
Seira pallidipes Reuter, 1895
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4312.3.5 |
|
publication LSID |
lsid:zoobank.org:pub:A9544653-220C-4555-Ad0E-Ef2D34344695 |
|
DOI |
https://doi.org/10.5281/zenodo.6021272 |
|
persistent identifier |
https://treatment.plazi.org/id/0E45486F-FF83-FFA0-FF20-F9EE66F3FD92 |
|
treatment provided by |
Plazi |
|
scientific name |
Seira pallidipes Reuter, 1895 |
| status |
|
Figs 1–42 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 – 10 View FIGURE 11 View FIGURE 12 – 14 View FIGURE 15 – 17 View FIGURE 18 – 20 View FIGURE 21 – 27 View FIGURE 28 View FIGURE 29 View FIGURE 30 View FIGURE 31 – 35 View FIGURE 36 – 41 View FIGURE 42
Sira (sic!) pallidipes Reuter, 1895: 114 ; Denis 1941: 45 (incertae sedis); Gisin 1944: 81 (mentioned as insufficiently described).
Seira pallidipes Reuter : Palissa 1964: 220 (mentioned as insufficiently described); Gisin 1960: 237 (species inquirenda); Loksa & Bogojević 1970: 133, Figs 36–37 View FIGURE 36 – 41 (description); Ellis 1976: 316 (keyed); MócZár & Bíró 1980: 153; Loksa 1983: 74; Loksa 1987: 79; SZeptycki 1979: 41 (chaetotaxy based on Loksa & Bogojević 1970); Kampichler 1991: 150.
Seira pallipes (sic!) Reuter: Hornung 1986: 138.
Pseudosira (sic!) pillichi Stach, 1930: 299 syn. nov.; Gisin 1944: 83 (keyed).
Lepidocyrtinus pillichi Stach : Denis 1941: 45, 87, 92 (keyed).
Seira pillichi Stach : Gisin 1960: 237 (species inquirenda); Palissa 1964: 218 (keyed, diagnosis, probable synonymy with pallidipes mentioned)
? Seira pillichi f. decolorata Loksa & Bogojević, 1970: 135, Fig. 38 View FIGURE 36 – 41 . non! Seira pillichi graeca Ellis, 1966: 142 , Figs 2 View FIGURE 2 a–e, 3a–h (valid species as Seira graeca Ellis, 1966 – according to Ellis 1976); SZeptycki 1979: 100.
non! Seira pillichi (Stach) : SZeptycki 1979: 41.
Material examined. Holotype, sex undeterminable ( HNHM coll-202): Kecskemét, Hungary, 01.x.1891, leg. L. Bíró ; 24 topotypic specimens (3 males, 1 female and 1 juvenile on slide; 11 adult and 8 juvenile specimens in alcohol): Bugacpuszta , Kecskemét, 107 m, N46°39'58"; E19°37'39", hand collecting, 6.iii.2017, leg. D. Winkler GoogleMaps ; 34 specimens (2 males, 1 female on slide; 14 adult and 13 juvenile specimens in alcohol; 4 adult specimens critical point dried and coated with gold–palladium for electron microscope scanning): Simontornya (type locality of S. pillichi Stach, 1930 ), Hungary, Barcsi Valley, hillside with loess steppe pasture, N46°45'59.28", E18°31'49.56", 115 m, D-vac sample, 14.iv.2016, leg. L. Dányi & N. Flórián ( HNHM coll-947). GoogleMaps
Other material examined. 8 specimens (1 male on slide, 4 adult and 3 juvenile specimens in alcohol): Börzsöny Mountains, Mount Nagy-Koppány, com. Pest ( Hungary), 549 m, N47°53'55"; E18°51'08", sub-pannonic grassland, hand collecting, 14. x. 2014, leg. D. Winkler & Á. Erdő GoogleMaps ; 14 specimens (1 female on slide, 7 adult and 6 juvenile specimens in alcohol) from Mount Somló , Mid-West Hungary, 360 m, N47°8'42"; E17°22'34", subpannonic grassland, mouth operated aspirator, 14.iii.2016, leg. D. Winkler. GoogleMaps
Redescription. Large sized Seira species (adult body length 2.1–2.8 mm). Holotype body length 2.75 mm according to the original description, but not measurable exactly in the strongly collapsed specimen. Main body colour dark blackish blue in alcohol ( Fig. 2 View FIGURE 2 ), living specimen with green-to-blue metallic lustre ( Fig. 3 View FIGURE 3 ). Apart from head and trunk, coxae, ventral tube and first 4/5 part of Ant. I blackish blue, some dark shade (depending on specimen) also on antennal segments and manubrium. Apical part of Ant. I, Ant. II–IV, legs and furca yellowish. Head and trunk densely covered by brownish scales of different shape (oval, short or elongated with apex rounded, pointed, truncate or irregular). Ant. I–II, basal part of Ant. III (occasionally also basal part of Ant. IV), legs except for foot complex, anterior and posterior side of ventral tube, manubrium and dens (ventrally) with scales ( Fig. 31 View FIGURE 31 – 35 ).
