
Juliomys pictipes ( Osgood, 1933 )
|
publication ID |
https://doi.org/ 10.5281/zenodo.181855 |
|
DOI |
https://doi.org/10.5281/zenodo.5623613 |
|
persistent identifier |
https://treatment.plazi.org/id/0C1487A9-8E66-5A72-FF08-FDFA5D82D78B |
|
treatment provided by |
Plazi |
|
scientific name |
Juliomys pictipes ( Osgood, 1933 ) |
| status |
|
Juliomys pictipes ( Osgood, 1933) View in CoL
Holotype. FMNH 26814, adult male (skin and skull) collected by C. C. Sanborn on Sept. 6, 1926. Photographs (skull and mandible) of this individual were recently published by Costa et al. (2007: figs. 4 and 5).
Type locality. “Caraguatay, Rio Parana, 100 miles south of Rio Iguassu, Misiones, Argentina ” ( Osgood, 1933:11). The exact location of this type locality was recently addressed by Pardiñas et al. (2007:397). These authors, taking into account the most probable route followed by C. Sanborn and the fact that he worked in Misiones’ Charles Benson property (see Field Museum of Natural History - Reports, 1927), restricted it to Puerto Caraguatay (26º 37’ S, 54º 46’ W, Department of Montecarlo, Province of Misiones). However, in a recent inspection of this area, Mr. Benitez, an old resident of the area and with a good knowledge of it, showed us ( UFJP and CAG) the exact place where the original house of Mr. Benson was emplaced. The main building is now in ruins and practically subsumed by the forest. Clearly, this is not Puerto Caraguatay, instead Puerto Benson with coordinates 26º 37’ 13” S and 54º 46’ 57” W and probably Sanborn trapped around there. Despite those findings, the extreme proximity of these two localities (ca. 0.5 km), the missing of Puerto Benson as toponomy, and the ambiguity around the exact place where Sanborn worked, support our restriction of the type locality of J. pictipes to Puerto Caraguatay.
Distribution ( Fig. 2 View FIGURE 2 ). J. pictipes is restricted to tropical and subtropical moist forests of the Atlantic coast in southeastern Brazil and in interior subtropical moist forests of Argentina ( Osgood, 1933; Pine, 1980; Costa et al., 2007).
New Argentinean specimens ( Fig. 2 View FIGURE 2 ). CIES-M 23, adult female preserved as skin in poor condition and skull, originally identified as Oligoryzomys nigripes . Collected by Silvana Montanelli in Sendero Macuco, Parque Nacional Iguazú (25º 41’ S, 54º 26’ W, Department of Iguazú, Province of Misiones). CNP 895, young female, preserved as cleaned skull, carcass in fluid and digestive, collected by Ulyses Pardiñas and Rosario Robles (original number LTU 379) in Arroyo de Salamanca, Parque Provincial “Ernesto Che Guevara” (26º 36’ 53” S, 54º 46’ 51” W, 147 m asl, Department of Montecarlo, Province of Misiones). MLP 1.I. 03.24, adult pregnant female (3 fetus) preserved as complete fluid, except digestive organs, collected by Graciela Navone and Juliana Notarnicola on 28 August 2006 (original number JN 702) in Balneario de la Reserva Privada de Usos Múltiples de la Universidad Nacional de La Plata “Valle del Arroyo Cuña Pirú” (27º 05’ S, 54º 57’ W, Department of Cainguas, Province of Misiones).
Emended diagnosis. The largest known species of Juliomys , characterized by the following combination of traits: body markedly bicolored light orange-brown above and white to cream-white below; tail slightly shorter than head-body length; tail bicolor except for the terminal half-inch, which is dusky all around; feet clear ochraceous-twany, the toes whitish; zygomatic notch moderately expressed; upper free border of the zygomatic plate reduced; incisive foramina short, not reaching the first upper molars; lateral expansion of frontal bones restricted; tympanic bulla small, squamosal-alisphenoid and sphenofrontal foramen absents, sphenopalatine vacuities absents or reduced to a narrow fissures, 2n = 36.
