
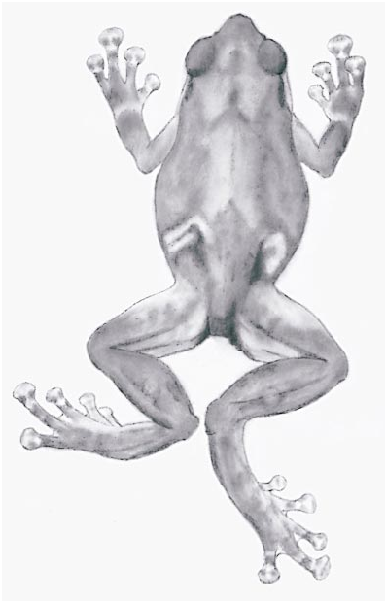
Oreophryne brachypus (Werner)
|
publication ID |
https://doi.org/10.1206/0003-0082(2003)415<0001:SOMFGO>2.0.CO;2 |
|
persistent identifier |
https://treatment.plazi.org/id/0D1FB15D-B912-F71E-4DDF-688963FFE904 |
|
treatment provided by |
Carolina |
|
scientific name |
Oreophryne brachypus (Werner) |
| status |
|
Oreophryne brachypus (Werner) View in CoL
Hylella brachypus Werner, 1898: 554 (type locality, ‘‘Ralum’’ [Gazelle Peninsula, East New Britain Province, Papua New Guinea]; syntypes ZMB 14666 [adult male], ZMB 15483 [ 1 adult male and 1 juvenile], collected by A. Rie on January 17 and February 1, 1897). Werner, 1900: 121.
Hyla brachypus: Barbour, 1912: 177 . Nieden, 1923: 220. Van Kampen, 1923: 37. Loveridge, 1948: 323. Zweifel, 1960: 2.
Oreophryne brachypus: Tyler, 1964a: 4 View in CoL (first use of combination). Tyler, 1967: 187.
DIAGNOSIS: The presence of webbing on the hands distinguishes O. brachypus from mainland species of northern New Guinea. The advertisement call apparently is unlike any other yet described for a New Guinean Oreophryne (see below).
MORPHOLOGY: Snout rounded but almost truncate in dorsal view, loreal region flat or very shallowly concave, nearly vertical, canthus rostralis rounded, nostrils scarcely visible from above and nearly terminal in lateral view. Eyes large, prominent, visible from beneath. Tympanum small, <onethird eye diameter, indistinct at best. Dorsum essentially smooth, only the faintest suggestion of a supratympanic fold. Relative lengths of fingers 3> 2 ± 4> 1, 1st half length of 2nd; fingers with basal webbing, all with enlarged, grooved terminal disks, disk of 3rd finger twice width of penultimate phalange; subarticular elevations low, rounded, inner metacarpal elevation the same (fig. 2B). Relative lengths of toes 4> 5> 3> 2> 1, fifth slightly but distinctly longer than 3rd; all with enlarged, grooved disks, the 1st scarcely broader than penultimate phalange; disks of 3rd finger and 4th toe similar in width, sometimes equal or either may be slightly wider; 5th toe about onehalf webbed, as is 3rd between it and 4th; subarticular elevations scarcely exist, inner metatarsal elevation elongate but barely distinguished (fig. 2B).
Tyler (1964a) described and illustrated the holotype, its pectoral girdle (with ligamentous connection of clavicle to scapula), and the Yshaped terminal phalanges.
COLOR AND PATTERN: ‘‘In life brachypus has a dusky brown ground colouration with indistinct and slightly darker markings on it. Of these markings the most common are a narrow transocular bar and a Wshaped patch immediately behind the head’’ ( Tyler, 1967: 187); ‘‘Upper side brown (in life dark brown or olive green), underside gray, wholly or only throat finely spotted, or white (in life greenish yellow)’’ ( Werner, 1898: 554). Notes accompanying the ZMUC specimens record a grayish to blackish brown dorsal color in life and a pale gray to grayviolet venter. Ventral surfaces of preserved specimens may be sparsely stippled with dark pigment or may have a denser, more mottled appearance.
VARIATION IN SIZE AND PROPORTIONS: Tyler (1967: 187) reported the average SVL of 16 adult males as 19.9 mm (range 19.2–22.3 mm) and of three adult females as 22.0 mm (range 21.5–22.8 mm). In our series of nine specimens the largest male is 22.5 mm and the largest specimen, presumably a female but not sexed, is 24.0 mm. See tables 1 and 2 for proportions.
ADVERTISEMENT CALL: ‘‘The call consisted of a single note, resembled a long squeak and had a duration of about four seconds’’ ( Tyler, 1967: 188). Known calls of other Oreophryne are groups of repeated shorter notes or pulsed single notes shorter than described for brachypus and not likely to be described as a ‘‘squeak’’.