Antennae long, antennal length to head diagonal length ratio around 2.5 (head diagonal measured from the cervical edge to the apex of the mouth part). Relation of antennal joints I–IV as 1:2.3:2.8:4.1.
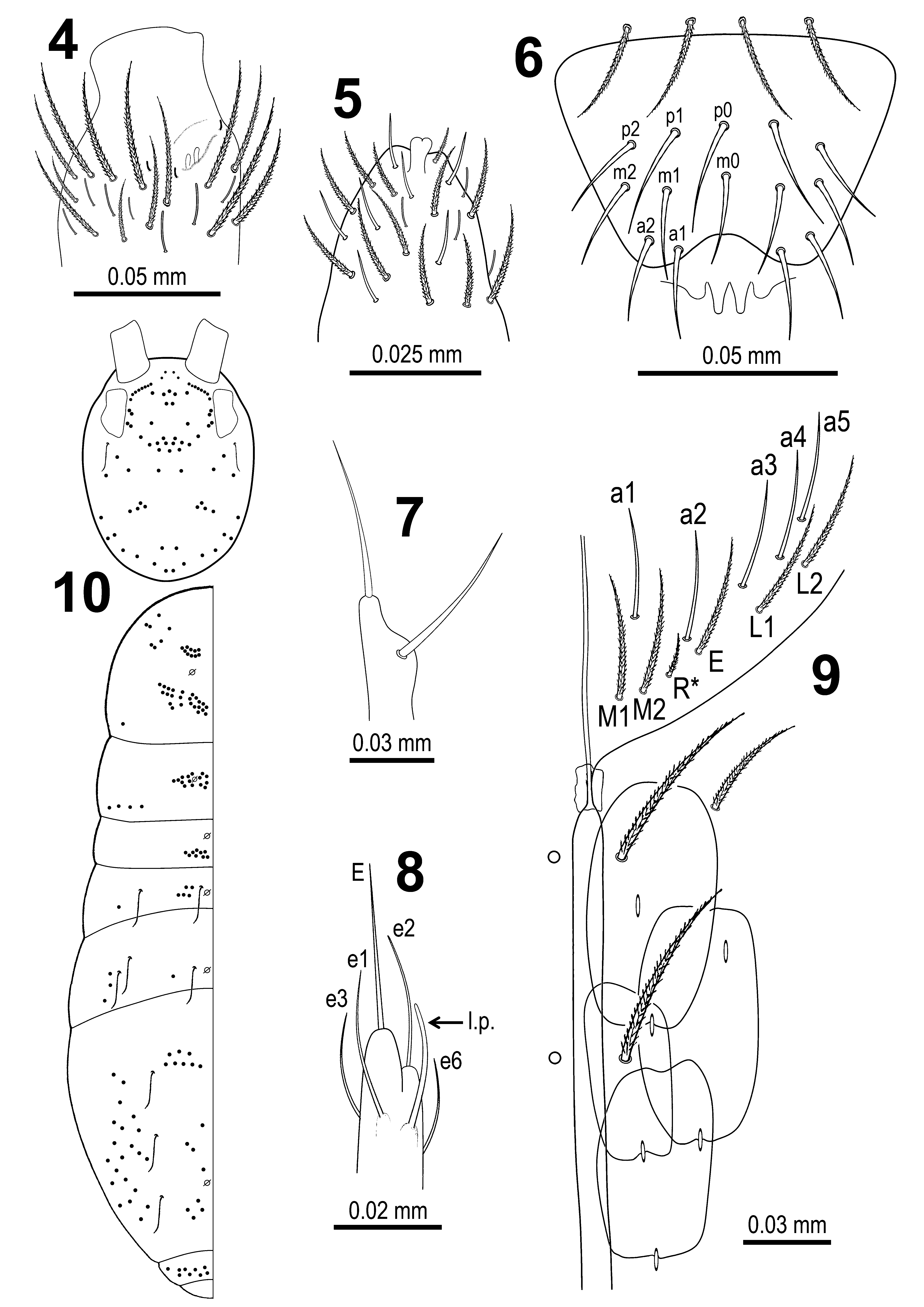
Ant. III sensillary organ composed by two sensory rods partially behind a cuticular fold ( Fig. 4 View FIGURE 4 – 10 ). Ant. IV weakly annulated, with bilobated apical bulb ( Figs 5 View FIGURE 4 – 10 , 34 View FIGURE 31 – 35 ).
Eyepatches oval, with largest ocellus B and smallest ocelli G and H, with three interocular mesochaetae (p, v, t) ( Figs 11 View FIGURE 11 , 33 View FIGURE 31 – 35 ).
