
STREPSIPTERA
|
publication ID |
https://doi.org/10.1111/j.1096-3642.2007.00314.x |
|
persistent identifier |
https://treatment.plazi.org/id/0E458F02-FF8C-B253-FE6B-F971FB58FEB0 |
|
treatment provided by |
Felipe |
|
scientific name |
STREPSIPTERA |
| status |
|
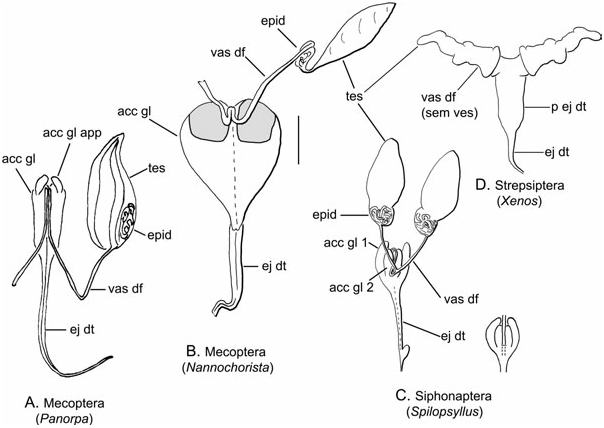
Description ( Fig. 1D View Figure 1 )
Testis: Each testis consists of an anteriorly rounded, compact lobe ( Hünefeld & Beutel, 2005) and with little external differentiation from the following vas deferens.
Epididymis: Not differentiated.
Vas deferens: These ducts are weakly distinguished externally from the testes, usually separated from each other (e.g. Xenos sp. , Mengenilla sp. ; Carcupino et al., 1995; Hünefeld & Beutel, 2005) or they can be fused (e.g. Corioxenos sp. ; Matsuda, 1976).
Accessory gland and seminal vesicle: No accessory glands are identified. The seminal vesicles are considered modified vasa deferentia and can be quite asymmetrical ( Hünefeld & Beutel, 2005).
Ejaculatory duct: The duct is unpaired and its proximal portion functions as a sperm pump ( Hünefeld & Beutel, 2005).
Ejaculatory apodeme, sperm pump, and aedeagus: The sperm pump is positioned at the anterior end of the ejaculatory duct ( Matsuda, 1976) as opposed to the posterior end as in Mecoptera and Diptera . A sclerotized ejaculatory apodeme is absent and the pump is simply a muscular chamber, comprising an internal ring of circular muscle and an outer sheath of longitudinal muscle. The aedeagus is tubular or needle-like with a narrow hooked tip. No parameres are present. The spermatozoa are pumped through the narrow aedeagus and no spermatophore is probably formed ( Hünefeld & Beutel, 2005).
Remarks: The reproductive tract is greatly reduced and modified in Strepsiptera . The seminal vesicle is in the same position as the epididymis in Mecoptera and is perhaps homologous. The sperm pump is clearly not homologous to that of Diptera , lacking sclerotized parts and derived from the proximal portion of the ejaculatory duct ( Hünefeld & Beutel, 2005). The ejaculatory duct is normally encircled by circular muscle, but the presence of longitudinal muscle is possibly unique to Strepsiptera .
This order was included in the present outgroup survey because of the recent views concerning the relationships of Strepsiptera and recognition of the Halteria [e.g. Whiting & Wheeler, 1994; Whiting et al., 1997; Whiting, 1998a, b, 2002, 2005; but see Rokas, Kathirithamby & Holland (1999) and Kukalová-Peck (1998) for evidence opposing this relationship]. Kristensen (1999) reviewed some of the morphological evidence for the Strepsiptera –Antliophora relationship and called into question most of the supporting characters either as regressive, problematical or possibly homoplastic. He concluded that the morphological evidence supporting the sister grouping Strepsiptera + Diptera was ambiguous. In fact, eight of the ten characters listed by Whiting (1998b) supporting Strepsiptera’s inclusion in the Antliophora are absent in Strepsiptera or inapplicable (characters 18–27; only 25 and 27 are present, with 26 a reversal). New information on the ground plan structure of the adult Strepsipteran mandible (character 18) shows that it is clearly chewing and not a dagger or blade-like structure as present in Antliophora ( Grimaldi, Kathirithamby & Schawaroch, 2005; Pohl, Beutel & Kinzelbach, 2005). A further purported synapomorphy proposed by Whiting (1998b; character 25) is a ring-like segment 9 of the male abdomen. However, this feature is also present in the ground plans of male Trichoptera and Lepidoptera ( Wood, 1991) . The anterior position of the sperm pump and the absence of an inner ejaculatory apodeme make it unlikely that the pump (character 27) in Strepsiptera is homologous to that of the Antliophora, a conclusion also reached by Hünefeld & Beutel (2005). We agree with Kristensen (1999) that the proposed Strepsiptera /Antliophoran clade can be refuted on morphological grounds. The reduced nature of the strepsipteran male gonads, especially the absence of accessory glands, provides no additional evidence supporting such a relationship.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.