COMPARISONS WITH OTHER SPECIES: Apart from possession of finger webbing, brachypus has average morphology for Oreophryne . The species is most similar to geislerorum and cannot reliably be distinguished from that species through any of the standard ratios or combinations of them. Only the mean EN/SVL ratios differ substantially, but the range of this ratio in geislerorum overlaps that of brachypus completely. Apparently brachypus is a smaller frog than geislerorum , males of which species attain almost 27 mm SVL and females 29 mm. The call of geislerorum , a harsh sounding series of closely spaced pulses lasting about 0.5 sec, evidently is quite different from the ‘‘long squeak’’ of brachypus .
HABITAT AND HABITS: Two of the syntypes were found ‘‘in a nest built of living moss ¼ meter high in a large Zingiberacae [ginger]’’ ( Werner, 1900: 121). Tyler (1967: 183) stated ‘‘The males were located... calling from vegetation at heights of from three to eight feet from the level of the ground.’’ Photographs in Wolff (1966: pl. 32, fig. 2; pl. 33, fig. 1) give a general impression of habitats around Yalom where the ZMUC specimens were taken.
DISTRIBUTION: Limited information indicates a wide distribution in New Britain, at least from the southwest coast to the northeast tip of the island, and from 140 to 1000 m elevation (fig. 9).
LOCALITY RECORDS AND SPECIMENS EXAM INED: PAPUA NEW GUINEA: East New Britain Prov.: Yalom, 1000 m, 35 km SE Cape Lambert, Gazelle Peninsula (ZMUC E385, 401, 411, 431, 434, 437); Gaulim, 140 m (AMNH A84513); Keravat, Gazelle Peninsula ( Tyler, 1967: 187); Talalo, Nakanai Mtns., 900 m (BPBM 1182–1183); Ralum ( type locality). West New Britain Prov.: Pomugu and Aliwoh, both near Kandrian ( Tyler, 1967: 187); Garu (UPNG 4107); Awit River, 05°59'S, 150°41'E (AMS 129629, 129630, not examined); 8 km NNE Amelei Village 06°02'S, 150°39'E (AMS 129618, not examined).
REMARKS: From the time of its description in 1898 up to 1964, this species was considered a hylid. Except for Werner’s (1900) reprise of the original description, however, evidently none of the reports published subsequent to the original description until Tyler (1964a) was based on examination of the syntypes or other specimens.
The morphology of the few specimens of brachypus gives no reason to suspect the presence of more than one species of Oreophryne on New Britain, but we urge future collectors to obtain recordings of calls in or der to test this assumption and, in any event, to provide for a more objective description of the call.
Oreophryne geislerorum (Boettger)
Figure 12 View Fig
Cophixalus geislerorum Boettger, 1892: 24 View in CoL (type locality, ‘‘Kaiserwilhelmsland, Neuguinea’’; holotype SMF 4197, collected by Bruno and Hubert Geisler in 1890 or 1891; see Type Material and Locality).
Sphenophryne biroi: Méhely View in CoL ¨, 1901: 252 (specimens from Sattelberg).
Hylella brachypus: Vogt, 1911: 427 (specimen from Sattelberg).
Oreophryne biroi: Parker, 1934: 170 View in CoL (part, specimens from Sattelberg, Kokoda,and Orrori). Loveridge, 1948: 423 (part, specimen from Gusiko). Menzies, 1976: 62, pl. 12f. Häupl et al., 1994: 34.
Oreophryne geislerorum: Menzies et al., 1980: 233 View in CoL (first use of combination). Iskandar and Colijn, 2000: 52.
Oreophryne sp. : Zweifel, 1980: 411.
TYPE MATERIAL AND TYPE LOCALITY: The holotype is in moderately good condition, though owing to dissection it lacks elements of the pectoral girdle diagnostic of the genus Oreophryne . Additional specimens, part of the same collection (and lacking precise locality data), were sold to at least one other museum (Vienna) where they are cataloged as ‘‘paratypes’’ of geislerorum ( NMW 19828, 3 specimens); Häupl et al., 1994: 19). But Boettger evidently used only the holotype in formulating his description, so the Vienna specimens have no formal status as types, though presumably they are topotypes.
The type locality as given in the original description encompasses about half of mainland Papua New Guinea. Information on the travels of the collectors ( Wichmann, 1912: 517–518) suggests that the holotype was obtained somewhere between the eastern tip of the Huon Peninsula and Bogadjim (= Stephansort), about 300 km to the westnorthwest on Astrolabe Bay. The known distribution of geislerorum does not include localities as far west as Bogadjim. The former German administrative center of Finschhafen, Morobe Province, Papua New Guinea, or the mission station of Sattleberg nearby are more likely candidates for the collection site.
