
Setosella spiralis Silén, 1942
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4728.4.1 |
|
publication LSID |
lsid:zoobank.org:pub:3E4C6C25-3630-4842-A776-F87CF2E693FD |
|
DOI |
https://doi.org/10.5281/zenodo.5586289 |
|
persistent identifier |
https://treatment.plazi.org/id/0C3887F8-FF8E-FFFE-FF72-FF61EF417ACE |
|
treatment provided by |
Plazi |
|
scientific name |
Setosella spiralis Silén, 1942 |
| status |
|
Setosella spiralis Silén, 1942 View in CoL
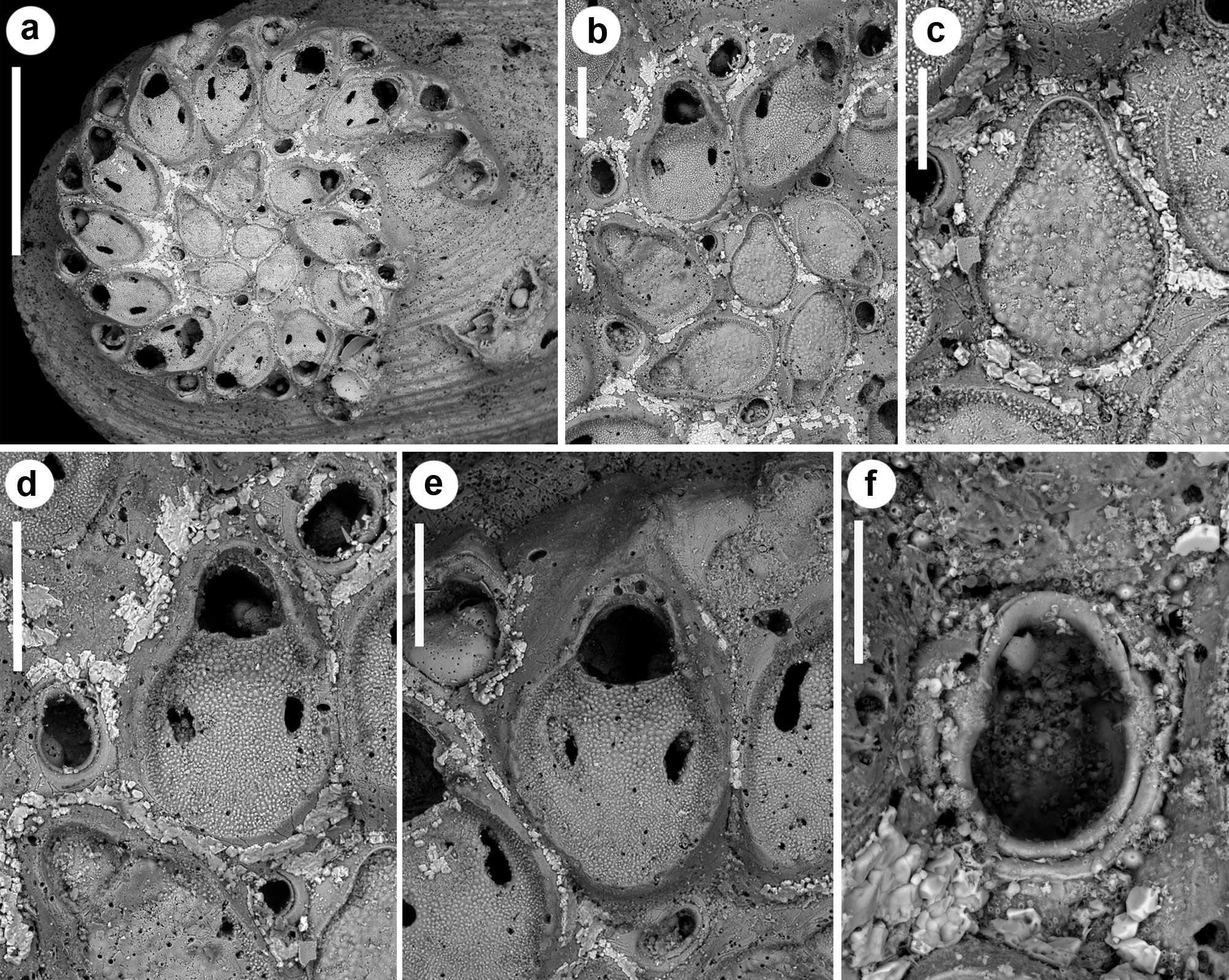
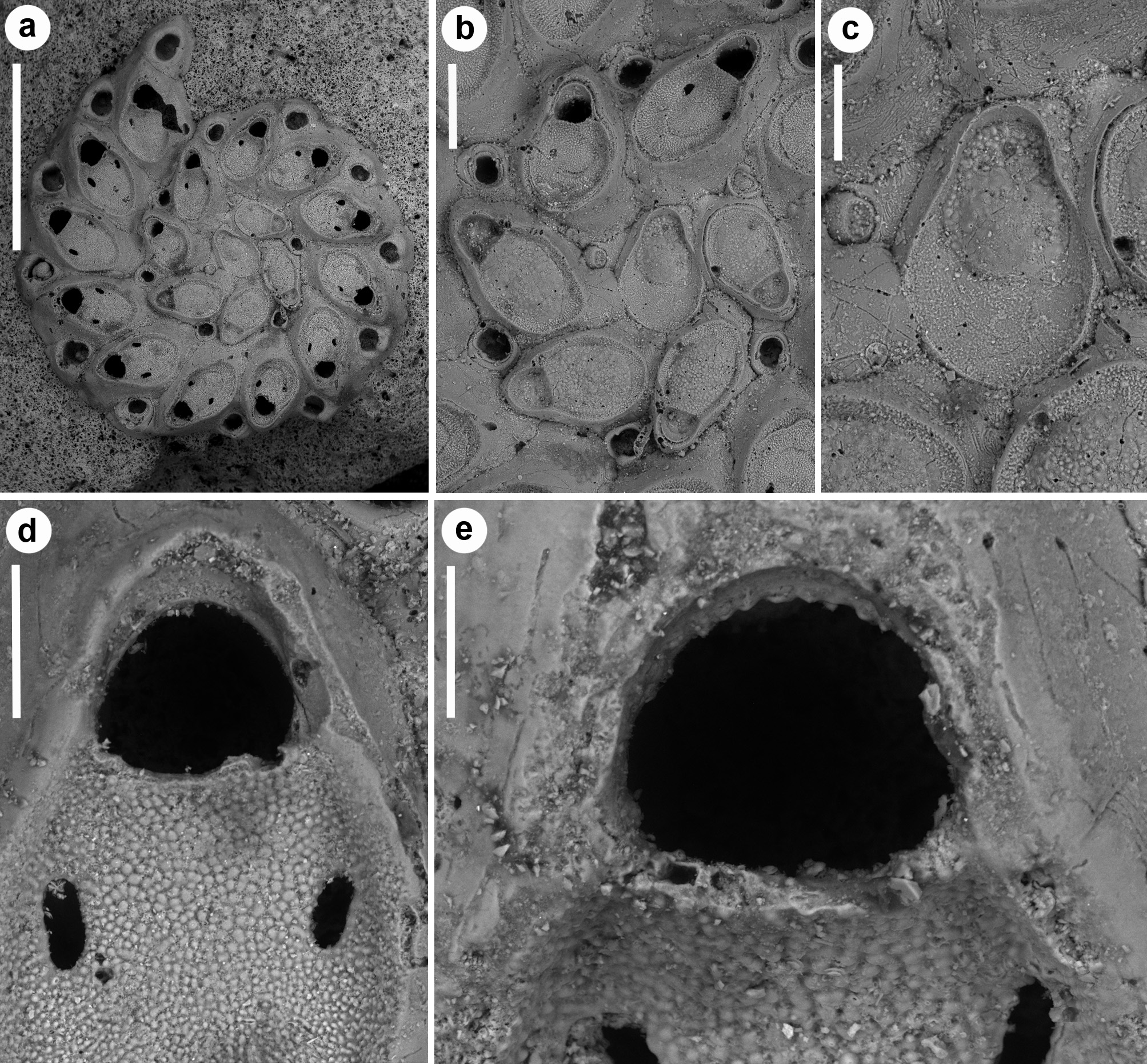
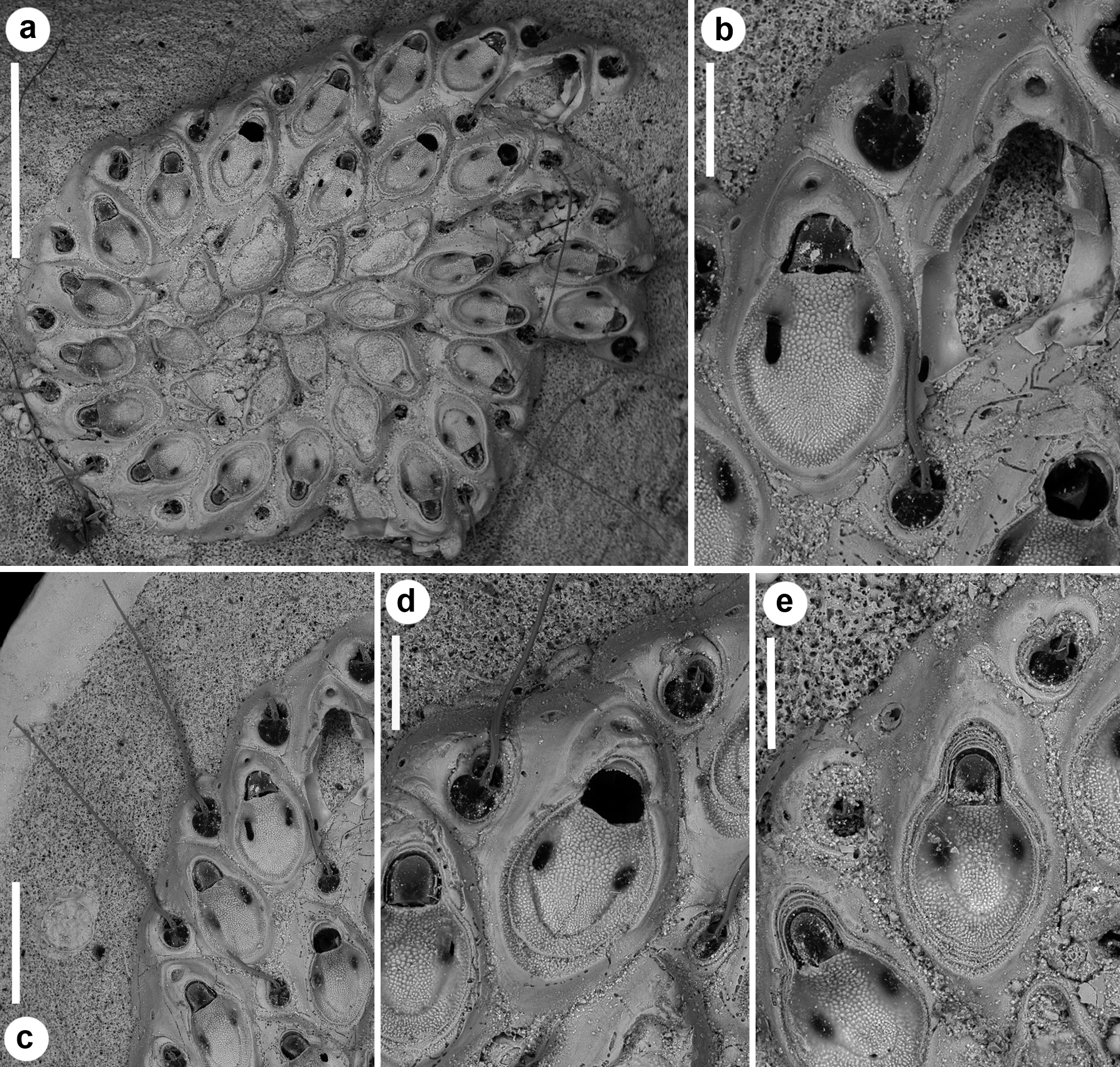
( Figs 15–17 View FIGURE 15 View FIGURE 16 View FIGURE 17 ; Tables 1 View TABLE 1 , 2 View TABLE 2 , 5 View TABLE 5 )
Setosella spiralis Silén, 1942: p. 4 View in CoL , fig. 3; pl. 2, fig. 7.
Examined material. Holotype SMNH-Type-1893, North Atlantic Ocean , Josephine Bank, 36°45,5’N, 14°12,2’W, 340–430 fathoms (c. 500–650 m), Josephine Expedition 1869, a dead colony on the external surface of a bivalve GoogleMaps . Paratypes, same collection number and details as the holotype: (1) five dead colonies, two on internal surfaces of bivalve shells, one on an unidentified bioclast, and two on a 4x 2.5x 2.5 cm biogenic concretion; (2) a live colony on a bioclast.