Description. Based on the new Argentinean specimens. A small-sized sigmodontine rodent, externally similar to Oligoryzomys nigripes , but with the head and eyes proportionally larger and with a shorter tail that is nearly equal to head and body ( Fig. 3 View FIGURE 3 ). Dorsal hairs are plumbeous gray at the base and distally reddish. Over hairs on the rump and hind legs are vivid orange at the tip, contrasting with the rest of the body. The venter is covered by bicolored hairs; each hair has a short gray base and is withish at the tip. Orange tipped hairs are visible in the inguinal region around the tail base. The philtrum, genal zone, and chin are covered by whitish hairs. Hairs around the nose are short and orange, contrasting with the rest of the body. The ears are small, covered mainly with short and sparse brownish hairs externally and mostly naked internally; the ear border is covered by short reddish hairs. Mystacial vibrissae are blackish, relatively short anteriorly, and long posteriorly, the latter largely extended beyond the posterior border of the ears. The supercilliary and genal vibrissae are thinner, blackish, and shorter than mystacial, and not surpassing the posterior border of the ears. The tail pelage is sparse, gradually becoming denser toward the tip, where a short tuft is visible. Hairs from proximal and distal halves of tail reach 2–3 scales and 3–6 scales, respectively, with three hairs emerging from each scale. Tail hairs are brown on the dorsum and rufous in the venter, except in the distal tip, which is uniformly brown. Eight mammae are present in a pregnant female examined (MLP 1.I. 03.24) that are arranged in one inguinal, one abdominal, one postaxial and one pectoral pair (see Voss & Carleton, 1993). The hands and feet are short, covered with orange hairs dorsally; the fingers are covered by short white hairs ( Fig. 4 View FIGURE 4 b). The hallux is much shorter than other pedal digits. Manal and pedal pads boulbous and enlarged, especially those of the distal phalanxes, remembering the pads of arboreal sigmodontine species of the genus Oecomys and Rhipidomys ( Hershkovitz, 1960; Rivas & Linares, 2006). Three large and rounded interdigital and two large and mostly ovale carpal pads are present ( Fig. 4 View FIGURE 4 a). The plantar surface of the hind feet has six bulbous tubercles, four interdigitals and two tarsal. The interdigital ones are nearly ovale and large. The thenar pad is more rounded and smaller than the interdigital tubercles. The hypothenar pad is large and is comma-shaped. The distal edge of the thenar just reaches the proximal edge of the hypothenar and is positioned approximately midway along the sole of the foot ( Fig. 4 View FIGURE 4 c). The skin between distal pads up to the thenar and hypothenar pad limit is covered by small and rounded scales, while the remainder of the plantar surface is smooth ( Fig. 4 View FIGURE 4 a, c). The claws on the hind feet are short and darker in color than the skin of the pes. Ungual tufts are grayish-white and large, reaching or slightly surpassing the claws.
The skull is robust with a short rostrum, a rounded braincase and a marked domed profile in its posterior region (Fig. 5,6A-L). The nasals diverge anteriorly, without contrasting expansions and with a slight projection anterior to the incisors, conforming a moderate tube ( Fig. 5 View FIGURE 5 ). The nasofrontal suture is straight but markedly denticulate, positioned anteriorly to the lacrimal level ( Fig. 6 View FIGURE 6 A). A clear central depression is present in the anterior part of the interfrontal region ( Fig. 6 View FIGURE 6 E); in addition, in this region the interfrontal suture is slightly open, contrasting with the condition of this suture posterior to the depression which appears clearly closed in adult individuals ( Fig. 6 View FIGURE 6 B, C). Following Weksler (2006:28, character 22) the interorbital region can be typified as symmetrically constricted with squared supraorbital margins ( Fig. 6 View FIGURE 6 B, C). The lacrimal bone is large. The coronal suture is V-shaped and the squamosal anterior-upper portion is clearly visible from above over the braincase margins ( Fig. 6 View FIGURE 6 B). Both parietal and interparietal sutures are meandrous, and the former suture includes supernumerary middle bones ( Fig. 6 View FIGURE 6 I). The interparietal is wide and broad ( Fig. 6 View FIGURE 6 I). The occipital region is dorso-ventrally compressed producing a oblique position of the foramen magnum ( Fig. 6 View FIGURE 6 H). Lateral parietal and lambdoid crests are inconspicuous ( Fig. 5 View FIGURE 5 ). The zygomatic plate is high and narrow