DIAGNOSIS: An Oreophryne of maximum length about 29 mm with ligamentous connection of procoracoid and scapula, fifth toe longer than third, fourth toe webbed to subarticular elevation, head width relatively narrow (mean HW/SVL 0.346), and internarial space relatively broad (mean IN/SVL 0.085). Among the north coast species, geislerorum shares its broad internarial space with only biroi and brachypus . The latter is distinguished by its finger webbing and insular isolation. Shorter tibia length and narrower head width of geislerorum afford an almost complete separation from biroi . The call of geislerorum is unique (see Advertisement Call).
MORPHOLOGY: Head slightly narrower than body, nares barely visible from above, canthus rostralis moderately distinct, loreal region steep, flat; interorbital space about 1.5× eyelid width, corneal outline visible from beneath; tympanic annulus barely distinct; a weak, diagonal postorbitalsupratympanic fold. Fingers and toes with welldeveloped terminal disks (fig. 2D). Widths of disks on third toe and fourth finger closely similar, typically equal or the finger disk slightly broader, less often the toe disk slightly broad er. Relative lengths of fingers 3> 4> 2> 1, tip of first finger reaches just past the subarticular elevation of the second, no webbing, subarticular elevations low, rounded, inner metacarpal elevation elongate, slightly more prominent. Relative lengths of toes 4> 5> 3> 2> 1, basal webbing, subarticular elevations low, rounded, inner metatarsal elevation low, elongate, slightly more prominent.
COLOR AND PATTERN: A specimen from Ko koda (R.G.Z. field notes) had the following coloration in life: Dorsum yellowish brown with a darker brown patch posterior to and between the eyes; a pair of indistinct dark brown lateral bands on the body, the area between the bands and the head spot somewhat reddish brown. Forelimbs reddish brown distally and yellowish proximally, no distinct markings. An indication of a light wrist band set off by a darker graybrown band and a band around the distal part of the forearm. Underparts yellow, chin body and legs included, relieved only where the viscera show through slightly darker in the abdominal region. The same yellow present laterally on the body where it gives way to the yellowbrown of the dorsal surfaces. A slight narrow dark brown canthal stripe continued faintly over the tympanum. Ground color of hind limbs light brown with no definite markings but some faint reddish brown splotches. Iris golden brown, pupil horizontal.
A specimen from Lae (R.G.Z. field notes) had a brown dorsal ground color with illdefined darker markings, most prominent in the scapular region. Anterior to a transverse line drawn through the middle of the eyes the snout was much lighter brown, almost golden. The ventral surfaces from chin to toes were grayish white without markings.
In preservative the specimens remain brown with darker pattern elements but lose the golden or yellow aspects of the living animal.
VARIATION IN SIZE AND PROPORTIONS: The largest of 14 females measures 29.4 mm SVL, the largest of 30 males 26.6 mm. The smallest gravid female is 24.1 mm, and the smallest male with vocal slits 20.6 mm. Proportions are summarized in tables 1 and 2.
ADVERTISEMENT CALL: The call of Oreophryne geislerorum (fig. 13A, table 3) is harsh with rapidly repeated notes that are not individually discrete to the human ear though they are readily resolved on waveform display. The duration is about 0.3–0.5 sec and the dominant frequency 3200–3400 Hz. The frogs call at irregular intervals averaging about 7 sec but ranging from 2 to 20 seconds in a sample of 65 calls of five individuals. Notes per call range from 18 to 40 in a sample of 64 calls of seven individuals, but any one frog shows much less variation (table 3). The note rate changes within each call, typ ically starting rapidly, then slowing down for the most of the call, and speeding up toward the end. Some individuals have a slower initial rate and fewer notes per call so that rates averaged over the entire call vary greatly, even at closely similar temperatures.
The call of O. geislerorum is quite distinctive and different from that of any other Oreophryne known to us. However, it is peculiar in the wide variation of note rate, about 50–135 notes per sec (table 3), while notes per call and call duration remain relatively constant. Different rates may have been associated with different (unobserved) behaviors. Differences in body temperatures could be responsible. Unfortunately, temperature was not noted by the recordists of the most divergent calls. Variation in tape speed in the equipment used to record and copy tapes could cause spurious rates, but cannot be the full explanation. (The fastest rate and a more typical one were recorded on the same machine a few days apart.) Another possibility is that geislerorum as we define it consists of two or more sibling species, but information available at present is insufficient to explore this option.
