
Setosella vulnerata ( Busk, 1860 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4728.4.1 |
|
publication LSID |
lsid:zoobank.org:pub:3E4C6C25-3630-4842-A776-F87CF2E693FD |
|
persistent identifier |
https://treatment.plazi.org/id/0C3887F8-FF95-FFE6-FF72-FC0CEED57FF2 |
|
treatment provided by |
Plazi |
|
scientific name |
Setosella vulnerata ( Busk, 1860 ) |
| status |
|
Setosella vulnerata ( Busk, 1860) View in CoL
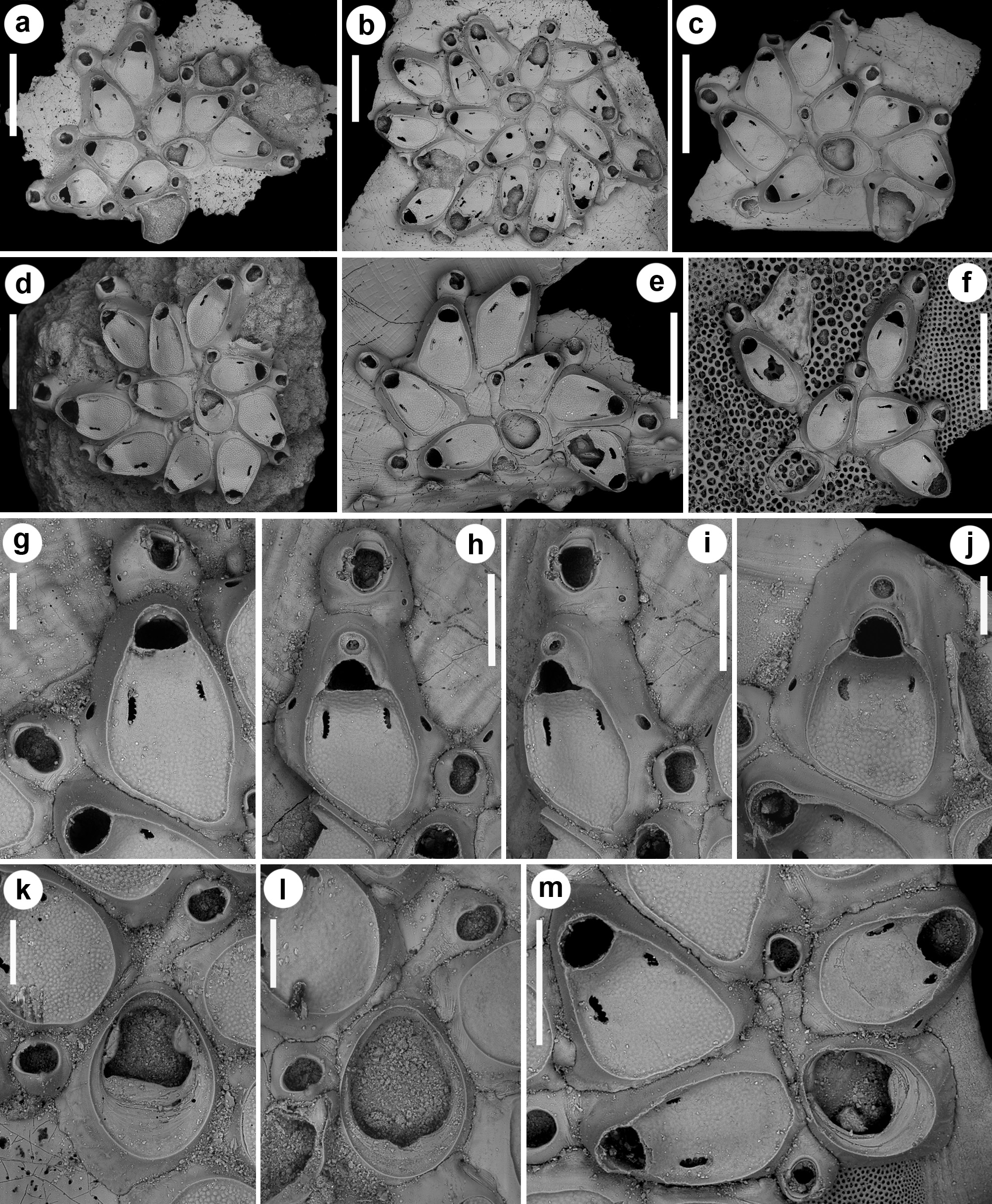
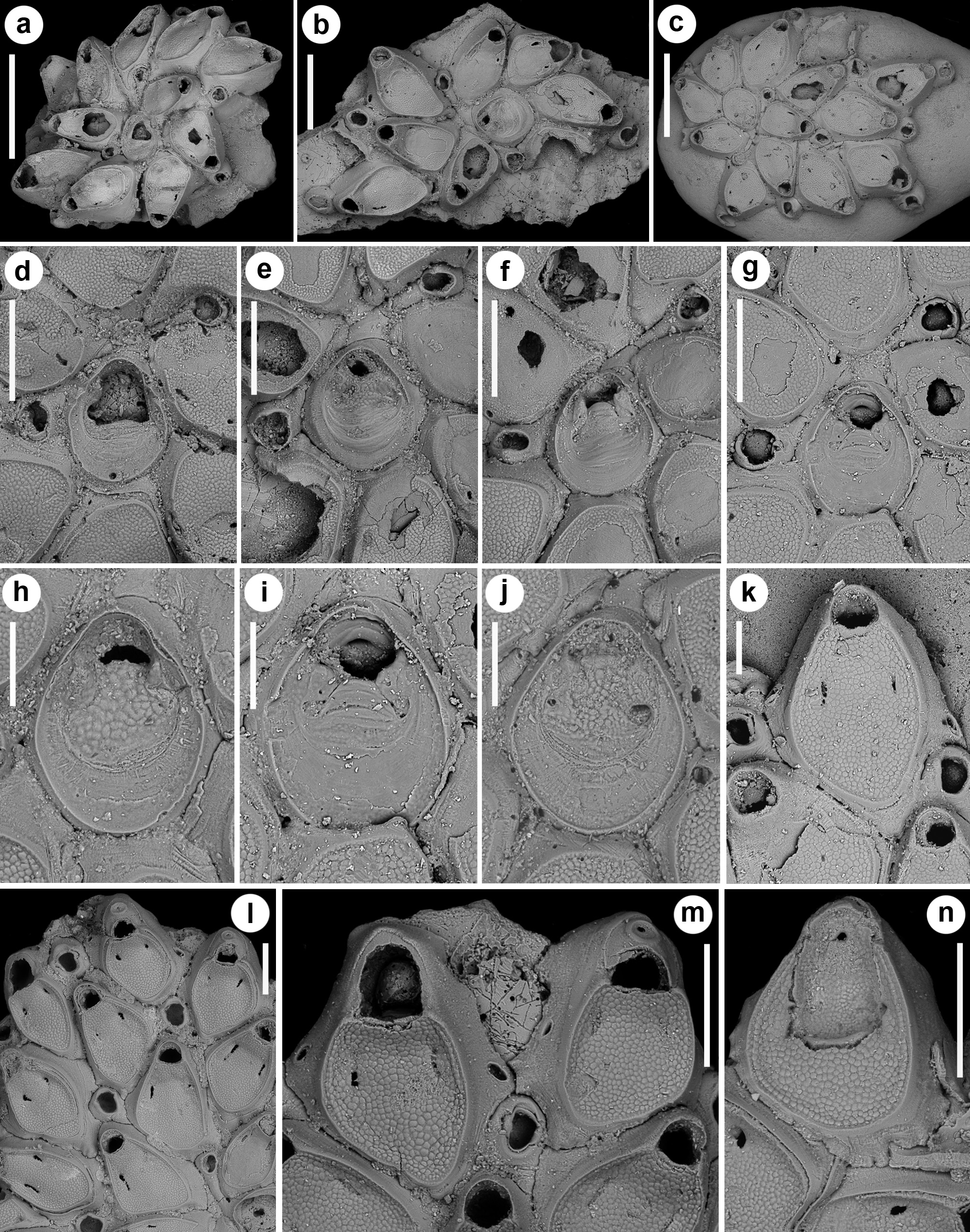
( Figs 1–2 View FIGURE 1 View FIGURE 2 ; Tables 1–3 View TABLE 1 View TABLE 2 View TABLE 3 )
Membranipora vulnerata Busk, 1860: p. 124 . Setosella vulnerata: Hincks, 1880: p. 181 View in CoL , pl. 21, fig. 7; Harmelin & d’Hondt, 1992: p. 28; Di Geronimo et al., 1993: table 3; Di Geronimo et al., 1997: table 3 (pars); Hayward & Ryland, 1998: p. 298, fig. 103; Rosso, 2005, table 3; Mastrototaro et al.,
2010: p. 416; Rosso et al., 2010: p. 364; Souto et al., 2016: p. 416, figs 33–38, table 4 (cum syn.). Setosella sp.: Reverter-Gil et al., 2012: p. 164, fig. 4.
