
Anolis, ELECTRUM
|
publication ID |
https://doi.org/ 10.1111/zoj.12159 |
|
DOI |
https://doi.org/10.5281/zenodo.10531067 |
|
persistent identifier |
https://treatment.plazi.org/id/0D5BB23D-CD79-FFB0-FC27-FBAB547DF8FD |
|
treatment provided by |
Carolina |
|
scientific name |
Anolis |
| status |
|
MORPHOLOGY OF ANOLIS ELECTRUM
During the fossilization process of amber, organic material fully surrounded by resin is preserved with remarkable detail. For vertebrate inclusions, the soft tissue, although apparently visible through the translucent amber, has actually rotted away, leaving an airfilled void lined by an impression of the skin. The skeletal elements are often preserved (particularly the limbs): they can remain in place, or may become disarticulated and free to move around inside the void. In both specimens, the lizard is surrounded by a reddish halo caused by the mineralization of the soft tissue that occurs when the amber is fractured and the organic matter comes into contact with air. From the HRXCT scans we found that the mineralized skin has a similar X-ray attenuation to bone, thereby obscuring the natural margin between the two materials. The HRXCT scans also revealed that there are few skeletal elements preserved in the holotype and paratype specimens, but that the outline of the air-filled voids retain remarkable details of the soft tissue ( Figs 1 View Figure 1 , 2 View Figure 2 , S 1 View Figure 1 and S 2 View Figure 2 ; Videos S1 and S2). Considering the anatomical parts preserved in each specimen, and the dimensions of the limbs in each, we agree with Lazell’s (1965) conclusion that these are two halves of the same animal. The following specimen descriptions are based primarily on CT scan reconstructions, and serve as a complement to the detailed scale information provid- ed by Lazell (1965).
Holotype ( UCMP 68496 View Materials , Figs 1 and S1 View Figure 1 )
This specimen includes the right hindlimb, a left hindtoe, and a portion of the body. The ventral aspect of the hindlimb and posterior portion of the body lie against the underside of the amber block ( Figs 1B and S1 View Figure 1 ). The only skeletal elements preserved in this piece are the parts of the forefeet. The hindlimb and abdomen are hollow, air-filled voids in the amber, the edges of which preserve great detail of the scales. The only part of the left foot that is preserved is most likely the fourth hindtoe, lying adjacent to the right lower leg; the most distal phalanx is preserved as skeleton, and there is an air-filled void surrounding the skin of the expand- ed toepad, outlining approximately two-thirds of the hindtoe. Phalanges and metatarsi of the right hindfoot are preserved in full and unbroken. An air-filled void outlines the right limb, specifically the hindtoes, foot, lower leg, and most of the thigh (rendered in green in Fig. 1A View Figure 1 ), and stops at the proximal end of the thigh. Skin of the upper thigh or groin has mineralized where the amber is fractured (rendered in white in Fig. 1A View Figure 1 ). Scales on the ventral side of the abdomen are visible, imprinted on the amber, with a yolk sac scar positioned medially. There are eight scales in 1 mm, measured along the yolk sac scar. The skin in the anteriormost and posteriormost areas of the abdomen has also mineralized. No skeletal elements are preserved in this region.
Reconstructing air-filled voids in the amber from the HRXCT data revealed details of an invertebrate inclusion. The specimen, an ant of the family Formicidae , genus Azteca (C. Moreau, pers. comm.), is very well preserved, lying adjacent to the right hindfoot ( Fig. 1C View Figure 1 ). The total length of the body is 3.6 mm, the width of the head is 0.8 mm, and the length of one antenna is 1.9 mm. Lazell (1965) did not remark on this inclusion.