COMPARISONS WITH OTHER SPECIES: Oreophryne geislerorum is not known to be sympatric with any other Oreophryne . Any specimens from the gap between the known range of geislerorum on the one hand and the ranges of biroi and hypsiops on the other (figs. 9, 14) should be readily identified (see Key). If O. geislerorum ranges farther to the southeast than is now known, it could contact several species of Oreophryne occurring in the Milne Bay area that are outside the scope of this study. These include the insular species O. inornata and O. insulana and O. loriae . Compared to geislerorum , inornata is a much larger species, up to about 42 mm SVL, and insulana is broader headed (mean HW/SVL 0.393 vs. 0.346). Menzies (1976: 62) recorded O. loriae from Milne Bay at the southeastern tip of the Papua New Guinea mainland. This species (based on examination of the type series) has much in common morphologically with geislerorum and perhaps may best be distinguished from geislerorum by its advertisement call, ‘‘a series of 6 to 12 short, rather nasal, buzzes, uttered at a rate of 2 per second’’ ( Menzies, 1976: 62).
HABITAT AND HABITS: Stocks (1987: 18) published an account of this species, from which we quote: ‘‘ Oreophryne sp. ... can be found in shrubs and trees at night, is common in suburban gardens in Lae as well as the tropical rainforest.... It frequently calls from banana leaves and lays its eggs inside the stalk of a banana leaf near the main stem.’’ Stocks observed a captive female approach a male calling on a banana leaf and follow him as he moved down inside the stalk. ‘‘The resulting eggs are laid in a sticky string which adheres to the stem of the banana leaf. The male remains with the eggs, mainly to drive off insects... and often can be found with two strings of eggs at different stages of development.’’ An excellent photograph (by Mike McCoy) illustrates a frog with an egg clutch.
Zweifel (1980: 411) found these frogs on the Huon Peninsula ‘‘hiding during daytime in banana plants and in cut bamboo stems in gardens and in second growth [at 1340 m].’’ He reported frogs calling ‘‘from sites 4 to 8 feet up in banana and pandanus plants’’, but ‘‘neither found nor heard this species at Gang Creek [ 1340 m] in the tall primary forest.’’ Calling sites include both under and upper surfaces of broad leaves (banana and ginger), sometimes ‘‘too high in the trees to be approached’’ (R.G.Z., field notes).
DISTRIBUTION: This species ranges from the tip of the Huon Peninsula westward at least to Lae and the Wampit River and from there southeast at least as far as Kokoda and Popondetta, Northern Province. The extent of the range inland from Lae is unknown, but neither geislerorum nor any other Oreophryne has yet been taken at Wau ( 1100 m), despite much collecting in the area. Records for Northern Province (verified by recordings of the call) are disjunct (fig. 14), but the Papuan coastal region north of Popondetta has been poorly collected for frogs, and the distribution may be continuous through this region. The range in elevation is from virtually sea level at Lae to 1340 m on the Huon Peninsula. See Remarks.
LOCALITY RECORDS AND SPECIMENS EXAM INED: PAPUA NEW GUINEA: German New Guinea ( SMF 4197 About SMF , holotype) ; New Guinea ( NMW 19828, 3 specimens) . Morobe Prov: Huon Peninsula, Sattelberg ( Méhely ¨, 1901: 252; Parker, 1934: 170; NMH 19825) ; Huon Peninsula, Mogisang , 550 m ( UPNG 5710 , 5711 ) ; Huon Peninsula, Manga, Go River , 600 m ( UPNG 5725 , 5726 ) ; Huon Peninsula, Dregerhafen ( UPNG 2989 , 2990 ) ; Huon Peninsula, Gusiko ( USNM 119199 About USNM ) ; Huon Peninsula, Tumnang , 1340 m ( AMNH A58374 , 75026–75030 , 75032 , 75034–75040 , 145570– 145582 , MCZ A 28402, 28403 About MCZ ) ; Huon Peninsula, Pindiu , 820–910 m ( AMNH A76025– 76030 , BPBM 1178 About BPBM ) ; Huon Peninsula, Mindik ( BPBM 5285 About BPBM , 5286 About BPBM ) ; Gurakor , 640 m ( AMNH A66992 ) ; Lae ( AMNH A75041–75044 , 81195 ; UPNG 3631–3634 , 3715–3717 , 4098 , 4099 , 4138 , 4394 , 4906 , 4908 ) ; 4 km N Lae ( MCZ A92446–92449 ) ; S of Buso River ( BMNH 1980.669 ) ; upper Mo River, Bowutu Mtns (call record only, no specimen). Northern Prov : Kokoda, 370 m ( AMNH A75045 , 75046 , BMNH 1934.1.5.7–9) ; ca. 1 km N, 9 km W Popondetta , 300 m ( UPNG 3964 , 5473 ) ; Orrori , 910 m, 16 Å km SW Kokoda ( BMNH 1934.1.5.10) .