showing a much reduced upper free border and having a masseteric tubercle well developed in their base ( Fig. 6 View FIGURE 6 D). Contrasting with the general solid construction of the skull, the zygomatic arches, well expanded, are gracile ( Fig. 5 View FIGURE 5 ). A moderate postorbital ridge is present. The lateral portion of the parietals is reduced ( Fig. 6 View FIGURE 6 H). Ventrally, the incisive foramina are wide and moderately long and their posterior ends almost reach the M1 anterior face ( Fig. 5 View FIGURE 5 , 6 View FIGURE 6 G). The palate is bombed and finely foraminate with two posterior palatine foramina and paired posterolateral palatal pits side by side the mesopterygoid fossa anterior region ( Fig. 6 View FIGURE 6 F). The mesopterygoid fossa is broad and has a lire-shape ( Fig. 6 View FIGURE 6 F). The mesopterygoid roof is almost totally ossified; the sphenopalatine vacuities remain as narrow fissures ( Fig. 6 View FIGURE 6 G). The parapterygoid fossae are broad, well ossified, and show the small posterior openings of the alisphenoid canal ( Fig. 6 View FIGURE 6 G). The anterior ends of the parapterygoid fossae are located at the level of M3 protocones ( Fig. 6 View FIGURE 6 F). Otic capsules are reduced and auditory meatus are clearly enlarged ( Fig. 6 View FIGURE 6 J). The carotid canal is big and without pteriotic participation; the stapedial foramen is minute and the petrotympanic fissure absent ( Fig. 6 View FIGURE 6 J). The tegmen tympani is well developed, superposed to the squamosal bone, and partially filling the postglenoid foramen ( Fig. 6 View FIGURE 6 K). The narrow hamular process is distally spatulated and directly applied on the mastoid ( Fig. 6 View FIGURE 6 K). The mastoid capsule is small but not flattened with a large mastoid fenestra. Cranial foramina related with cephalic arterial pattern include a well developed oval foramen and a small anterior opening of the alisphenoid canal; the alisphenoid strut is absent, and the same is true for the squamosal-alisphenoid groove while the trough for the masticatory-buccinator nerve is shallow but well expressed ( Fig. 6 View FIGURE 6 G, K). These traits coupled with the absence of the sphenofrontal foramen characterizing the carotid circulatory pattern 2 of Voss (1988).
The mandible is robust and short ( Fig. 7 View FIGURE 7 A–B). The anteriormost point of the diastema is well below the alveolar plane ( Fig. 7 View FIGURE 7 A). The mental foramen is scarcely visible from lateral view. Both upper and lower ridges of masseteric crest are clearly expressed; the same is true for the capsular projection of the incisor. The angular process is short with respect to the condyloid process ( Fig. 7 View FIGURE 7 B).
Upper incisors are robust and deep, opisthodont, with bright orange frontal enamel. Both upper and lower molars are brachyodont, crested and with the main cusps slightly alternate. M1 ( Fig. 6 View FIGURE 6 L): subrectangular in outline; main cusp slightly alternating; procingulum almost wide as the remainder portion of the tooth; deep and penetrating anteromedian flexus with their fund engulfed producing two conules being the lingual smaller than labial one; anteroloph and mesoloph transverses in orientation with their labial tips spatulated and showing medium point-connections to the anterior structures (anterolabial conule and paracone, respectively) like displaced paralophules; short posteroloph transverse in orientation; very wide and penetrating protoflexus, subequal in morphology to hypoflexus; protostyle and enterostyle present, the latter with a short enteroloph (developed also in the young individual CNP 895). M2: subcuadrangular in outline; gross morphology very similar to the M1 comparable portion; main cusp something more alternating than M1. M3: about 0.7 of the M2 length; anteroloph conspicuous. Lower molars are crested and the main cusp slightly alternating. m1: procingulum wide; anteromedian flexid well developed producing subequal conulids; anterolophid transverse in orientation with tendency to fusion to anterolingual conulid; protoflexid externally obliterate by a sharp anterolabial cingulum; mesolophid narrow but independent and clearly developed well projected lingually; broad hypoflexid showing a small but marked ectostylid. m2: quadrangular in outline and subequal to the m1 comparable portion; mesolophid well developed; ectolophid plus ectostylid. m3: almost large as m2 (0.9); hypoflexid flanqued by oblique proto- and hypoconid walls resulting in a triangular outline, with their fund oriented to a bifid structure composed by fused mesolophid plus entoconid; annular posterofossetid.