Arrangement of chaetae on labrum 4/554, prelabral chaetae ciliated, posterior, median and anterior labral chaetae smooth. Labral chaetae p0 and p 1 in posterior row longer than others ( Figs 6 View FIGURE 4 – 10 , 32 View FIGURE 31 – 35 ). Labrum with four distinguishably separated labral papillae with middle two protruding, cone-shaped ( Figs 6 View FIGURE 4 – 10 , 32 View FIGURE 31 – 35 ). Outer maxillary palp with two smooth chaetae ( Figs 7 View FIGURE 4 – 10 , 32 View FIGURE 31 – 35 ) and three smooth sublobal chaetae ( Fig. 32 View FIGURE 31 – 35 ). Labium with five smooth proximal chaetae. Labial palp with five papillae ( Fig. 36 View FIGURE 36 – 41 ), papilla A consisted of one terminal sensillum (A); papilla B with one terminal sensillum (B) and five smooth guard chaetae (a1, b1, b2, b3 and b4); papilla C with one terminal sensillum (C); papilla D with one terminal sensillum (D) and four smooth guard chaetae (d1, d2, d3 and d4 – latter one not visible on Fig. 36 View FIGURE 36 – 41 ); papilla E ( Fig. 8 View FIGURE 4 – 10 , 36 View FIGURE 36 – 41 ) with one terminal sensillum (E), four smooth guard chaetae (e1, e2, e3 and e6) and curved lateral process (sensu Fjellberg 1999) surpassing top of papilla. Hyaline plate with hypostomal papilla H ( Fig. 36 View FIGURE 36 – 41 ) with one terminal sensillum and two smooth guard chaetae (h1 and h2).
Labium chaetotaxy formed by 5 smooth "A" chaetae and in the basal row by ciliated chaetae M1M2R*EL1L2 ( Figs 9 View FIGURE 4 – 10 , 35 View FIGURE 31 – 35 ) with R* smaller than other chaetae (ratio of R*/M ~0.4). Ventral cephalic grove with 2+2 ciliated chaetae, basal chaeta absent ( Fig. 9 View FIGURE 4 – 10 ).
Distribution pattern of dorsal macrochaetae on head and trunk as in Fig. 10 View FIGURE 4 – 10 . Complete dorsal head chaetotaxy as in Fig. 11 View FIGURE 11 . Antennal series ‘An’ with a maximum of 10+10 acuminate macrochaetae, microchaeta An2a typical for Seira absent; anterior series ‘A’ with A4 absent, A0, A2, A3 and A5 as macrochaetae, A1 as microchaeta; medio-ocellar series ‘M’ with 5+5 chaetae, M1, M2, M 4i and M4 as macrochaetae, M3 as microchaeta; sutural series ‘S’ with macrochaetae S0, S1, S2, S3, S4, S5, S6 and S7, respectively); post-sutural series ‘Ps’ with 3+3 chaetae, Ps2 as macrochaeta, Ps5 as mesochaeta, Ps3 as microchaeta, Ps1 and Ps4 absent (typical of Seira spp., see Soto-Adames 2008); postoccipital anterior series ‘Pa’ with 6+6 chaetae, Pa1, Pa2, Pa3, Pa4, Pa5 as macrochaetae, Pa6 as trichobotrium; postoccipital media series ‘Pm’ with 2+2 macrochaetae (Pm1 and Pm3), Pm2 absent (typical of Seira ); postoccipital posterior series ‘Pp’ with 6+6 chaetae, Pp1, Pp2, Pp3, Pp5 as macrochaetae, Pp4 and Pp6 as microchaetae; postoccipital external series ‘Pe’ with 4+4 macrochaetae (Pe2, Pe3, Pe3a and Pe5, respectively).