DISCUSSION: From the time of its description in 1892 up to 1980, geislerorum appeared in the literature as a species of Cophixalus of questionable provenance, and at least one author questioned the generic assignment (van Kampen, 1919: 54, ‘‘= Oreophryne sp. ?’’; van Kampen, 1923: 146, ‘‘Probably... belongs to Oreophryne or Sphenophryne ’’). Menzies et al. (1980) identified geislerorum as an Oreophryne and recognized the name as the most senior among all New Guinean Oreophryne . These authors, however, did not determine whether geislerorum was a senior synonym of a species then recognized and did not associate it with a natural population.
Our determination that the name geislerorum is properly identified with the species here characterized derives both from morphological considerations and from geography. As noted under Type Material and Type Locality, although the place of collection cannot be precisely inferred, it can be bound ed more closely than the more generalized published type locality. In the region thus circumscribed, and in Morobe Province as a whole so far as we know, only this one species of Oreophryne occurs.
We have calculated and plotted regression lines for several measurements in a sample of 48 specimens of geislerorum . In all measurements, the holotype of geislerorum falls within the ranges seen in this series, in most cases close to the regression lines, even on them in two instances. The body size is well centered in the distribution, and relative sizes of finger and toe disks, relative toe lengths, and amount of toe webbing also agree.
REMARKS: The status of Oreophryne occurring southeast of Popondetta to Milne Bay remains to be elucidated. Menzies (1976: 62) recorded O. loriae at Milne Bay, but several unidentifiable species of Oreophryne are present as well. Specimens from Mt. Dayman recorded by Zweifel (1956) as O. biroi are neither that species nor O. geislerorum and probably not O. loriae .
Oreophryne hypsiops , new species
Figure 15 View Fig
HOLOTYPE: AMNH A83044 (field tag RZ
8041) from the vicinity of Sempi, 9 km north
of Alexishafen , sealevel elevation, Madang Province, Papua New Guinea, collected by R.G. Zweifel on July 13, 1969 .
PARATYPES (all from Papua New Guinea): Madang Prov.: UPNG 9474 , Yilu Village , 25 km N, 46 km W Simbai, 500 m, collected by J. Menzies ; AMNH A83043 , 83045 , vicinity of Sempi, 9 km N Alexishafen , collected by R. Zweifel , July 12–13, 1969; UPNG 7354 , Nobanob , 7 km N, 5 km W Madang, collected by J. Menzies ; UPNG 7030 , Bauman Village , 27 km S, 13 km W Madang, collected by J. Menzies ; AMNH A 83046 , 83047 , Adelbert Mtns., Wanuma, 975 m, 35 km N, 54 km W Madang , collected by R. Zweifel , August 5–6, 1969. East Sepik Prov.: BPBM 1171 About BPBM , Dreikikir , 350–400 m, collected by M. and J. Gressitt , June 23, 1961; BPBM 1179 About BPBM , Wewak , 2 m, collected by M. and J. Gressitt , June 26, 1961. West Sepik Prov.: AMNH A78141 , Miliom, 3 km E Lumi , 460 m , July 20–23 1966, collected by Jared Diamond.
ETYMOLOGY: The specific name is a noun in apposition, derived from the Greek hypsi (on high) + ops (with included meanings of ‘‘eye’’ and ‘‘voice’’). The allusion is to a presumably watching frog that often vocalizes frustratingly well above the heads of collectors/recordists.
DIAGNOSIS: An Oreophryne with a maximum length of about 26 mm, ligamentous connection of scapula and procoracoid, narrow head (mean HW/SVL 0.343), closely spaced nostrils (mean IN/SVL 0.067), moderate eyenaris distance (mean EN/SVL 0.090), third and fifth toes approximately equal in length, and toes partially webbed, the fourth short of the proximal subarticular elevation. Dorsal and lateral surfaces lacking abundant tiny white spots. This combination of characters together with the distinctive advertisement call will distinguish O. hypsiops from other species in the north coast region of New Guinea.
DESCRIPTION OF HOLOTYPE: Adult male (calling when taken) with the following measurements and proportions: SVL 22.6, HW 7.5, TL 9.9, EY 2.55, EN 1.85, IN 1.6, HD 6.3, FT 9.6, FD 1.7 (penultimate phalanx 0.7), TD 1.6 (0.8); HW/SVL 0.332, TL/SVL 0.438, EY/SVL 0.113, EN/SVL 0.082, IN/ SVL 0.071, HD/SVL, 0.279, FT/SVL 0.425, FD/SVL 0.075, TD/SVL 0.071.