We examined the stomach gross anatomy of one juvenile specimen (CNP 895). The morphology is of the type unilocular hemiglandular (see Carleton, 1973); in this morphology the glandular epithelium from the antrum extends to the corpus, near the esophagus opening ( Fig. 8 View FIGURE 8 ). Measures of the thick intestine and caecum are 3.5 and 1.8 cm, respectively.
Measurements. See Table 2 View TABLE 2 .
a Holotype; measurements taken from Osgood (1933:12) Habitat. All specimens were collected in the Interior Atlantic Forest of the Misiones Province, Argentina. This biotic unit, also known as “Selva Paranaense,” is part of the South American Atlantic Forest macroregion (Plací & Di Bitetti, 2005). Three of four specimens were collected in the vicinity of watercourses of the Paraná river basin. One individual from Sendero Macuco, Parque Nacional Iguazú, was collected in a mature forest of “Laurel and Guatambu” (Martínez-Crovetto 1963), where the predominant arboreal species were Matayba elaeagnoides , Balforoundendron riedelianum , Sorocea ilicifolia , Cordia ecaliculata , and Prumus sphaerocarpa. Dense underbrush vegetation at Sendero Macuco includes the bamboos Merostachys clausseni and Chusquea ramossisima . The individual CNP 895 was collected ca. 2 km of the type locality. The rainforest in this area is highly disturbed by human activities, including wood extraction and timber plantations. In fact, the small provincial park “Ernesto Che Guevara” (18 ha), where the specimen was trapped, constitutes one of the last patches of forests in this area. The mean height of the trees in this place is 10-12 m, with a discontinuous dosel. The vegetal community is dominated by the Lauraceae Ocotea spp. and Nectandra spp. and other tree species like Inga sp., Luehea divaricada , Fagara hyemalis , Peltophorum dubium , and Balfourodendron riedelianum . Vegetation in the underbrush includes dense communities of bamboo Chusquea ramosissima . Finally, the individual MLP 1.I. 03.24 was caught in a riverine forest of Ocotea acutifolia and Nectandra megapotamica . The mean hight of the trees in this place is ca. 20 m, with a discontinuous dosel. Other tree species present there include Luehea divaricada , Chrysophyllum marginatum , Balfourodendron riedelianum , Bastardiopsis densiflora , Inga affinis and Peltophorum dubium . At this place, the specimen was caught in dense underbrush of the bamboo Merostachys clausseni .
Genetic variation. Only five cytochrome- b sequences of J. pictipes are available; these specimens were collected in one Argentinean and three Brazilian (one in Minas Gerais and two in São Paulo States) localities. Observed genetic variation among these cyt- b haplotypes is moderate; it ranges form 0.3 to 2.3%. In addition, there is not a pattern of isolation by distance. The most distinct haplotype is the one recovered at the Minas Gerais locality and not the one from the Argentinean locality that is the most apart of the four (see Fig. 1 View FIGURE 1 ). Comparisons involving the haplotype from Minas Gerais and the other J. pictipes haplotypes range from 1.6 to 2.3%, while comparisons between the Argentinean haplotype and the others range from 0.4 to 1.6%. Future studies involving more haplotypes, especially from intermediate localities will test the pattern of genetic structure described here, and explain its biological meaning if any.
Conservation status. D'Elía et al. (2006) categorized J. pictipes in Argentina as data deficient. Due to the lack of primary data about J. pictipes natural history and the fragmentary knowledge about its distribution we prefer to maintain this category for the populations in Misiones Province. Globally, J. pictipes was treated by Baillie (1996) as Lower risk, least concern.