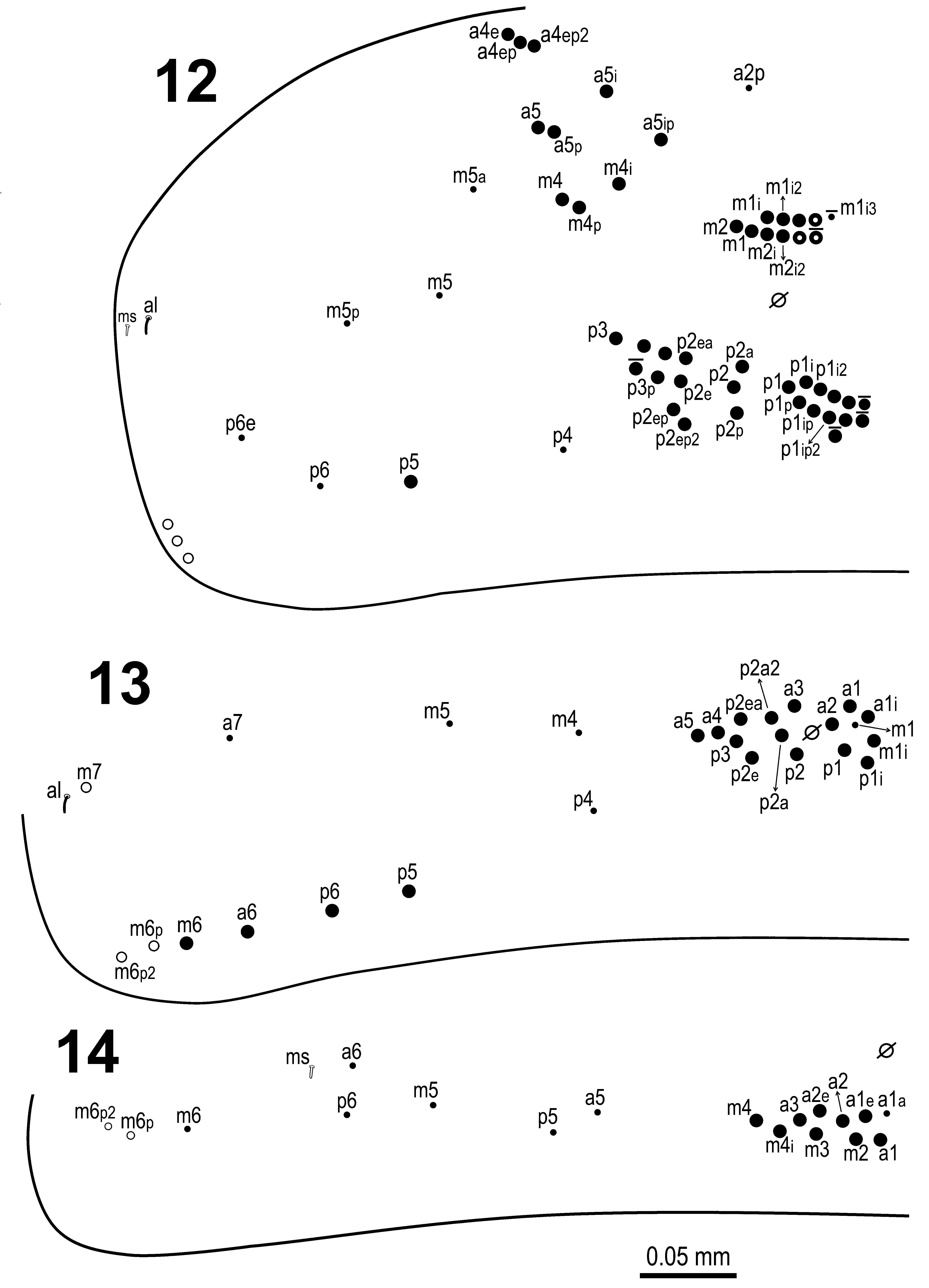
Mesothorax dorsal chaetotaxy as in Figs 12 View FIGURE 12 – 14 , 28–29 View FIGURE 28 View FIGURE 29 , 37–38 View FIGURE 36 – 41 . Anterior series ‘a’ (without anterior setal collar) with 8+8 chaetae ( Fig. 12 View FIGURE 12 – 14 ), a4e, a4ep, a4ep2, a5, a5p, a 5i and a5ip as macrochaetae, a2p as microchaeta; medial series ( Figs 28 View FIGURE 28 , 37 View FIGURE 36 – 41 ) with 15+15 to 17+17 chaetae, m1, m 1i, m 1i 2, m2, m 2i, m 2i 2, m4, m4p, m 4i as macrocahetae, one macrochaeta with uncertain homology next to m 1i 2, three additional unnamed macrochaetae (occasionally present as microchaetae), m5, m5a, m5p and m 1i 3 (present or absent) as microchaetae; posterior series ( Figs 29 View FIGURE 29 , 38 View FIGURE 36 – 41 ) with 24+24 to 28+28 chaetae, “PmA” group (see Soto-Adames 2008) with 9 to 12 macrochaetae (p1, p1p, p 1i, p1ip, p 1i 2, p1ip2 and six macrochaetae of uncertain homology, three of them present or absent); “PmB” group with three macrochaetae (p2, p2a, p2p); “PmC” group with 8 to 9 macrochaetae (p2e, p2ep, p2ep2, p2ea, p3, p3p and three macrochaetae of uncertain homology, one of them present or absent). Chaeta p5 as macrochaeta, p4, p6 and p6e as microchaetae. Three lateral mesochaetae external to p6 present. Lateral sensilla al and Smicrochaeta ms also present.
Metathorax dorsal chaetotaxy as in Fig. 13 View FIGURE 12 – 14 . Series ‘a’ with 8+8 chaetae, a1, a 1i, a2, a3, a4, a5 and a6 as macrochaetae, a7 as microchaetae; series ‘m’ with 8+8 chaetae, m 1i and m6 as macrochaetae, m6p, m6p2 and m7 as mesochaetae, m1, m4 and m5 as microchaetae; series ‘p’ with 11+11 chaetae, p1, p 1i, p2, p2e, p2a, p2a2, p2ea, p3, p5 and p6 as macrochaetae, p4 as microchaeta. Lateral sensilla (S-chaeta) al external to mesochaeta m7 also present.