Head somewhat narrower than body; canthus rostralis rounded, not distinct; loreal region flat and nearly vertical; nostrils barely visible from above, internarial span relatively narrow (mean IN/SVL 0.069), shorter than eye–naris distance (mean EN/SVL 0.086); interorbital space 1.5× eyelid width, tympanic annulus barely visible. Relative lengths of fingers 3> 2 = 4> 1, 1, 1st> onehalf 2nd, all with welldeveloped terminal disks, no webbing, low, rounded subarticular elevations (fig. 2B); relative lengths of toes 4> 5 = 3> 2> 1, all with broadened terminal disks, that of 4th toe slightly narrower than that of 3rd finger; low, rounded subarticular elevations, sparse webbing reaching barely to proximal subarticular elevations of toes 1–3 and 5, less on toe 4 (fig. 2B).
In preservative, the dorsal surfaces are brown, darker laterally than in the middorsal region but not differentiated from the color of the top of the snout. There are moderately conspicuous lumbar ocelli. The loreal region has a paler area between the more darkly pigmented snout tip and the area beneath the eye. The groin and proximal anterior part of the thigh are pale and unmarked, the area proximal to the knee is darker. The cloacal region is dark, the rest of the posterior thigh coarsely mottled dark and light. The upper surfaces of the legs are largely brown with some paler markings in no definite pattern; the underside of the thigh is brown, that of the shank largely pale. The chin, throat, chest, and abdomen are brown with numerous paler light flecks. There are no specific notes on the color of the holotype in life except for its similarity to others captured nearby (see below).
VARIATION IN THE TYPE SERIES: Four males 22.3–23.1 SVL are adult; three females 24.6– 25.9 mm SVL are gravid. Proportions are summarized in tables 1 and 2.
The dorsal ground color in life is dark gray to graybrown with a more or less distinct dark, Wshaped mark in the scapular region. One specimen had an hourglassshaped rusty area occupying most of the middle of the back. Other features of the dorsal pattern may include a dark interocular bar and a dark mark in the postocularsupratympanic region. Black lumbar ocelli with white anterior edging typically are present. The groin and anterior and posterior surfaces of the thighs are yellow, with light gray mottling in some individuals. The chin, chest, and abdomen are gray, sometimes almost black, darker on the chin than posteriorly, with tiny white flecks. The iris is grayish gold to dark graybrown.
ADVERTISEMENT CALL: The call of Oreophryne hypsiops is a series of 20–28 notes (12 calls from five individuals) with a dominant frequency of 2900–3200 Hz uttered at about 7–10 notes per sec, and lasting about 2.2–3.3 sec (fig. 8B, table 3). The nature of the notes varies considerably, both within and among calls. The highenergy part may be pulsed or not, and ranges from about 0.04 to 0.07 sec in duration. A note may taper off gradually or abruptly, in some instances continuing weakly almost until the initiation of the next note (reverberation?). The range of temperatures at which recordings were made (24–27°C) is not wide enough to demonstrate an influence of temperature on the variables.
Calls appear to be given at relatively long and irregular intervals. In one recorded sequence, the second call was initiated 29 sec after the first, and the third 1 min 53 sec later.
COMPARISONS WITH OTHER SPECIES: Oreophryne hypsiops is sympatric with O. biroi and is abundantly distinct from that species; see Discussion under biroi (figs. 6, 7). See the account of O. parkeri for distinctions from that species.
HABITS AND HABITAT: Calling males were generally high, some probably 5 m or more above the ground. One male was seen on the underside of a nearly vertical banana leaf about 3 m above ground and another at a similar elevation atop a broad leaf in a spray of leaves at the top of sapling. A female was found on a much lower banana leaf, and one male on a leaf only about 1 m up.
DISTRIBUTION: The Adelbert Mountains and adjacent coastal region of Madang Province, Papua New Guinea, westward to the south slope of the Schrader Mountains, and probably northwest as far as the vicinity of Lumi, West Sepik Province; from sea level to at least 975 m. The three specimens from East Sepik and West Sepik Provinces conform closely to those from Madang Province in morphology, but the specific identity of frogs in these regions has not been confirmed by recordings of vocalizations .
LOCALITY RECORDS AND SPECIMENS EXAM INED: See Holotype and Paratypes.
Figure 16 View Fig
Cophixalus geislerorum: Loveridge, 1948: 423 View in CoL .
Oreophryne parkeri Loveridge, 1955: 3 View in CoL (type locality, ‘‘ Matapan [= Matapau], Australian New Guinea’ ’ [ West Sepik Prov., Papua New Guinea]; holotype MCZ A12964, collected by E.A. Briggs ‘‘about January, 1923’’).
TYPE LOCALITY: Matapau is a village on the coast at 143°00'E. The spelling ‘‘Matapau’’ appeared in Loveridge’s (1948: 423) first report on these specimens but was modified to ’’Matapan’’ in the 1955 description of the species. The former spelling is used in the ‘‘Village Directory’’ ( Territory of Papua and New Guinea, Department of Native Affairs, 1960) as well as on the Suain quadrangle, sheet 7491 of the 1:100,000 topographic map series, and we assume it to be the correct one.
