
Setosella cavernicola Harmelin, 1977
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4728.4.1 |
|
publication LSID |
lsid:zoobank.org:pub:3E4C6C25-3630-4842-A776-F87CF2E693FD |
|
persistent identifier |
https://treatment.plazi.org/id/0C3887F8-FF99-FFF6-FF72-F9A8EB7A7FFB |
|
treatment provided by |
Plazi |
|
scientific name |
Setosella cavernicola Harmelin, 1977 |
| status |
|
Setosella cavernicola Harmelin, 1977 View in CoL
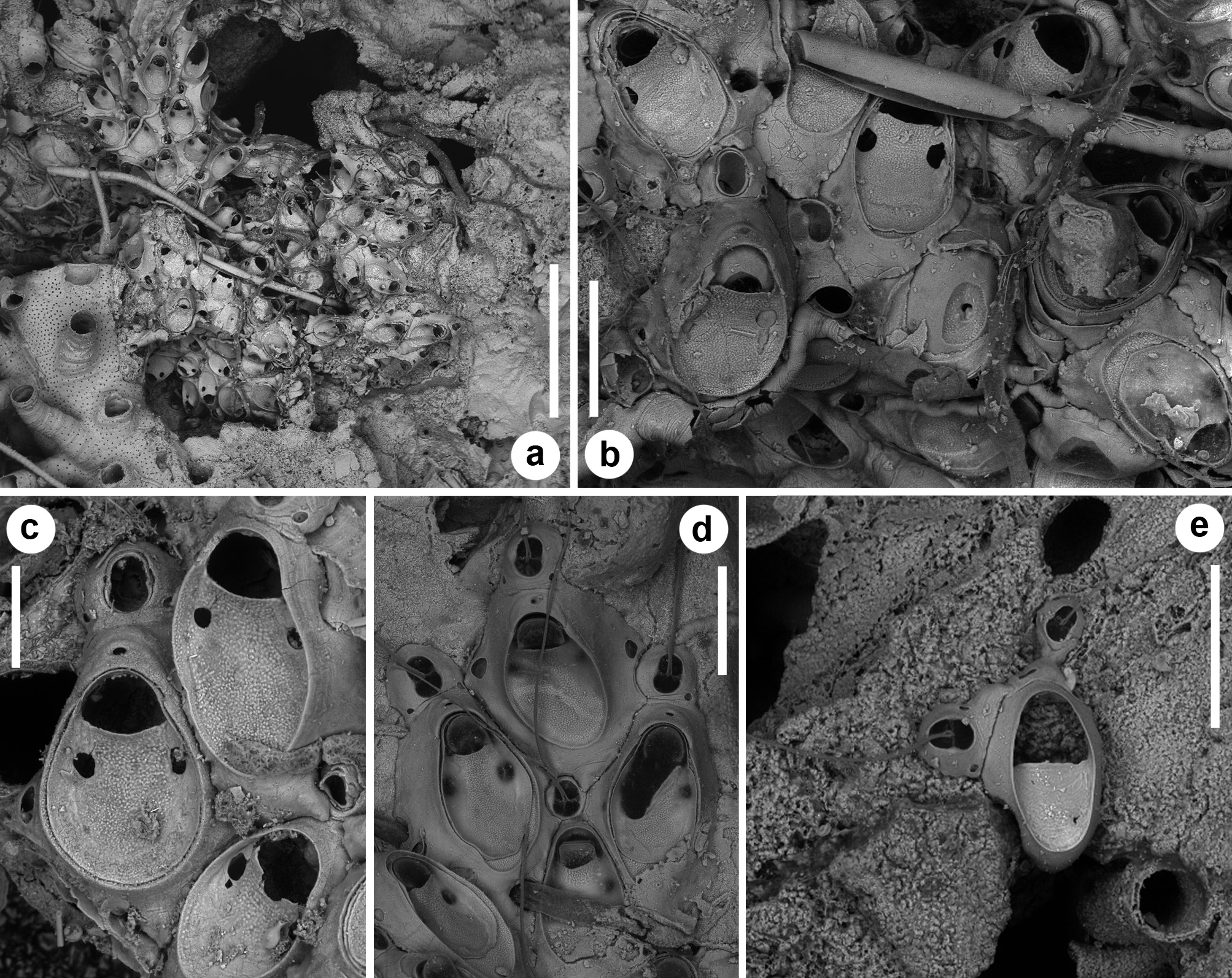
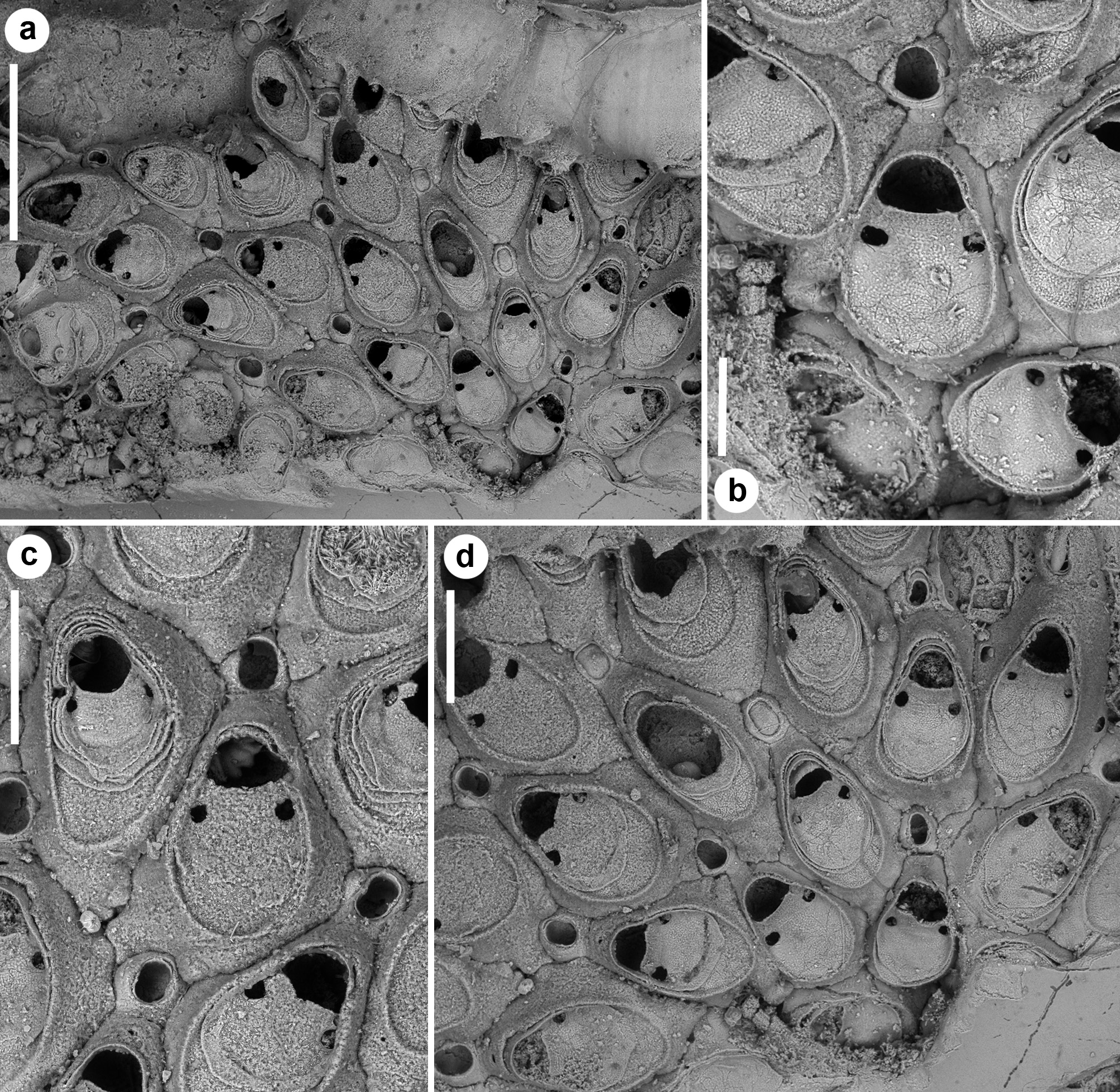
( Figs 8a, b View FIGURE 8 , 9–12 View FIGURE 9 View FIGURE 10 View FIGURE 11 View FIGURE 12 ; Tables 1 View TABLE 1 , 2 View TABLE 2 , 4 View TABLE 4 )
Setosella vulnerata ( Busk, 1860) View in CoL : Harmelin, 1969: table 1 (pars).