Abd. I dorsal chaetotaxy as in Fig 14 View FIGURE 12 – 14 . Series ‘a’ with 8+8 chaetae, a1, a1e, a2, a2e and a3 as macrochaetae, a1a, a5 and a6 as microchaetae; series ‘m’ with 8+8 chaetae, m2, m3, m4 and m 4i as macrochaetae, m6p and m6p2 as mesochaetae; m5 and m6 as microchaetae, and with S-microchaeta (ms) next to microchaeta a6; series ‘p’ with 2+2 microchaetae (p5 and p6).
Abd. II dorsal chaetotaxy as in Fig. 15 View FIGURE 15 – 17 . Series ‘a’ with 7+7 chaetae, a2 and a3 as macrochaetae, a6 and a7 as microchaetae, as as sensilla (S-chaeta), a2p as fan-shaped microchaeta, a5 as trichobotrium with five surrounding fan-shaped microchaetae; series ‘m’ with 8+8 chaetae, m3, m3e, m 3ei and m5 as macrochaetae, m3ea, m6 and m7 as microchaetae, m2 as trichobotrium with five surrounding fan-shaped microchaetae; series ‘p’ with 3+3 chaetae, p5 and p7 as microchaetae, p6 as micro- or mesochaeta; sensilla (S-chaeta) acc.p6 present above p6. Lateral microchaeta el also present.
Abd. III dorsal chaetotaxy as in Fig. 16 View FIGURE 15 – 17 . Series ‘a’ with 7+7 chaetae, a7 as mesochaeta or microchaeta, a1 and a2 as fan-shaped microchaetae, a3 and a6 as microchaetae, as as sensilla (S-chaeta), and a5 as trichobotrium; series ‘m’ with 8+8 chaetae, m3, am6, pm6 as macrochaetae, m4 and m3ea as fan-shaped microchaetae, m7 as microchaeta, m2 and m5 as trichobotria; series ‘p’ with 6+6 chaetae, with p6 as macrochaeta, p7 and p8 as mesochaetae, p3, p5 and p 7i as microchaetae. Sensilla (S-chaeta) acc.p6 between macrochaetae p6 and pm6, and S-microchaeta ms next to p5 also present. Trichobotrium m2 surrounded by five fan-shaped microchaetae (including a1, a2, p3 and two unnamed chaetae). Trichobotrial complex around trichobotria a5 and m5 ( Figs 17 View FIGURE 15 – 17 , 39 View FIGURE 36 – 41 ) include c3 as fan-shaped or smooth microchaeta, 10 fan-shaped microchaetae (m3ea, m4, em, emp and additional unnamed chaetae) and microchaeta a6. Lateral microchaeta el and an additional unnamed micro- or mesochaeta also present.
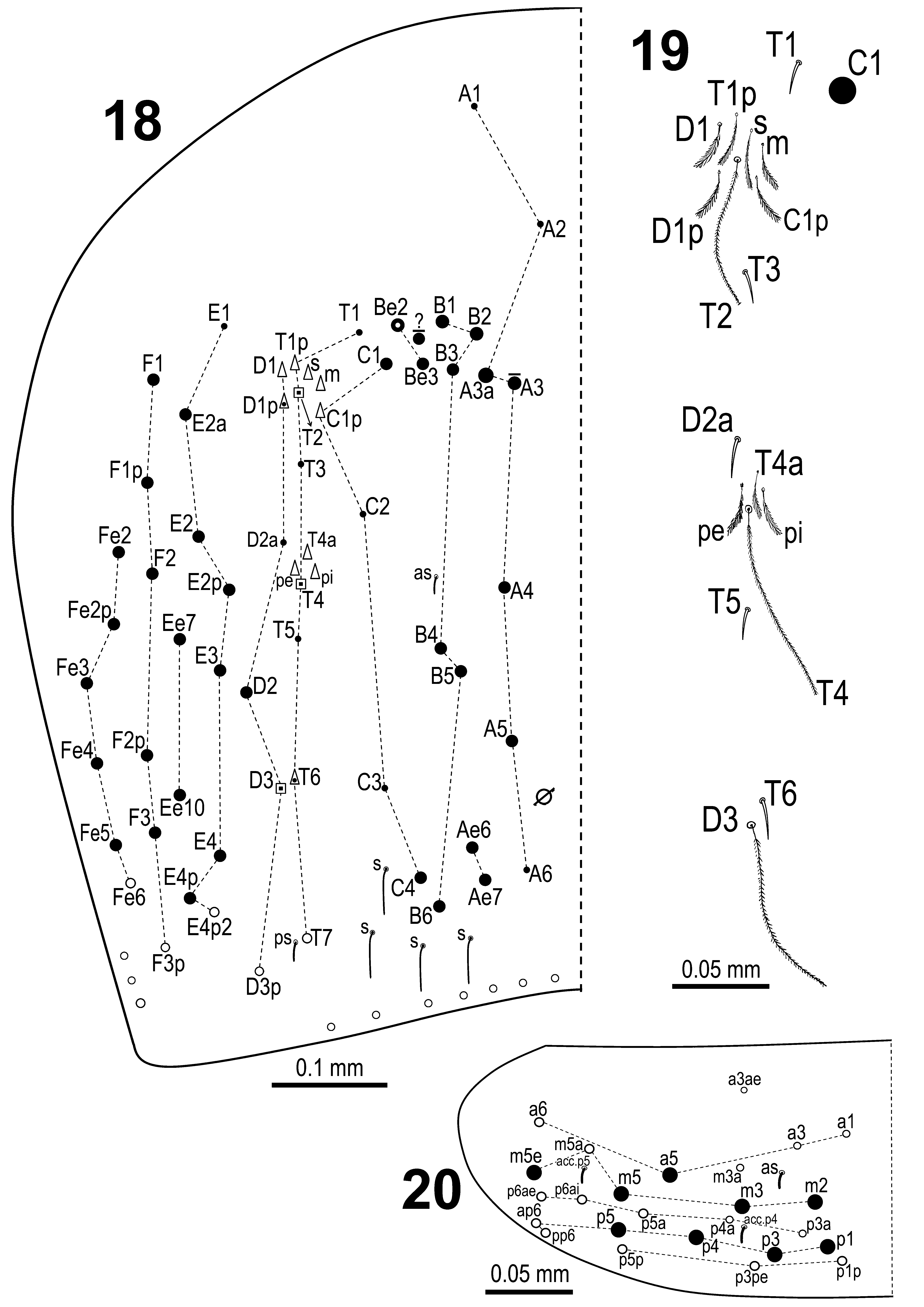