DIAGNOSIS: Oreophryne parkeri is distinguished by its combination of narrow head (mean HW/SVL 0.325), small eyes (mean EY/SVL 0.109), short internarial span (mean IN/SVL 0.065), and a color pattern emphasizing numerous tiny white spots over all dorsal surfaces.
MORPHOLOGY: The holotype and only paratype are females with the following measurements and proportions ( paratype in parentheses): SVL 28.9 (25.6), HW 8.4 (7.8), TL 11.8 (10.1), EY 3.05 (2.5), EN 2.05 (2.2), IN 1.7 (1.6), HD 7.6 (7.5), FT 11.2 (10.3), FD 2.0 (penultimate phalanx 0.6), TD 1.7, 0.7 (1.4), TY 1.6 (1.3); HW/SVL 0.291 (0.305), TL/SVL 0.408 (0.395), EY/SVL 0.105 (0.098), EN/SVL 0.071 (0.086), IN/ SVL 0.059 (0.063), HD/SVL 0.263 (0.293), FT/SVL 0.387 (0.402), FD/SVL 0.069 (0.070), TD/SVL 0.059 (0.055).
The holotype and paratype are in poor condition, appearing somewhat shrunken so that the eye and head width measurements in particular may be shorter than in life. The following description generalizes from study of the type specimens and fresh material from Papua .
Snout bluntly rounded, almost truncate seen from above, vertical in profile, nares just visible from above, canthus rostralis rounded, loreal region sloping, slightly con cave; interorbital span 1.4 × 1.7 width of eyelid; tympanum moderately distinct to indistinct. Fingers unwebbed, relative lengths 3> 4 = 2> 1, 1st> onehalf 2nd, all with welldeveloped disks, subarticular elevations low, rounded (fig. 2C). Toe webbing reaches the subarticular elevations of the third and fifth toes but not that of the fourth, relative lengths 4> 5> 3> 2> 1, 5th only slightly longer than 3rd or the two essentially equal, disks broad but narrower than those of fingers, inner metatarsal elevation very indistinct, outer absent (fig. 2C). Supratympanic and hourglassshaped interscapular are weakly indicated. A ligament connects the procoracoid and scapula (examined on the holotype only).
The only major discrepancy between our assessment of the holotype and that of the describer involves the size of the eyes. Loveridge (1955: 4) stated ‘‘horizontal diameter of the eye... about equal to seveneighths its distance from the nostril.’’ By our measurement, the eye is almost 1.5× the eye– naris distance (1.1× in the paratype).
COLOR AND PATTERN: In preservative, ‘‘Above, brown, uniform except for some dark crossbars on the limbs and light areas in groin. Below, brown, paling posteriorly towards groin; thighs flecked with white, tibiae largely white’’ ( Loveridge, 1955: 4). The holotype is now faded brown dorsally with no conspicuous markings. The undersides have dense, fine brown mottling, essentially the same from chin through hind legs. The paratype is brown dorsally with paler spotting and mottling; flanks more coarsely mottled; conspicuous lumbar ocelli; top of head brown with no interocular bar; loreal area and upper lips brown with little trace of lighter color but no dark facemask effect; upper surfaces of thighs faintly mottled, shanks somewhat more so; posterior surfaces of thighs not distinctly patterned; chin and chest pale with dense melanic spotting that disperses on the abdomen.
In life, MZB 8155 (DP 95037) had the following colors and pattern: dorsum from snout to sacral region light brown with irregular darker small spots interrupted by a sharply defined, dark scapular ‘‘W’’; abundant tiny white spots over all dorsal and lateral surfaces and limbs; moderately distinct lumbar ocelli with a white anterior border; facial region—lips, below eye, loreal area— mottled dark and light brown with tiny white spots; groin and anterior of thigh white with a yellowish tinge and with a few brown speckles; posterior of thigh brown with white spots around cloacal opening, paler distally; ventral ground color white; throat with dense but pale graybrown speckle, chest and abdomen more mottled; underside of thigh mottled, of shank largely white. Each phalanx bears a narrow white band just proximal to the disk. The iris was dark brown. MZB 8156 had similar colors and pattern to MZB 8155 but with the skin more rugose with pointy warts emphasized by terminal white spots. The venter was melanistic with light areas of the abdomen appearing as spots rath er than as ground color; groin and anterior of thigh more brown than white; posterior of thigh coarsely mottled in brown and white.
VARIATION IN SIZE AND PROPORTIONS: The holotype, a gravid female (fide Loveridge, 1955; not verified by us as the specimen is stuffed with cotton) measures 28.9 mm SVL and the paratype, probably female, 25.6 mm SVL. Males of 23.0 and 23.8 mm SVL are adult. See tables 1 and 2 for body proportions.