Setosella cavernicola Harmelin, 1977: 1064 View in CoL , fig. 17; tav. 1, fig. 7 (pars).
? Setosella cavernicola: Harmelin, 1979 View in CoL (listed); Zabala, 1986: 299, fig. 82c.
Setosella cavernicola: Rosso et al., 2013a View in CoL : table 17.1 (pars); Rosso et al., 2013b: table 1 (pars).
Setosella View in CoL sp. 1: Rosso et al., 2019a: table 1; Rosso et al., 2019b: table 1, fig. 1a.
Examined material. SEM micrographs of the colony MNHN _IB_2008-13068 supposedly designated as the holotype by Harmelin himself when depositing the material at MNHN, and provided by Pierre Lozouet; Rosso Collection. PMC. R.I.H. B-13a. Gymnasium cave, c. 20 m depth, GM2s, c. 35 m from the entrance, four dead colonies each consisting of 3–5 zooids on small biogenic clasts; GM3s, c. 55 m from the entrance, one dead colony of 4 zooids; Mazzere cave c. 20 m depth, MZ 2s, c. 35 m from the entrance, one dead fragment. Granchi cave, GR1P, c. 20 m depth and 10 m from the entrance, seven live colonies on bioeroded concretions, often including the ancestrula and fertile zooids, one of which on the same substratum as the holotype of S. rossanae n. sp. Plemmirio MPA, SE Sicily. Rosso Collection PMC. R.I.H. B-13b. Fara cave, Lesvos Island, 11–18 m depth: FC2, 15– 20 m from the entrance (10 live and six dead colonies); F4, 20– 30 m from the entrance (17 live and nine dead colonies); Agios Vasilios cave, Lesvos Island, 24–40 m depth: VC2, 15– 20 m from the entrance (one dead colony); V 2, 15 – 20 m from the entrance (one live and one dead colony).
Additional material. SEM micrographs of two (out of six) colonies stored at MNHN, catalogue number MNHN _IB_2013-998, as Setosella cavernicola , but belonging to different species, and provided by Pierre Lozouet.
