snoeks 1992). The question of the possible synonymy of P. insignis and O. ovatus , species with different color patterns and currently classified in different genera, remains of considerable biological interest, but its resolution is outside the scope of this study.
In addition to the previously described species Protomelas spilopterus and P. insignis , several presently undescribed species are also believed by a few workers to belong to Hemitaeniochromis . Extensive lake-wide exploration of littoral habitats using SCUBA, notably by Adrianus Konings, has resulted in the discovery and photographic recording of at least seven additional forms that he refers to Hemitaeniochromis ( Konings 2007) . Unfortunately, no museum specimens of these species are available for study.
All eleven species that have been referred to Hemitaeniochromis are listed and briefly characterized in Table 3 View TABLE 3 . However, there are two problems with assigning all the species in Table 3 View TABLE 3 to Hemitaeniochromis . First, the midlateral stripe, whose apomorphic configuration is a key feature in the definition of the genus, varies widely among these species, differing in its point of origin, width, degree of continuity, and even presence or absence. Second, nearly all of the tabulated species are either known to be, or suspected of being, paedophages, with evident adaptations in their jaw structure, dentition and, presumably, behavior for this specialized mode of predation. In contrast, the type species, H. urotaenia , lacks these specializations and is known to be a “standard” generalized piscivore ( Eccles & Trewavas 1989). Given these two problematic character distributions, attempting to force all the varied taxa represented in Table 3 View TABLE 3 into Hemitaeniochromis would seem to render the genus undefinable and, very probably, polyphyletic. Perhaps some or all of the paedophagous species among them represent a single lineage (although, given their heterogeneous markings, this is open to doubt), which might even require a new genus. This, however, cannot be determined until specimens are available for detailed study. How should the various tabulated species be provisionally classified in the meantime?
placement. Most data were gleaned from published text and photographs in the cited sources; the author is aware of no museum
specimens representing any of the undescribed species.
Species Midlateral stripe Diet Authority for placement
in Hemitaeniochromis Origin Modal structure
Taxonomic recommendations. (1) I propose that Protomelas insignis and P. spilopterus be retained in Protomelas . Even though that genus is not demonstrably monophyletic, transferring the two species to Hemitaeniochromis would only serve to render the latter genus nonmonophyletic as well, since they share no synapomorphies with H. urotaenia , the type species. (2) Until voucher specimens of those species now known only from underwater photographs become available and are taxonomically studied, Hemitaeniochromis should be restricted to H. urotaenia and H. brachyrhynchus . These two species do share the putative synapomorphy of a midlateral stripe that originates well behind the operculum and that is discontinuous, composed both of spots and of line segments. (3) The “photo species” are, in effect, incertae sedis within the Lake Malaŵi non-mbuna haplochromines and should be referred to as species of ‘ Hemitaeniochromis ’ (in single or double quotation marks) to show that they are not necessarily related closely to H. urotaenia ; thus, ‘ Hemitaeniochromis ’ sp. ‘paedophage’, ‘ Hemitaeniochromis ’ sp. ‘ spilopterus yellow’, etc.
Of the undescribed species in Table 3 View TABLE 3 , only ‘ Hemitaeniochromis ’ sp. ‘ insignis mumbo’, ‘ H. ’ sp. ‘ spilopterus kande’, and possibly ‘ H. ’ sp. ‘ urotaenia tanzania’ appear to fall within Hemitaeniochromis as here defined (although the form of their teeth is not known). The other species in Table 3 View TABLE 3 lack the derived states of the midlateral stripe found in H. urotaenia and H. brachyrhynchus and should be excluded from Hemitaeniochromis proper. For example, in ‘ H ’. sp. ‘ spilopterus blue’, to judge by the female depicted by Konings (2007: 167 Fig. 2 View FIGURE 2 ), the midlateral stripe originates well behind the operculum but retains the plesiomorphic unbroken configuration seen in some Protomelas , as noted above, instead of being broken into spots as in Hemitaeniochromis .