Description. Colony small, including c. 20 zooids, exceptionally 37 in the largest available colony, entirely adhering to the substratum, consisting of spirally arranged zooids typically in a single right-coiled row and tightly adherent whorls giving a very solid appearance ( Figs 15a View FIGURE 15 , 16a View FIGURE 16 ). Each autozooid budding a disto-lateral daughter zooid from an elliptical septulum (30–45 µm long) located at about mid-length on the right side ( Figs 15a, b View FIGURE 15 , 16a, b View FIGURE 16 , 17 View FIGURE 17 ). A smaller, comparably more distally located subcircular pore (15–20 µm wide) present on its left side, seemingly inactive at the periphery of the colony, but functional to connect zooids in two subsequent whorls. New budded zooids variable in position in relation to the parental zooid, from disto-lateral at an angle of c. 45° with the longitudinal axis of the parental zooid, to mid-lateral or even proximo-lateral and subparallel (or sometimes orthogonal) with the longitudinal axis of the parental zooid. In one of the paratypes, the second periancestrular zooid contemporaneously buds three zooids (one distally and two disto-laterally) developing a ‘triple’ spiral ( Fig. 17 View FIGURE 17 ). Autozooids thick in lateral view ( Figs 15e View FIGURE 15 , 16d View FIGURE 16 ), irregularly elongate and asymmetrical in frontal view, with a cuneiform proximal end and a straight oblique termination on the left side contributing to the continuous, regularly curved colony edge. Lateral gymnocyst contributing to zooidal asymmetry, extensively exposed and gently sloping proximally only on the right side, except in zooids of the peripheral whorl, where it is largely exposed and steeply sloping towards the base also on the disto-lateral left side ( Fig. 16a, b View FIGURE 16 ). Frontal area ovoidal to rhomboidal, bordered by a raised, thin and smooth rim. Cryptocyst extensive, slightly depressed and relatively flat, imperforate except for two opesiules, with variably inclined lateral and proximal edges, gently raising distally to form the proximal border of the opesia; sculptured with roughly radially aligned, fine granulations in the proximal half and along lateral edges, and coarser granulations in the flat area between the opesiules and the proximal margin of the opesia ( Figs 15d, e View FIGURE 15 , 17b View FIGURE 17 ). Opesia transversely Dshaped, approximately as long as wide, with an arched, deeply-located distal shelf of coarse granulations, occupying about one-third of the opesial width, and a proximal straight to gently convex proximal border with two slightly marked lateral indentations ( Figs 15d, e View FIGURE 15 , 16e View FIGURE 16 ). Opesiules paired ( Figs 15d, e View FIGURE 15 , 16d View FIGURE 16 , 17d View FIGURE 17 ), smooth-sided, subcircular to elongate or irregularly-shaped, placed at about zooidal mid-length in the most depressed area of the cryptocyst, at c. 45 µm from the opesia. Vibraculum subglobular to subtriangular in frontal view, budded at the very disto-lateral end of each autozooid, highly inclined to orthogonal, on the right side of the associated zooid and often coaxial to the next autozooid in the row ( Figs 15d View FIGURE 15 , 17b View FIGURE 17 ); cystid visible only along the colony margin, swollen, somewhat asymmetrical with a protruding gymnocystal bulge on the left side; opesia acorn-shaped with blunt condyles separating the subelliptical talon from the short, slightly flared rostrum with a median shallow furrow ( Fig. 15f View FIGURE 15 ); mandibular seta slender, more than twice as long as an autozooid, directed radially outside the colony in most zooids ( Fig. 17c, d View FIGURE 17 ); circular (c. 14–20 µm in diameter), uniporous septula located on the distolateral corners and on the proximo-lateral left corner, visible on the colony periphery. Ovicells ( Fig. 17 View FIGURE 17 b–d) only observed in some zooids of the peripheral whorl, as a gently swollen prominence distal to the opesia; ectooecium smooth defined by a marked flam-shaped margin and with an asymmetrical, subcircular window (40–80 µm wide, mean 65 μm) pierced by a median pore. Terminal, subimmersed ovicell cleithral, with distinct ooecium; the proximal border of the ooecium deeply arched with lateral wings joining the mural rim almost at opesial corners, producing dimorphic opesiae with lateral indentations in ovicellate zooids. Ancestrula ( Figs 15b, c View FIGURE 15 , 16b, c View FIGURE 16 ) oval, nearly as large as the first budded zooids, but decidedly smaller than later autozooids; a narrow rim of gymnocyst exposed distally; cryptocyst gently sloping towards the opesia, smooth, occupying less than half of the proximal frontal area, tapering disto-laterally and disappearing distally; opesia nearly eight-shaped owing to symmetrical indentations of the mural rim, the proximal lobe wider. Opesia, or sometimes the whole frontal surface of the ancestrula, occluded by a coarsely granular plate. Ancestrula budding a single zooid with reversed polarity disto-laterally on the right side, and two vibracula, one disto-laterally, caudate, visible beyond the first budded zooid, and the second one nearly mid-laterally on the left side ( Figs 15a, b View FIGURE 15 , 16a, b View FIGURE 16 ). Closure plates sealing the opesia observed only in periancestrular autozooids in small colonies, but also in later autozooids (excluding those in peripheral rows) in larger colonies ( Figs 15b, d View FIGURE 15 , 16b View FIGURE 16 , 17a View FIGURE 17 ).