Abd. IV dorsal chaetotaxy and trichobothrial complex on Abd. IV as in Figs 18–19 View FIGURE 18 – 20 . Series A with 7+7 chaetae, A3, A3a (long and acuminate), A4, A5 as macrochaetae, A1, A2 and A6 as microchaetae; series ‘Ae’ with two macrochaetae (Ae6 and Ae7, latter one as long acuminate macrochaeta); series ’B’ with 6+6 macrochaetae (B1, B2, B3, B4, B5 and B6); series ’Be’ with 2+2 chaetae, Be3 as macrochaeta and Be2 as macro or rarely microchaeta; series ’C’ with 5+5 chaetae, with C1 and C4 as long acuminate macrochaetae, C1p as fan-shaped microchaeta, C2 and C6 as microchaetae; series ’T’ with 9+9 chaetae, with T7 as mesochaeta, T1, T3 and T5 as microchaetae, T6 as fan-shaped chaeta or smooth microchaetae, T2 and T4 as trichobotria with associated fanshaped microchaetae T1p, s and m (around T2) and T4a, pe and pi (around T4); series ’D’ with 6+6 chaetae, with D2 as long acuminate macrochaeta, D3p as mesochaeta, D1 as fan-shaped microchaeta, D1p as fan-shaped or smooth microchaeta, D2a as microchaeta and D3 as trichobotrium; series ’ E’ with 8+8 chaetae, with E2a, E2, E2p, E3, E4 and E4p as macrochaetae, E4p2 as mesochaeta and E1 as microchaeta; series ’Ee’ with two macrochaetae (Ee7 and Ee10); series F with 6+6 chaetae, F1, F1p, F2, F2p, F3 as macrochaetae, F3p as mesochaeta; series ’ Fe’ with 6+6 chaetae, Fe2, Fe2p, Fe3, Fe4, Fe5 as macrochaetae, and Fe6 as mesochaeta. Additional macrochaeta of uncertain homology within region Be2-Be3-B1 occasionally present. S-chaetae as anteriorly to macrochaeta B4 and ps next to mesochaeta T7 present. Four elongated posteromedial sensillae also present.
Abd. V dorsal chaetotaxy as in Fig. 20 View FIGURE 18 – 20 . Series ’a’ with 5+5 chaetae, a5 as macrochaeta, a1, a3, a3ae and a6 as mesochaeta; series ’m’ with 6+6 chaetae, m2, m3, m5 and m5e as macrochaetae, m3a and m5a as mesochaetae; posteroanterior series ’pa’ with 5+5 mesochaetae (p3a, p4a, p5a, p6ai and p6ae); series ‘p’ with 6+6 chaetae, p0 absent, with p1, p3, p4 and p5 as macrochaetae, ap6 and pp6 as mesochaetae; posterior-posterior series ‘pp’ with 3+3 mesochaetae (p1p, p3pe, p5p); three S-chaetae (as, acc.p4 and acc.p5) typical of Seira also present.