Description. Colony encrusting, unilaminar, forming irregular, uneven patches occupying a few mm 2, usually in the cavities of organogenic concretions ( Figs 9a View FIGURE 9 , 10a View FIGURE 10 , 11a View FIGURE 11 ). Autozooids quincuncially arranged ( Figs 10e, f View FIGURE 10 , 12a, c, d View FIGURE 12 ), elongate rhomboidal, the maximum width reached at about mid-length ( Fig. 12c View FIGURE 12 ). Gymnocyst largely exposed laterally and proximally, gently sloping, outlining a basal surface significantly larger than the frontal one. Proximal end elongate, wedged between proximal zooids, sometimes forming a short cauda-like ( Fig. 10e View FIGURE 10 ). Frontal area ovoidal, marked by a raised, thin and smooth rim, often accounting for nearly two-thirds of the zooidal width. Cryptocyst finely granular, occupying nearly three-fourths of the frontal area, depressed and relatively flat, with a steep edge, slightly raising distally to form the slightly concave proximal border of the opesia. Opesia nearly bellshaped ( Figs 9a View FIGURE 9 , 11c View FIGURE 11 , 12b View FIGURE 12 ), slightly wider than long, with a distal shelf formed by 5–8 slightly prominent tubercles situated in the central arched portion, keeping the operculum in closure position. Spines absent. A single pair of large, c. 30–35 µm wide, roundish-to-slightly elongate, smooth-sided, lateral opesiules inserted on the steep margin of the cryptocyst, at about 40 µm from the proximal margin of the opesia ( Figs 10d View FIGURE 10 , 11c View FIGURE 11 , 12b View FIGURE 12 ). One large uniporous septulum, at nearly mid-length, in each lateral zooidal wall. Interzooidal vibracula circular to ovoidal, placed distally to each autozooid ( Figs 9 View FIGURE 9 b–d, 10e, 11c, d, 12b, c), with two small uniporous septula proximo-laterally, later communicating with distolateral autozooids; opesia ovoidal to eight-shaped, laterally marked by two slightly asymmetrical blunt constrictions bearing barely developed, proximally directed indentations; rostrum arched and flared; vibracular seta about twice as long as an autozooid. Ovicell produced by the maternal autozooid, visible distally to the orifice as a crescent-shaped, low prominence; ectooecium smooth, with a median, circular (less than 20 μm in diameter), infundibular pseudopore ( Figs 9b, d View FIGURE 9 , 10e, f View FIGURE 10 , 11c View FIGURE 11 ). Ancestrula oval, smaller than autozooids ( Figs 9c View FIGURE 9 , 10b View FIGURE 10 , 11e View FIGURE 11 ); cryptocyst smooth with faint growth lines, occupying the proximal half of the frontal area and leaving a wide semi-elliptical opesia; typically buddying a hourglass-shaped kenozooid distally ( Fig. 11e View FIGURE 11 ) budding in turn a small vibraculum distally, and a large vibraculum disto-laterally on the left side of the ancestrula, orthogonally oriented. Occlusion plates observed in some vibracula ( Fig. 11a, d View FIGURE 11 ).
Remarks. Setosella cavernicola was introduced by Harmelin (1977) to distinguish colonies from Trémies submarine cave (near Cassis, Marseille, France) previously attributed to S. vulnerata ( Harmelin 1969) from true colonies of this species collected at Concepcion Bank (Canary Islands). Harmelin (1977, fig. 17) distinguished the new species based on the smaller colony size, the slightly granular cryptocyst and the rounded morphology of the opesiules. These diagnostic characters can also be observed in the colony deposited at MNHN, catalogue number MNHN_IB_2008-13068 ( Fig. 8a, b View FIGURE 8 ), and indicated as the holotype in the MNHN web collection database (accessible at https://science.mnhn.fr/institution/mnhn/collection/ib/item/2008-13068?listIndex=2&listCount=37; accessed 19.11.2019), although Harmelin (1977) does not provide any information on the type material.
In addition, Harmelin (1977, fig. 16) illustrated a group of zooids with four opesiules, stating that the number of opesiules can double in some colonies. The two illustrations also differ in the position of the vibraculum with respect to the autozooid, placed distolaterally in fig. 16 and distally in fig. 17. Four opesiules and disto-laterally placed vibraculum characterise one of the colony ( Fig. 8c, d View FIGURE 8 ), out of six, deposited at the MNHN, catalogue number MNHN_IB_2013-998 (accessible at https://science.mnhn.fr/institution/mnhn/collection/ib/item/2013-998?listInde x=3&listCount=37; accessed 19.11.2019). Both the number of opesiules and position of vibracula are constant within colonies in the type series as well as in the material examined for this study. Consequently, we separate the two species: S. cavernicola Harmelin, 1977 with holotype MNHN_IB_2008-13068 ( Fig. 8a, b View FIGURE 8 ) and we introduce a new species, S. rossanae n. sp. (described below), to which we attribute the colony in MNHN_IB_2013-998 figured here ( Fig. 8c, d View FIGURE 8 ). Zabala (1986, fig. 82A) illustrated the co-occurrence of two, four and five equidimensional opesiules in different autozooids in the same colony with inconstant vibracula always placed distolaterally to autozooids, which it is likely to be a mixture of characters observed in several different colonies as also proved for other bryozoan species (e.g. Rosso 1999).