Paratype ( UCMP 68497 View Materials , Figs 2 and S2 View Figure 2 )
This specimen includes the anterior portion of the body, and the preserved skeletal elements are restricted to the forelimbs and to the skull. The forelimbs and dewlap are visible through both sides of the amber block ( Fig. S2 View Figure 2 ). Despite this, Lazell (1965) indicated the absence of a throat fan. The skeletal elements preserved are mainly of the forelimbs. Of the right forelimb, the humerus is preserved, but the proximal end is not well defined. The ulna and radius are preserved, although they are not perfectly aligned with each other. The metacarpals and phalanges of all five foretoes are also preserved. The wrist appears to be broken so that the bones are not continuous as in life, but slightly separated. The ends of the limb bones appear to be square-ended, indicating the epiphyses have not yet fused and thus that the specimen is a juvenile. Of the left forelimb, only the phalanges and metacarpals are preserved. An air-filled void in the amber outlines the upper and lower parts of the forelimb, showing details of the scales. The proximal end of the left forelimb shows mineralization of the skin where it meets the torso.
The trunk, neck, and head are preserved as an airfilled void that is open to the edge of the amber piece. The trunk is broken where the ribcage (not preserved) would be. The skin of these body parts has mineralized around the periphery of the void. The head is partially preserved. Figure 2B View Figure 2 shows the head from ventral view, in which most of the skull has broken away at the edge of the amber piece. The head appears to have been severed at an angle across the rostrum, removing the left side of the head and the right side, from the middle of the mouth forwards. The right side of the skull is preserved in part ( Fig. 2C View Figure 2 ): the lateral margins of the parietal are preserved, adjacent to the postorbital bone and posterolateral part of the frontal bone. Mineralized soft tissue obscures the lateral portion of the skull, but the quadrate is visible. The squamosal may be preserved, but if so, it is obscured by the mineralized tissue. The surangular is preserved and continuous with the dentary bone. Only the posterior half of the dentary is preserved. The coronoid process is visible behind the jugal. The jugal is almost completely preserved: a portion is missing at the boundary between the jugal and postorbital bar. The maxilla is articulated with the jugal and lies anterior to the dentary, as in life, up to the edge of the amber. Only the posterior half of the maxilla is preserved. Four maxillary teeth and four or five dentary teeth are preserved, all tricuspid. The right half of the palate, the right pterygoid, and ectopterygoid are preserved in place. The presence of pterygoid teeth could not be confirmed given the level of mineralization in the surrounding tissue.
From the HRXCT renderings of the fossils, we update the morphometric and meristic data provided by Lazell (1965; his measurements are given in parentheses, when available): the thigh is 5.5 mm (5.2 mm) and the lower leg is 4.5 mm (4.1 mm), measured from the insertion of the limb on the body wall to the knee joint, and from this point to the point of inflexion on the heel, respectively. The distance from the heel to the base of hindtoe IV is 3.2 mm, and from the base to the tip is 4.4 mm, giving a total of 7.7 mm (7.0 mm). The toepad of right hindtoe IV is 0.5 mm (0.9 mm) at the widest part. We count between 17 and 20 (21 or 22) lamellae under the third and fourth phalanges of right hindtoe IV, depending on the landmark used for counting (lamellae are counted from the most distal end of the third phalanx to the most proximal end of the fourth phalanx, with the latter landmark usually identified by bending the hindtoe). In addition, the preserved section of the torso is 3.2 mm wide and 6.2 mm long. From the estimated thigh length, we estimate the snout–vent length (SVL) of A. electrum to be 24 mm, based on juvenile and adult data of Anolis brevirostris Bocourt, 1870 , Anolis coelestinus Cope, 1862 , and Anolis cybotes Cope, 1862 (T. Sanger, unpublished data), using the equation logSVL = logThigh/ 1.01–0.65. Given the range of hindlimb variation in these three species, the SVL of the specimen could range between 20 and 28 mm. The left upper arm is 3.1 mm, measured from the limb insertion to the elbow joint, and the right humerus is 3.0 mm (3.9 mm), although this is an underestimate as the most proximal end (the epiphysis and a small length of the diaphysis) is missing. The left forearm is 3.6 mm (3.2 mm), measured from the elbow to the wrist, and the right ulna and radius are each 2.2 mm. The left forefoot is 3.5 mm from the wrist to the tip of foretoe IV (3.6 mm). The preserved limb bones are only represented by the diaphyses; the epiphyses are not preserved, therefore the limb lengths reported are underestimates of the total bone length. The section of lower jaw preserved (from the posteriormost point on the surangular to the anteriormost point on the broken dentary) is 4.3 mm long, and the preserved section of maxilla is 1.7 mm. The height of the head at the parietal is ∼ 2.7 mm. Lazell (1965) measured 4.8 mm for the head ‘at the level of the interparietal’, but there is no reliable boundary of the back of the skull preserved and visible on the HRXCT scan.