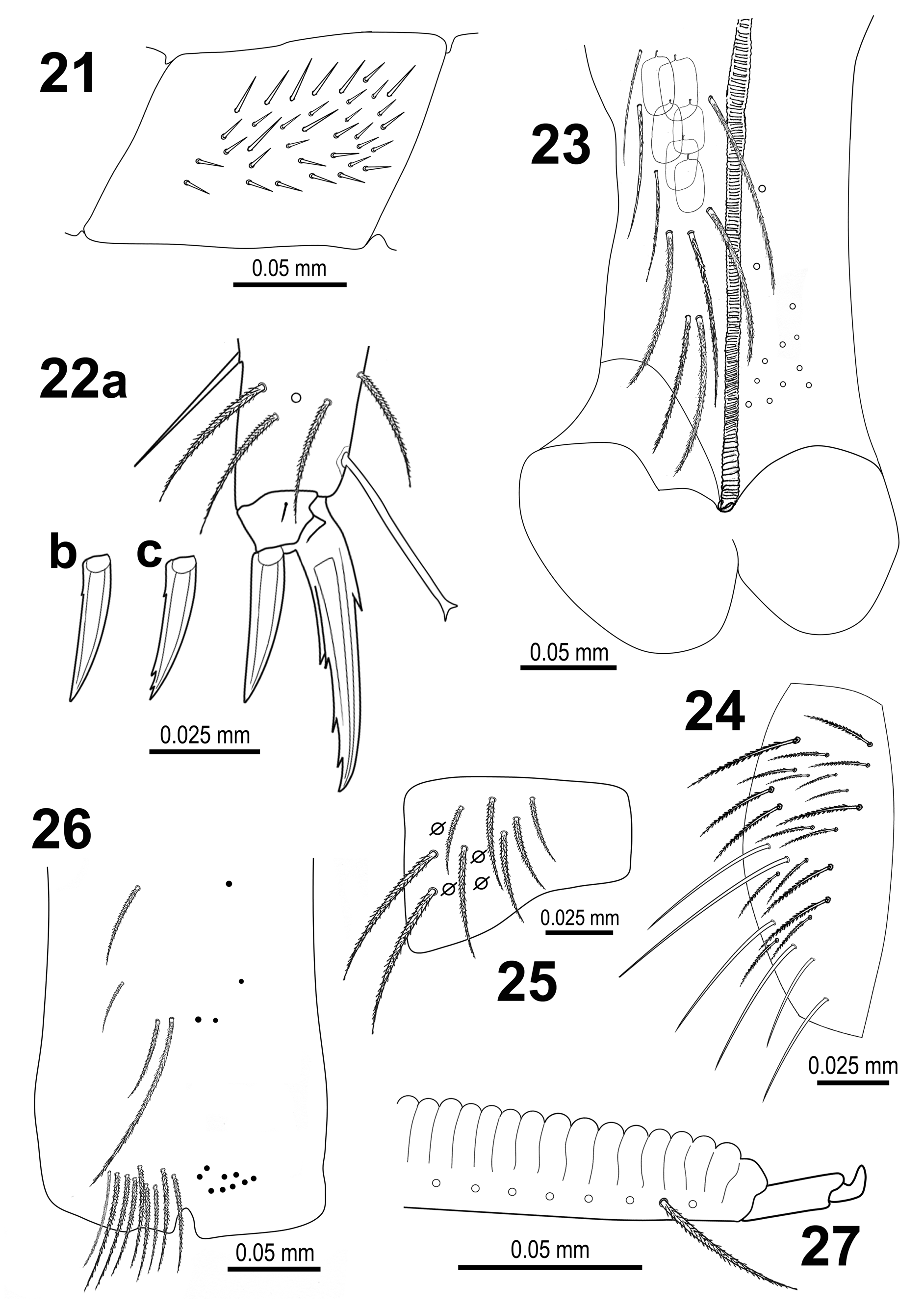
Legs. Trochanteral organ with 26–38 smooth spiny chaetae forming a rectangular shape pattern ( Figs 21 View FIGURE 21 – 27 , 40 View FIGURE 36 – 41 ). Unguis and unguiculus of claw III as in Figs 22 View FIGURE 21 – 27 a–c and Fig. 41 View FIGURE 36 – 41 . Unguis with sub-equal paired basal teeth distinct (at 50%) from the inner edge, and with two more unpaired teeth at 74% and 87% from inner edge, respectively.
A pair of outer teeth also present. A small pretarsal chaeta present on both anterior and posterior surfaces. Unguiculus lanceolate, outer lamella smooth ( Fig. 22 View FIGURE 21 – 27 a) or with one basal denticle ( Fig. 22 View FIGURE 21 – 27 b), occasionally also with 2–3 small apical denticles ( Fig. 22 View FIGURE 21 – 27 c), depending on specimen; other lamellae smooth. Tibiotarsal tenent hair spatulate, weakly ciliated, as long as claw. Ratio of supraempodial chaeta (smooth chaeta on tibiotarsus III opposite to the tenent hair) / unguiculus about 0.7.