Examined material. SEM micrographs of the lectotype NHMUK 1899.7.1.1487 designated by Souto et al. (2016), provided by B. Berning; PMC. Rosso Collection I.H. B-44a: APLABES cruise, Santa Maria di Leuca, off S Apulia, NE Ionian Sea, Holocene: samples AP1 (seven colonies), AP7 (77 colonies), AP8 (five colonies), AP9 (eight colonies), AP11 (one colony), AP14 (56 colonies), AP15 (72 colonies), AP17 (seven colonies), AP19 (one colony), AP20 (three colonies), AP21 (16 colonies), AP30 (three colonies), depth range 513–749 m; PMC. Rosso Collection I.Ps. B-44b: samples Archi 6 (pars: 73 colonies), Archi 7 (68 colonies), Archi 8 (two colonies), Archi 9 (two colonies), Archi, Calabria, southern Italy, Pleistocene (Mnn19f zone); PMC. Rosso Collection I.Ps. B-44c, Lazzàro, Calabria, Pleistocene: small boulder (two colonies), large boulder (15 colonies), 1993 boulder (seven colonies), sample 723 (16 colonies), ancient quarry (five colonies); PMC. Rosso Collection I.Ps. B-44d, Furnari, Sicily, early Pleistocene: three colonies from two samples; PMC. Rosso Collection I.Ps. B-44e, Würmian to Holocene, Graham Bank, Sicily Strait: CR/90-5 (>20 colonies), CR/90-7 (five colonies), CR/90-12 (>30 colonies), depth range 112–220 m.
Remarks. Reverter-Gil et al. (2012) suggested that the presumed widespread records of Setosella vulnerata might correspond to different species. Souto et al. (2016) fixed the status of the species, re-describing and illustrating S. vulnerata using for the first time SEM images, and selecting the lectotype and paralectotypes from the type locality in the Shetland Islands. Following this revision, present-day and fossil Mediterranean material stored in the collection of one of us (AR), and previously assigned to this species, were re-examined resulting in the identification of a new species, S. cyclopensis n. sp., and numerous specimens falling within the intraspecific variability of S. vulnerata .
Colonies here identified as S. vulnerata were collected from the present-day N Ionian Sea and Pleistocene outcrops in southern Italy. Fossil colonies from Archi resemble the lectotype and paralectotypes in the bi-spiral ar- rangement of periancestrular zooids, becoming less evident later in astogeny, the morphology of zooids and their asymmetrical slit-like opesiules. They also show the two typical caudate vibracula budded from the ancestrula, one short-caudate, budded mid-laterally on the left side, and the other long-caudate, budded distally but strongly curved on the right side ( Fig. 2 View FIGURE 2 d–g). These caudate vibracula are a constant feature and represent the starting point of the two spirals, contributing with the ancestrula to the budding of the first zooids in each spiral row. The tight coiling of the spirals usually produces strongly adjacent zooids but in colonies from Leuca spirals are loose, leaving exposed some portions of the substratum around the ancestrula ( Fig. 1b, c, e View FIGURE 1 ). Occasionally, teratological colonies were observed, in which the short-caudate vibraculum is budded proximo-laterally, leaving its usual mid-lateral budding position to an autozooid and disrupting the normal budding pattern ( Fig. 1f View FIGURE 1 ). Differences between these two groups of specimens and the type material are also visible in the appearance of the cryptocyst, which is tessellated in colonies from Leuca and Archi and granular in the types. However, this difference in the texture of the cryptocyst can be interpreted as falling within the variability of the species, with the coarser granulation observed by Souto et al. (2016) in a colony from the Galicia Bank as a transitional form between them.