Setosella cavernicola appears well characterised, and easily distinguishable from all congeners, based on its two, large-sized, circular opesiules and distal vibraculum coaxial with the associated proximal autozooid, and narrow ooecium lacking a window but with a small frontal pseudopore. Setosella folini also has a distally placed vibraculum coaxial with the associated proximal autozooid but differs in having free-living, ring-shaped colonies and slit-like opesiules.
Setosella cavernicola autozooids are variable in size and shape.Although the size of autozooids rapidly increases in the zone of astogenetic change like in other congeners, the size of vibracula appears to be constant. Autozooidal shape seems to change remarkably in S. cavernicola probably because it encrusts particularly uneven surfaces in cavities of concretional walls and ceilings of submarine caves. In this habitat, S. cavernicola has been found in the inner dark recesses and in the transition zone between dark and semi-dark cave sectors ( Harmelin 1969; Rosso et al. 2019 a, 2019b), where food supply is limited. Colonies reach a relatively large size (2–3 mm in diameter) and often autozooids show evidence of multiple regenerations with intramural buds visible as series of mural/oral rims. Up to seven distal oral rims have been counted, and as many roughly concentric, mural rims of superimposed cryptocystal layers, progressively shifting distally up to the autozooid mid-line ( Fig. 11a, c View FIGURE 11 ). In these repeatedly regenerated autozooids, the opesia becomes smaller and irregularly shaped. Regenerations have also been observed in some ovicells and vibracula, although more rarely. Regenerations also occur following breakage of autozooids at any point along walls. All these features suggest that colonies persist for long time and are able to ‘self-repair’.
Distribution. Setosella cavernicola is presently known mostly from submarine caves. First reported ( Harmelin 1969, 1977) from the dark recesses of the Trémies cave near Marseille (about 6 m depth), it has repeatedly been cited from the same area ( Harmelin 1986, 2000), and new colonies were more recently found in submarine caves of the eastern Mediterranean. Setosella cavernicola is common in the Plemmirio Peninsula, Syracuse, eastern coast of Sicily, W Ionian Sea, and the Lesvos Island, NE Aegean Sea ( Rosso et al. 2019 a, 2019b, as Setosella sp. 1), in all the caves sampled at date (three and two, respectively). However, following the introduction of S. rossanae n. sp. (see below), the occurrence of S. cavernicola and/or S. rossanae n. sp. in submarine caves from the Medes Islands ( Zabala 1986), and in shallow bathyal bottoms (250–750 m depth) from the Sicily Strait ( Harmelin 1979) remains to be ascertained.
Ecology. Setosella cavernicola is typical of submarine caves where it has been found in small cavities of organogenic concretions collected along walls and ceilings (6–40 m depth), in dark recesses and in transitional areas between semi-dark and dark sectors. Its occurrence in shallow bathyal bottoms must be checked, also in order to maintain the species within the ecological contingent of taxa typical of submarine caves and deep-water habitats (group 3 of Rosso et al. 2013b). Harmelin (2000) stated that colonies of S. cavernicola are obligate spots, each consisting of less than 10 zooids, but several colonies in the Plemmirio and Lesvos caves are comparably large.



| MNHN |
Museum National d'Histoire Naturelle |
| R |
Departamento de Geologia, Universidad de Chile |
| MZ |
Museum of the Earth, Polish Academy of Sciences |
| V |
Royal British Columbia Museum - Herbarium |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Setosella cavernicola Harmelin, 1977
| Rosso, A., Martino, E. Di & Gerovasileiou, V. 2020 |
Setosella cavernicola:
| Zabala, M. 1986: 299 |
Setosella cavernicola Harmelin, 1977: 1064
| Harmelin, J. - G. 1977: 1064 |