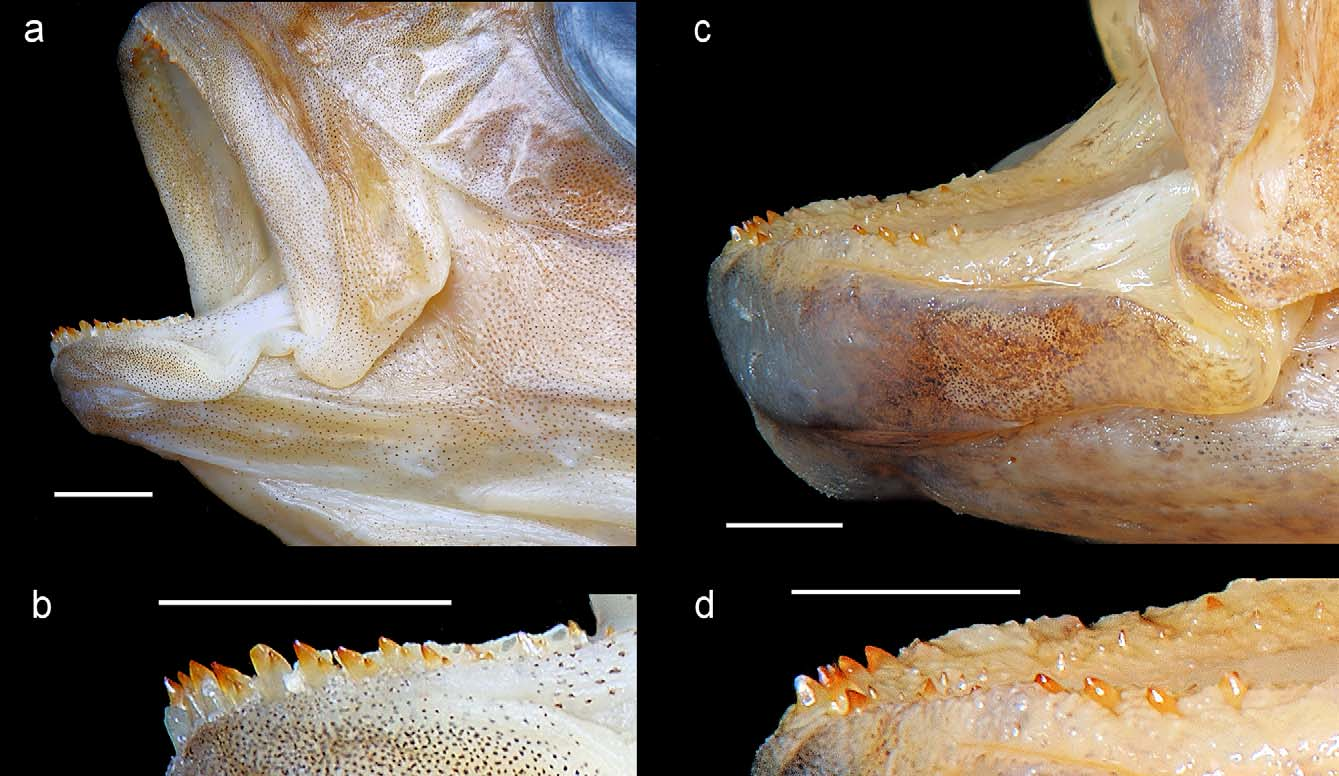
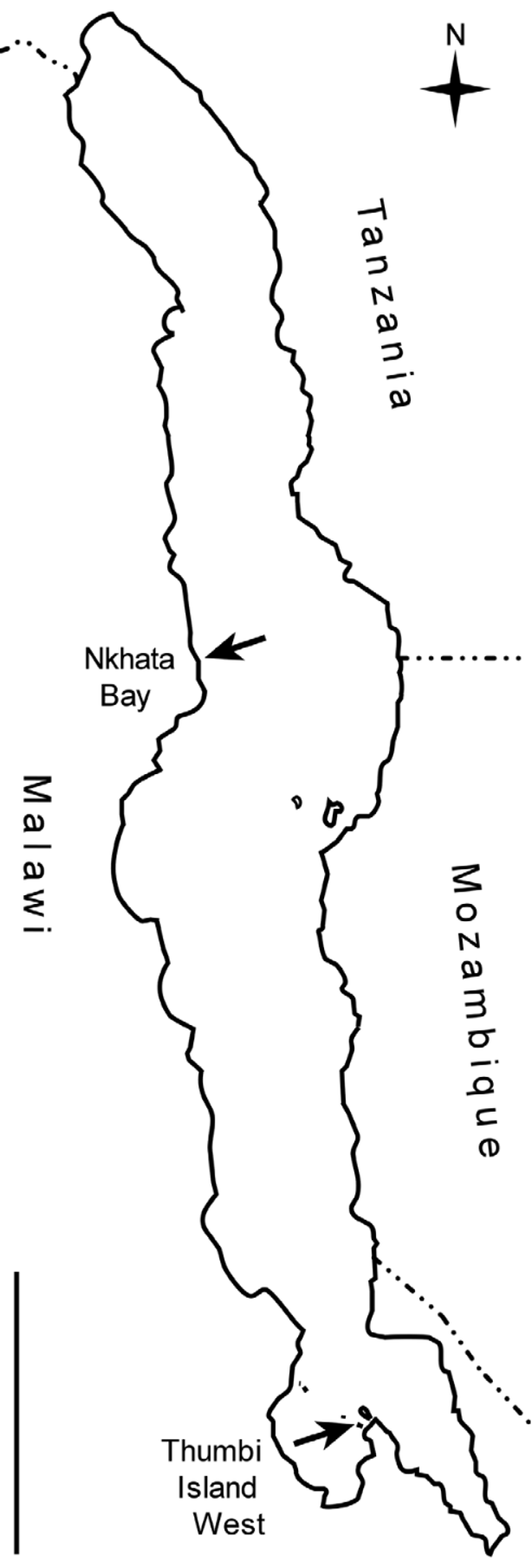




TABLE 1. Morphometric characters of Hemitaeniochromis brachyrhynchus. Measurements preceded by a tilde (~) have large uncertainty because of the specimens having been fixed with the mouth open.
| Standard length (mm) | 123.6 | 81.5 |
|---|---|---|
| Head length (mm) | 42.1 | 28.7 |
| % of standard length: | ||
| Head length | 34.1 | 35.2 |
| Body depth | 35.4 | 37.5 |
| Dorsal-fin base length | 53.3 | 54.6 |
| Predorsal length | 35.0 | 37.4 |
| Prepectoral length | 37.6 | 36.2 |
| Prepelvic length | 49.1 | 46.0 |
| Preanal length | 72.4 | 72.9 |
| Belly length | 25.9 | 30.3 |
| Anal-fin base length | 19.0 | 19.5 |
| Caudal peduncle length | 17.1 | 13.6 |
| Caudal peduncle depth | 12.4 | 13.0 |
| Pectoral-fin length | 33.6 | 35.8 |
| Pelvic-fin length | 28.3 | 25.6 |
| % of head length: | ||
| Head width | 49.4 | 46.0 |
| Interorbital width | 25.9 | 22.0 |