Remarks. Most of the studied colonies of Setosella spiralis are thick and characterised by a readily apparent spiral arrangement, resulting from the tightly and regular coiling of either a single or multiple, uniserial zooidal rows. A single colony exhibits a three-spiral arrangement. In contrast, there is no sign of the double spiral arrangement described by Silén (1942, p. 5). It is worth of note that the three-spirally coiled colony in the collection ( Fig. 17 View FIGURE 17 ) shows evidences of organic tissues, including frontal membranes, opercula and vibracular setae, suggesting that it was alive when collected. This contrasts with Silén’s (1942, p. 4, 5) statement that none of the colonies he had available was alive or had traces of vibracular setae. However, Silén’s description of an additional spiral, derived from the first spiral and not directly from the ancestrula, corresponds well with present observation ( Fig. 16a View FIGURE 16 ). Also, some ovicells occur in this colony that were not mentioned by Silén (1942). As Silén (1942) reported to have available only five colonies of S. spiralis , all encrusting “small shells and worm tubes”, it is likely that the biogenic concretion with the two additional colonies (including the only live and fertile specimen) included in the paratype series were added later. The occurrence of the large, three-spiral colony on the concretion indicate that colonies tend to grow larger and to last longer if larger and more persistent substrata are available, although they become more irregular, possibly in relation to surface roughness. As already pointed out by Silén (1942), zooidal morphology is generally constant but the size of both zooids and vibracula is highly variable, remarkably and progressively increasing from the periancestrular zone to the growing termination of a whorl. Regenerations are common in both zooids and vibracula, especially in large colonies. Up to two intramural buds were counted in the vibracula ( Figs 15f View FIGURE 15 , 17d View FIGURE 17 ), while up to five intramural buds, as nested rims in the opesia and cryptocyst, were observed in the autozooids ( Fig. 17e View FIGURE 17 ).
Distribution. Setosella spiralis is presently known exclusively from the type locality, the Josephine Bank in the NE Atlantic, from which colonies were collected in the 500–650 m depth range during the Josephine Expedition in 1869. Although the species was never recorded again after its original description ( Silén 1942), the present re-examination of the type material ascertains that both living and dead colonies were collected. Reverter-Gil et al. (2012, p. 166) reported as possibly fitting in the definition of S. spiralis colonies from a nearby locality, the Lion Seamount near Madeira (35°15,4’N, 15°34,6’W), and a comparable depth (630 m). However, the identity of this material previously identified as S. vulnerata , must be ascertained.
Ecology. Setosella spiralis usually encrusts the convex side of small bioclasts (mostly mollusc shells, as the holotype) less than 1 cm-sized, but two colonies were found on the concave surface of a concretion about 3 cm in maximum diameter. Thus, this species seems adapted to the colonisation of small substrata in soft bottoms of the upper bathyal zone.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Setosella spiralis Silén, 1942
| Rosso, A., Martino, E. Di & Gerovasileiou, V. 2020 |
Setosella spiralis Silén, 1942 : p. 4
| Silen 1942: 4 |