PHYLOGENETIC ANALYSES
Ten of the 91 morphological characters described in Poe (2004) were scored for Anolis electrum . In agreement with Lazell (1965), we scored the following characters: digital pad of the ‘raised’ type (i.e. toepads overlap second phalanx, or ‘alpha type’, as opposed to toepads non-overlapping the second phalanx, or ‘beta type’), absence of enlarged mid-dorsal scales (as opposed to presence of mid-dorsal scales larger than surrounding scales), ventral scales arranged in transverse rows (i.e. each ventral scale is bordered posteriorly by two scales, as opposed to arranged in diagonal rows, in which each ventral scale is bordered posteriorly by three scales), interparietal scale bordered posteriorly by small scales gradually transitioning into dorsal granules (as opposed to mid-nuchal scales in rows of bulbous scales distinct from dorsal scales), dorsal and ventral scales smooth (as opposed to keeled), and supradigital scales keeled (as opposed to smooth). Additionally, we scored: (1) the preoccipital scale absent (as opposed to present); (2) the fold of skin over the dorsal rim of the ear opening absent (as opposed to present); (3) the interparietal scale separated from the supraorbital semicircles by one or more rows of scales (as opposed to in contact with supraorbital semicircles); (4) the posteroventral corner of the jugal convex (as opposed to concave); and (5) the coronoid labial process present (as opposed to absent). The dewlap is extended completely and is attached posteriorly to the level of the arm insertion. The imprint left by the skin on the amber reveals that scales covering the dewlap are scattered throughout the skin, as opposed to organized in rows ( Figs 2A and S2F View Figure 2 ).
Köhler (2014) recently compiled a list of characters useful in taxonomic descriptions. Several of these (condition of terminal phalanx, number of rows of enlarged dorsal scales, condition of supradigital, ventral, and dorsal scales, number of scales between supraorbital semicircles and interparietal scale, and size of scales adjacent to interparietal scale) are included