Ventral tube ( Fig. 23 View FIGURE 21 – 27 ) with 9+9 ciliated chaetae on anterior side and 12+12 ciliated chaetae on posterior side. Lateral flap ( Fig. 24 View FIGURE 21 – 27 ) with a maximum of 25 laterodistal chaetae (19 ciliated and 6 smooth).
Furcula. Manubrial plate with 2 inner and up to 6 outer chaetae, and with 4 pseudopores ( Fig. 25 View FIGURE 21 – 27 ). Ventral manubrial chaetotaxy as in Fig. 26 View FIGURE 21 – 27 .
Mucro typically falcate ( Fig. 27 View FIGURE 21 – 27 ). Ratio manubrium/dens/mucro as 50:71:1.
Variability. Intraspecific and very often bilateral variabilities in chaetotaxy are characteristic for populations found both in Kecskemét (type locality of S. pallidipes ) and in Simontornya (type locality of S. pillichi ). Percentage values presented here are based on 18 adult specimens (case number N=36 due to bilaterality). Macrochaetae number and distribution shows high variability on Th. II region 2 and region 3 (group PmA and PmC), and on Abd. IV region A according to the modified Jacquemart system ( Christiansen & Bellinger 2000). Th. II region 2 is composed of 7 to 10 macrochaetae ( Fig. 28 View FIGURE 28 ). The most frequent number of macrochaetae is 9 (n=23; 64%), while region 2 with 7 macrochatae was observed in only one case (~3%), asymmetrically. Group PmA of Th. II region 3 is composed of 9 to 12 macrochaetae ( Fig. 29 View FIGURE 29 ). PmA group with 10 and 11 macrochaetae were found evenly frequent (n=13; ~36%), group with 9 macrochaetae were observed in 9 cases (25%), while the highest number (12) of macrochaetae was detected asymmetrically in only one specimen (~3%). Group PmC of Th. II region 3 is composed of 8 (n=29; ~72%) or 9 (n=10; ~28%) macrochaetae ( Fig. 29 View FIGURE 29 ). Abd. IV region A is predominantly composed of 8 macrochaetae ( Fig. 30 View FIGURE 30 ). 7 macrochaetae were observed in two cases (both as bilateral variability), with (i) chaeta A3 absent or with (ii) Be2 present as microchaeta instead of macrochaeta. Abd.
IV region A with 9 macrochaetae was found in three cases, always asymmetrically, with an additional macrochaeta of uncertain homology.
Ecology and distribution. The type localities are characterized by closed grasslands (sandy grassland in Kecskemét and loess steppic grassland in Simontornya). Earlier occurrence data are mostly connected to the same habitat types ( Loksa 1983, 1987, Hornung 1986), but also to extremely saline open pioneer swards ( Móczár & Bíró 1980). The newly discovered populations in the Börzsöny Mountain (part of the North Hungarian Mountains) and on the Mount Somló (part of the Transdanubian Mountains) inhabit sub-pannonic steppic grasslands on stony soils, which often share a relatively common Collembola fauna with sandy or loess steppic grasslands. Outside Hungary, S. pallidipes is known from Serbia occurring in sandy grasslands ( Loksa & Bogojević 1970), and from Austria from a mid-mountain dry grassland ( Kampichler 1991). A map summarizing the distribution of S. pallidipes is shown in Fig. 42 View FIGURE 42 .
Based on the known distribution and habitat associations, this species can be considered xerothermophilous, and, concerning soil characteristics, psammophilous, lithophilous and halotolerant as well.










| HNHM |
Hungarian Natural History Museum (Termeszettudomanyi Muzeum) |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Seira pallidipes Reuter, 1895
| Dányi, László 2017 |
Seira pallipes
| Hornung 1986: 138 |
Seira pillichi
| SZeptycki 1979: 100 |
| Loksa 1970: 135 |
| Ellis 1966: 142 |
Seira pallidipes
| Kampichler 1991: 150 |
| Loksa 1987: 79 |
| Loksa 1983: 74 |
| MocZar 1980: 153 |
| SZeptycki 1979: 41 |
| Ellis 1976: 316 |
| Loksa 1970: 133 |
| Palissa 1964: 220 |
| Gisin 1960: 237 |
Seira pillichi
| Palissa 1964: 218 |
| Gisin 1960: 237 |
Lepidocyrtinus pillichi
| Denis 1941: 45 |
Pseudosira (sic!) pillichi
| Gisin 1944: 83 |
| Stach 1930: 299 |
(sic!) pallidipes
| Gisin 1944: 81 |
| Denis 1941: 45 |
| Reuter 1895: 114 |