The morphology of the ancestrular opesia varies from wide and ovoidal to almost trifoliate, owing to the development of two prominent lateral constrictions. In the fossil specimens, the opesia can be partly to nearly completely closed by cryptocystal plates ( Fig. 2e, h, j View FIGURE 2 ), or by rings of relatively smooth calcification, occasionally forming converging triangular wings from lateral and proximal sides, leaving lateral oblique fissures ( Fig. 2f, i View FIGURE 2 ).
Some colonies also show evidences of breakage and regeneration associated with intramural budding. Zooids sealed by cryptocystal closure plates, with an asymmetrical small pore ( Fig. 2n View FIGURE 2 ) occur in a colony from Archi.
Distribution. Reverter-Gil et al. (2012, as Setosella sp.), Souto et al. (2016), who also revised the material in Harmelin & d’Hondt (1992), and the present study support the Recent North Atlantic and Mediterranean distribution of Setosella vulnerata , although the high morphological variability of the examined material. Mediterranean occurrences relate to the Ionian Sea and the Sicily Strait. However, further records from this area (e.g. Gautier 1962; Hayward 1974; Harmelin 1976; Zabala & Maluquer 1988; Gerovasileiou & Rosso 2016), as well as from the Atlantic and neighbouring seas still need to be checked to assess the real distribution of the species. This also applies to its bathymetric distribution. The new species described here (see below), S. cyclopensis n. sp., based on Mediterranean specimens first identified as S. vulnerata , pertain to shelf bottoms. Colonies reported by De Blauwe (2006, pl. 3, fig. 9; 2009, figs 257, 258) on shells, collected between c. 10 and 25 m depth, in the Kwintebank (North Sea, Belgium) possibly represent a different species. These colonies have smaller autozooids with larger opesiules located closer to the opesia, and vibracula shifted laterally on the right side. It is likely that S. vulnerata will remain restricted to deep habitats from the shelf break and the continental slope. The material from the Sirte Gulf recorded by d’Hondt (1981) probably also belongs to a different species (see Remarks for S. alfioi n. sp.).
The fossil record of S. vulnerata ranges back at least to the MNN 19e biozone, corresponding to the middle-late Calabrian (late early Pleistocene) of Archi. However, it cannot be ruled out that the species appeared earlier owing to the collection of rare colonies at Furnari (NE Sicily) and Lazzàro (Calabria) in sediments referred to a generic early Pleistocene age ( Di Geronimo et al. 2005 and AR, unpublished data). The persistence of the species in the Mediterranean Basin is also indicated by its occurrence in late Würmian-to-very early Holocene submerged sediments in the Ionian Sea and Sicily Strait. The record of S. vulnerata by Moissette et al. (1993, fig. 6a, b) in the late Tortonian of the Heraklion Basin (Crete, Greece) remains questionable on the basis of the figured specimen, although it clearly attests the occurrence of the genus also in the eastern Mediterranean Basin before the Messinian Salinity Crisis.
Ecology. Material studied here was found in silty-sandy sediments. Fresh-looking and alive colonies were collected in the N Ionian Sea at 513–528 m depth ( Mastrototaro et al. 2010), while dead colonies were found down to 749 m, both on surface sediment ( Rosso et al. 2010) and in cored layers dated at about 12,000 years ago ( Malinverno et al. 2010). They were associated with different facies of the Cold-Water Coral habitat and neighbouring coral rubble. Pleistocene colonies originate from comparable palaeoenvironments and depths, as well as from coarse grained facies of the Bathyal Muds Biocoenosis (VP) ( Di Geronimo et al. 1997; Rosso 2005), which are consistent with information reported for the species in the Atlantic Ocean ( Souto et al. 2016). Dead colonies from the Graham Bank in the Sicily Strait, were collected from slightly shallower habitats of the shallow horizons of the continental slope and the shelf break, associated with the VP and the Off-shore Detritic (DL) biocoenoses ( Di Geronimo et al. 1993).
| NHMUK |
Natural History Museum, London |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Setosella vulnerata ( Busk, 1860 )
| Rosso, A., Martino, E. Di & Gerovasileiou, V. 2020 |
Setosella vulnerata
| : Hincks 1880: 181 |
Membranipora vulnerata
| Busk 1860: 124 |