in our phylogenetic analysis. Three other characters can be scored in A. electrum , but were not included in the phylogenetic reconstruction because comparable data are not available for many species. These characters are: (1) diameter of parietal scale, longitudinal = 0.95 mm, transverse = 0.48 mm; (2) subdigital pad width (toe IV), forefoot = 0.58 mm, hindfoot = 0.51 mm; and (3) condition of parietal depression, deep. Researchers interested in using these traits should bear in mind that the A. electrum specimen is a juvenile, and thus data may not be comparable with that obtained from adults of extant species.
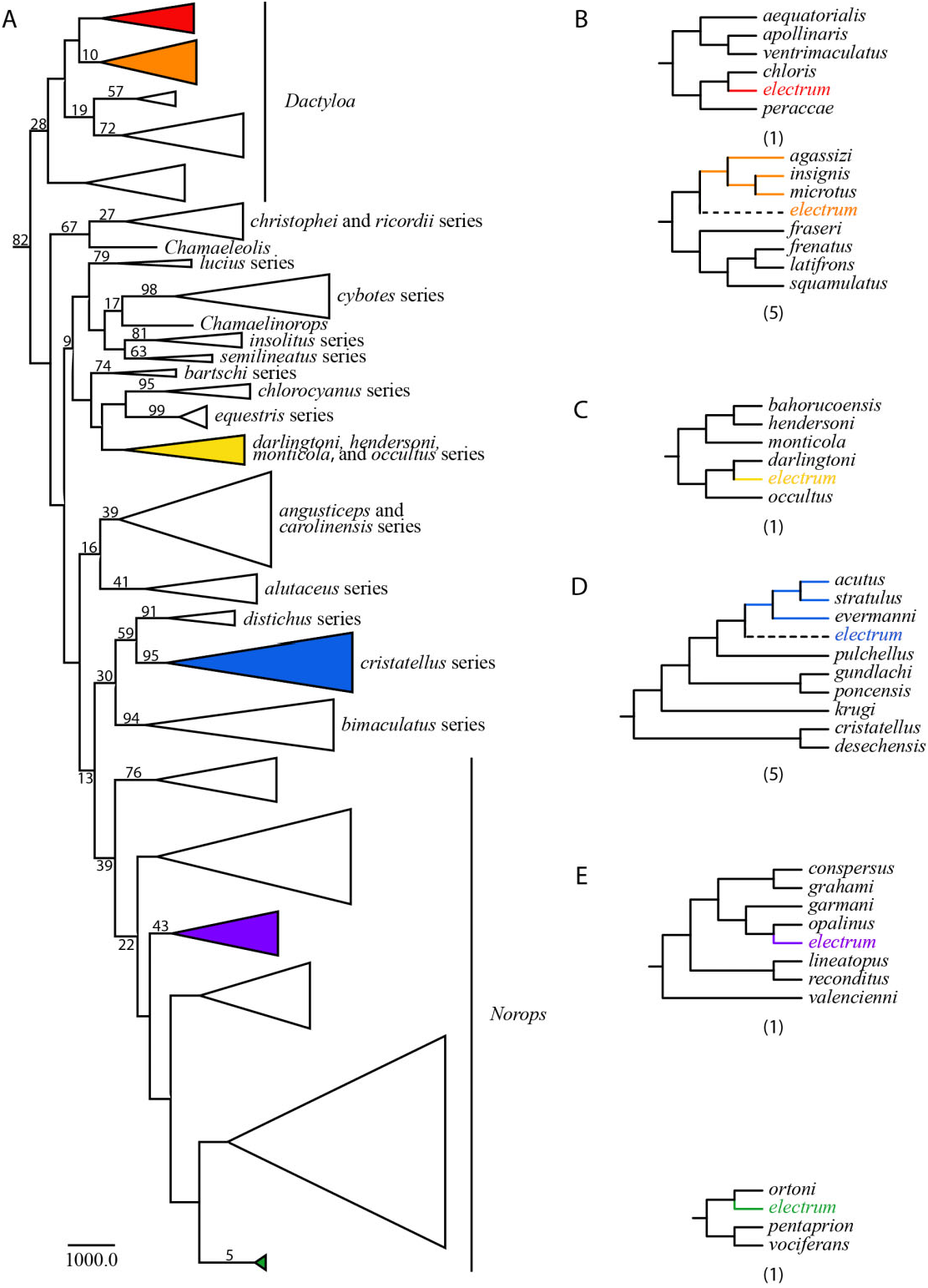
One hundred bases corresponding to sections of the tRNAs and the O L were excluded from the analyses because of ambiguous alignment. The resulting matrix includes 1374 bp, 91 morphological characters, and 182 taxa, 41 of which are missing molecular data. The parsimony analysis yielded 14 fully resolved most parsimonious trees of 224 233.85 steps (consistency index, CI = 0.089; retention index, RI = 0.517), which only differ in the position of A. electrum . All major subclades of Anolis (as shown in previous phylogenetic analyses; Jackman et al., 1999; Nicholson, 2002; Castañeda & de Queiroz, 2013) are inferred, except the sagrei series within the Norops clade, which was inferred to be paraphyletic. Anolis electrum is placed in 14 alternative positions ( Fig. 3B–E View Figure 3 ): six within the Dactyloa clade ( Fig. 3B View Figure 3 ), sister to Anolis darlingtoni (Cochran, 1935) ( Fig. 3C View Figure 3 ), five within the cristatellus series ( Fig. 3D View Figure 3 ), and two within the Norops clade ( Fig. 3E View Figure 3 ). A list of synapomorphies supporting the alternative sister relationships of A. electrum is provid- ed as Supporting Information Appendix S1. The maximum agreement subtree (or common pruned tree; Finden & Gordon, 1985), which results from excluding the same set of taxa from the primary trees (in this case A. electrum ), is shown in Figure 3A. View Figure 3


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.