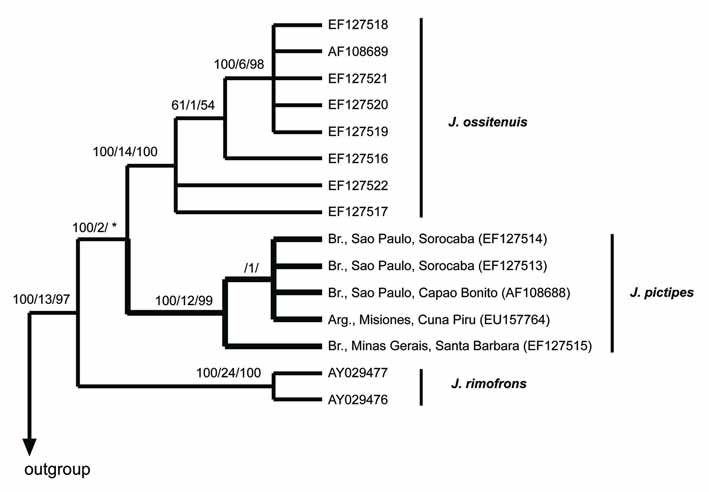
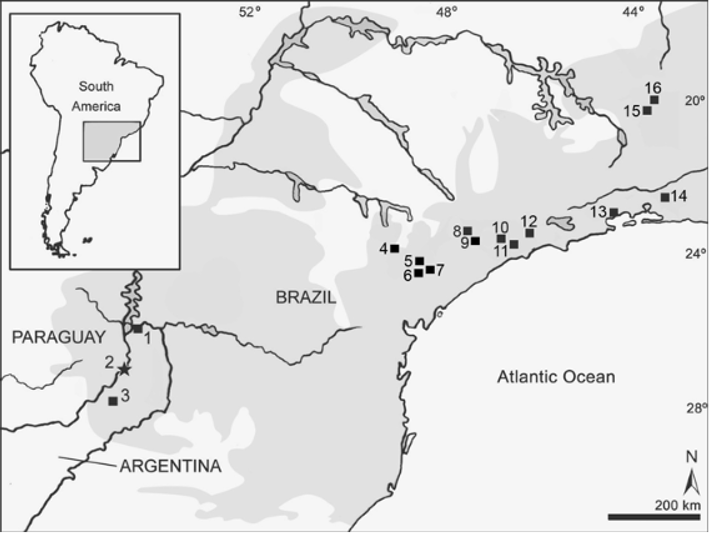
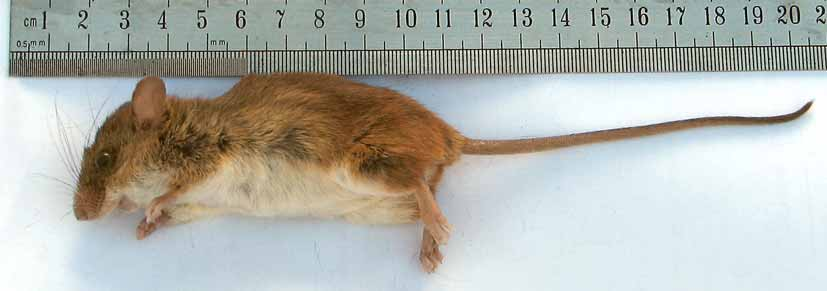
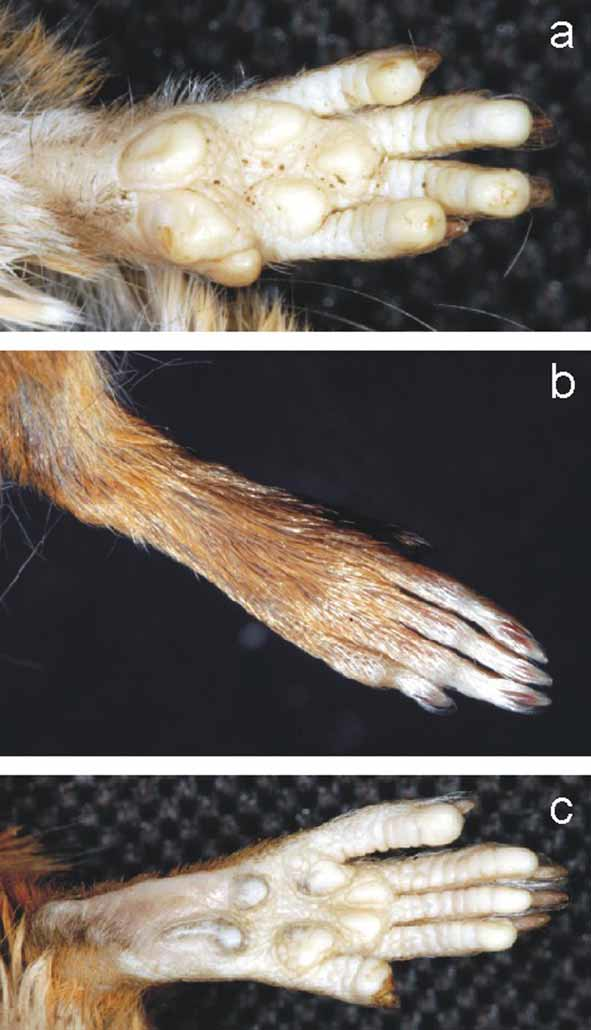
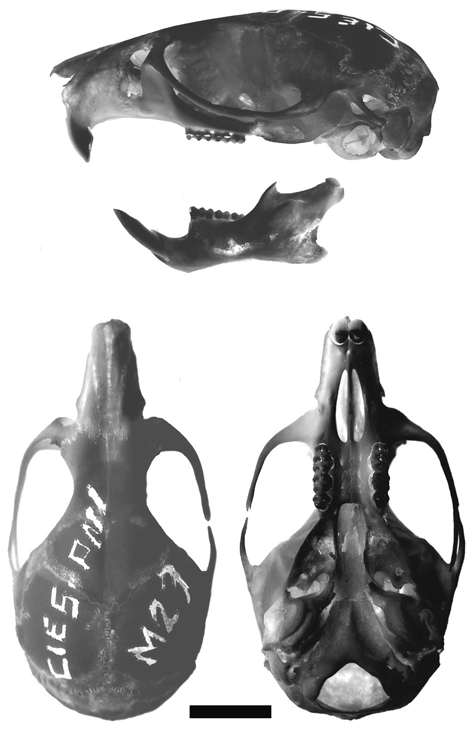
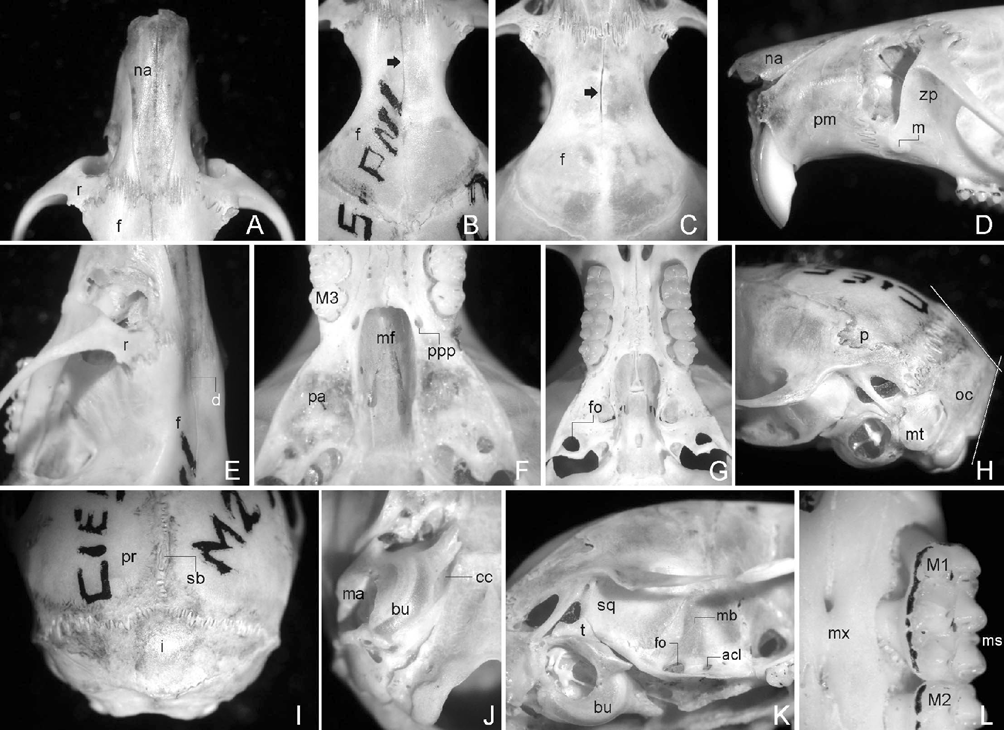
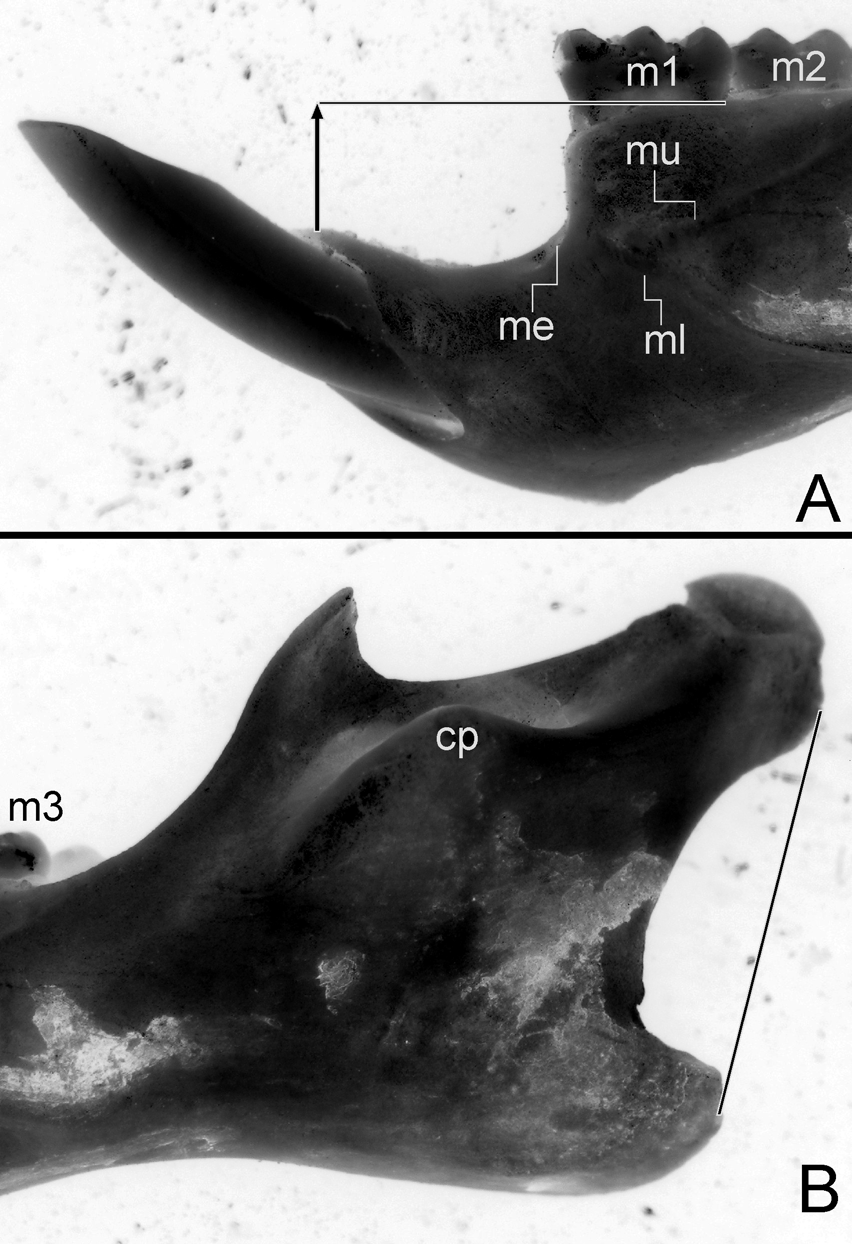
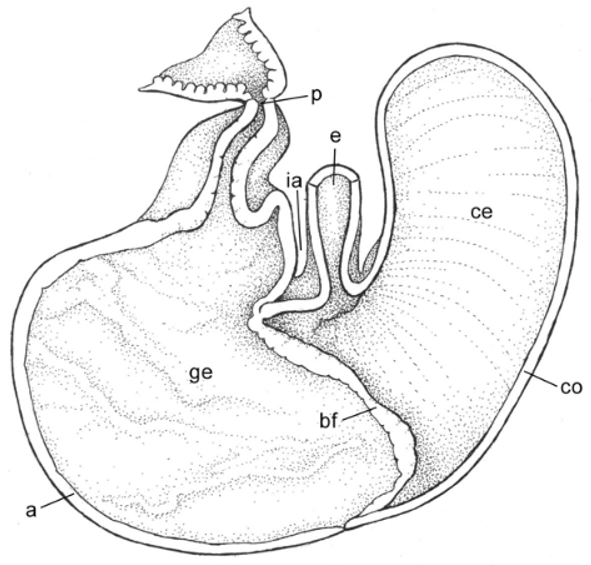
TABLE 2. External and craniodental measurements of Argentinean specimens of Juliomys pictipes. Age classes were defined according to the criteria of Costa et al. (2007).
| MLP 1.I.03.24 | CIES-M 23 | CNP 895 | FMNH 26814a | |