ADVERTISEMENT CALL: The call is a train of 21 to 34 notes uttered in about 5 sec at a rate of about 5 per sec with a dominant frequency of 2600–2900 Hz (table 3, fig. 13B). The aural impression is of ‘‘peeps’’, sometimes slightly harsh. The envelope of individual notes can vary, even within a single train. At the simplest, a note can be unpulsed with the amplitude dropping abruptly but evenly near the end at about 0.07 sec. At the other extreme, the body of the note may have two or three distinct pulses with an additional weak pulse or two in a tail tapering off almost to the initiation of the succeeding note. The recordings were made within a narrow temperature span (24.2–26.3°C), so the call durations and note repetition rates can be expected to vary more than we report here.
COMPARISONS WITH OTHER SPECIES: Its narrow internarial span distinguishes O. parkeri from other north coast species except O. hypsiops . These two are similar in all proportions with broad overlaps in all ratios except for TD/SVL. No specimen of hypsiops showed the spotted pattern seen in the parkeri from Papua. The advertisement calls are similar, but differ in duration and in note repetition rate with no overlap in these parameters (table 3). We attach significance to the large size of the holotype of parkeri (female, SVL 28.9 mm) compared to that of the largest female hypsiops ( 25.9 mm), while recognizing that hypsiops may prove to be a synonym of parkeri with the Papua specimens representing an undescribed species.
Oreophryne albopunctata (van Kampen) from the Lorentz River in Papua, far to the southwest across the central ranges, resembles parkeri in three of the critical proportions ( HW, EY, IN) but has a more typical, wider internarial distance—IN/SVL 0.083 and 0.084 in the two syntypes .
HABITS AND HABITAT: The holotype was ‘‘taken near running water in Sago Palm Forest’’ ( Loveridge, 1948: 423) or ‘‘taken from running water in sagopalm forest’’ ( Loveridge, 1955: 3). Considering that no Oreophryne is known to be even partially aquatic, the former alternative is the more likely. At Sentani these frogs called from about 3 to 6 m above ground in banana plants and sago palms in an area of garden regrowth.
DISTRIBUTION: Known only from the type locality, a coastal village lying about midway between Aitape and Wewak, and from Sentani , Papua (fig. 9) .
LOCALITY RECORDS AND SPECIMENS EXAM INED: PAPUA NEW GUINEA: West Sepik Prov.; Matapau ( MCZ A 12964 [ holotype], BMNH 1955.1 .1.17, paratype, formerly MCZ A 12963). INDONESIA: Papua: Sentani (east end of airport, MZB 8155 View Materials , 8156 View Materials ) ; Iffar, Lake Sentani ( BMNH 1935.6.5.88) .
REMARKS: It is unfortunate that Oreophryne parkeri was described from two poorly preserved specimens. One problem we faced was whether to recognize the Papua specimens and parkeri as two species or to pool the specimens as parkeri . We have taken the latter conservative course, influenced by the probability that some, at least, of the seemingly distinctive morphology of the type and paratypes of parkeri may be due to their poor state of preservation. New material from the vicinity of the type locality is needed to verify our assignment of the specimens from Papua to this species (and, indeed, to verify the distinction from hypsiops ); recordings of the advertisement call would be virtually essential.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Oreophryne brachypus (Werner)
| ZWEIFEL, RICHARD G., MENZIES, JAMES I. & PRICE, DAVID 2003 |
Oreophryne geislerorum :
| Menzies, James I. 1980: 233 |
Oreophryne sp.
| Zweifel, Richard G. 1980: 411 |
Oreophryne brachypus : Tyler, 1964a: 4
| Tyler, Michael J. 1967: 187 |
| Tyler, Michael J. 1964: 4 |
Oreophryne parkeri
| Loveridge, Arthur 1955: 3 |
Cophixalus geislerorum : Loveridge, 1948: 423
| Loveridge, Arthur 1948: 423 |
Oreophryne biroi : Parker, 1934: 170
| Haupl, Michael 1994: 34 |
| Menzies, James I. 1976: 62 |
| Loveridge, Arthur 1948: 423 |
| Parker 1934: 170 |
Hyla brachypus :
| Zweifel, Richard G. 1960: 2 |
| Loveridge, Arthur 1948: 323 |
| Nieden 1923: 220 |
| Kampen 1923: 37 |
| Barbour, Thomas 1912: 177 |
Hylella brachypus : Vogt, 1911: 427
| Vogt, Theodor 1911: 427 |
Hylella brachypus
| Werner, Franz 1900: 121 |
| Werner, Franz 1898: 554 |
Cophixalus geislerorum
| Boettger 1892: 24 |