| Snout length | 28.7 | 28.2 |
| Snout width | ~35.6 | ~29.3 |
| Lower jaw length | 40.9 | 39.4 |
| Lower jaw width | ~38.4 | ~25.8 |
| Premaxillary pedicel length | 25.4 | 27.9 |
| Upper jaw length | ~39.0 | ~34.1 |
| Cheek depth | 21.1 | 16.7 |
| Orbit length | 38.2 | 36.2 |
| Vertical eye diameter | 33.3 | 32.8 |
| Lacrimal (preorbital) depth | 12.8 | 12.5 |
| Postorbital head length | 34.7 | 35.9 |
| Pharyngeal bone overall length | 25.9 | — |
| Pharyngeal bone fork length | 19.7 | — |
| Pharyngeal bone width | 25.4 | — |
| Pharyngeal toothplate overall length | 14.3 | — |
| Pharyngeal toothplate fork length | 11.6 | — |
| Pharyngeal toothplate width | 18.1 | — |
| Angular measurements in degrees: | ||
| Premaxillary pedicel | 5 | 10 |
| Interorbital | 30 | 30 |
| Nuchal region | 5 | 15 |
| Lower jaw underside | — | ~50 |
| Gape inclination | ~60 | ~60 |
TABLE 2. Meristic characters of Hemitaeniochromis brachyrhynchus.