|---|---|---|---|---|
| Age class | 3 | 3 | 1 | 3 |
| Total length | 205 | 185 | 154 | 197 |
| Tail length | 95 | 95 | 82 | 97 |
| Hind foot without/with claw | 18/20 | - | 18/19.5 | -/21 |
| Ear length | 15 | - | 13 | 12 (dry) |
| Weigth (in grams) | - | 22 | 8.5 | - |
| Occipto-nasal length | - | 24.82 | - | 26.30 |
| Palatal length | - | 9.34 | 8.34 | - |
| Postpalatal length | - | 9.36 | 7.91 | 9.00 |
| Molar row–crown length | - | 3.92 | 3.74 | 3.90 |
| First molar breadth | - | 1.08 | 1.06 | - |
| Palatal bridge length | - | 3.82 | 3.74 | - |
| Temporal fossa length | - | 7.77 | 6.90 | - |
| Diastema length | - | 6.03 | 5.07 | 6.50 |
| Incisive foramen length | - | 4.43 | 3.65 | 4.30 |
| Incisive foramen breadth | - | 1.76 | 1.67 | - |
| Palatal breadth at first molar | - | 2.43 | 1.83 | - |
| Palatal breadth at third molar | - | 2.50 | 2.38 | - |
| Mesopterygoid fossa breadth | - | 1.75 | 1.69 | - |
| Breadth across incisor tips | - | 1.66 | 1.51 | - |
| Bullar width | - | 3.28 | 3.23 | - |
| Bullar length | - | 4.11 | 3.26 | - |
| Braincase breadth | - | 11.88 | 10.64 | 12.30 |
| Skull height | - | 7.86 | 6.97 | - |
| Rostral height | - | 4.68 | 4.26 | - |
| Rostral breadth | - | 4.15 | 3.87 | - |
| Rostral length | - | 6.86 | - | - |
| Nasal length | - | 8.04 | - | 9.00 |
| Zygomatic plate length | - | 2.29 | 1.71 | 2.50 |
| Zygomatic breadth | - | 13.23 | - | 14.40 |
| Interorbital breadth | - | 4.19 | 3.76 | 4.50 |
| Greatest length of mandible | - | 12.63 | 11.47 | - |
| Mandibular molar row–alveolar length | - | 4.02 | 3.90 | - |
| Depth of ramus | - | 3.08 | 3.00 | - |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.