| Lateral line scales | 33 | 33 |
|---|---|---|
| Upper lateral line scales | 27 | 22 |
| Lower lateral line scales | 17 | 13 |
| Lateral line scales on caudal fin | 2 | 1 |
| Upper transverse line scales | 5 | 5 |
| Lower transverse line scales | 12 | 11 |
| Predorsal scales | ~16 | ~13 |
| Prepelvic scales | ~22 | ~17 |
| Belly scales | ~25 | ~23 |
| Cheek scale rows | 3 | 3 |
| Scales between pectoral- and pelvic-fin bases | 7 | 8 |
| Scales around caudal peduncle | 16 | 16 |
| Fins: | ||
| Dorsal-fin spines, segmented rays | XVII, 10 | XVII, 10 |
| Anal-fin spines, segmented rays | III, 9 | III, 9 |
| Pectoral-fin rays | 14 | 14 |
| Gill rakers: | ||
| Epibranchial/angle/ceratobranchial | 3/1/12 | 4/1/10 |
| Teeth: | ||
| Outer upper jaw teeth left/right | ~22/~22 | 19/18 |
| Inner rows upper jaw | 1 | 1 |
| Outer lower jaw teeth left/right | 15/13 | 16/15 |
| Inner rows lower jaw | 2 | 2 |
| Lower pharyngeal teeth, posterior edge | 28 | — |
| Lower pharyngeal teeth, median column | 8–9 | — |
| Lower pharyngeal teeth, oblique count | 6 | — |
TABLE 3. Species that have been referred to Hemitaeniochromis, with selected characteristics and authority for this generic
| Hemitaeniochromis brachyrhynchus Oliver | Far behind operculum | Spotted anteriorly, more continuous poste- riorly | Unknown (pae- dophage?) | Oliver (this paper); Snoeks & Hanssens (2004) |
|---|---|---|---|---|
| Hemitaeniochromis insig- nis (Trewavas) = H. sp. ‘insignis’ | Far behind operculum | ± Continuous, wide | Steals eggs from nests (Lewis et al. 1986) | Turner (1996); Duponchelle & Ribbink (2000); Snoeks & Hanssens (2004) |
| Hemitaeniochromis sp. ‘ insignis mumbo’ | Far behind operculum | Discontinuous, wide | Unknown; strong lower jaw as in pae- dophages | Konings (2007) |
| Hemitaeniochromis sp. ‘paedophage’ | Far behind operculum | Continuous, narrow | Cichlid eggs, larvae (paedophage) | Konings (2007) |
| Hemitaeniochromis spi- lopterus (Trewavas) | Close to oper- culum | Continuous, ± wide | Paedophage? | Konings (2007) |
| Hemitaeniochromis sp. ‘ spilopterus blue’ | Far behind operculum | Continuous, wide | Cichlid & catfish eggs & larvae (paedophage or opportunistic?) | Konings (2007) |
| Hemitaeniochromis sp. ‘ spilopterus jalo’ | — | None? | “Strong jaws suggest a predatory lifestyle” | Konings (2007) |
| Hemitaeniochromis sp. ‘ spilopterus kande’ | Far behind operculum | Spotted anteriorly, more continuous poste- riorly | Unknown | Konings (2007) |
| Hemitaeniochromis sp. ‘ spilopterus yellow’ | Far behind operculum | Continuous, ± wide | Cichlid eggs, larvae (paedophage?) | Konings (2007) |
| Hemitaeniochromis uro- taenia (Regan) (type of the genus) | Far behind operculum | Spotted anteriorly, more continuous poste- riorly | Piscivore | Eccles & Trewavas (1989) |
| Hemitaeniochromis sp. ‘ urotaenia tanzania’ | Far behind operculum | Discontinuous, wide, with spots like those of Otopharynx brooksi | Seen to hunt small fishes | Konings (2007) |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Hemitaeniochromis brachyrhynchus
| Oliver, Michael K. 2012 |
Hemitaeniochromis
| Snoeks 2004: 284 |