
Apostolepis collaris, Peracca, 1897
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4392.3.4 |
|
publication LSID |
lsid:zoobank.org:pub:8621C6F2-C541-4771-A813-3034CC3E352D |
|
DOI |
https://doi.org/10.5281/zenodo.5963618 |
|
persistent identifier |
https://treatment.plazi.org/id/0F7C87F4-FFA0-DD06-9186-6BB15E889B92 |
|
treatment provided by |
Plazi |
|
scientific name |
Apostolepis collaris |
| status |
|
Atractus collaris Peracca, 1897
Figures 2 View FIGURE 2 , 3A–C View FIGURE 3 , 4–6 View FIGURE4 View FIGURE5 View FIGURE 6
Atractus collaris PERACCA 1897 : 4 (CONONACO RIVER, PASTAZA, ECUADOR). Leptocalamus limitaneus AMARAL 1935: 219 (LA PEDRERA, AMAZONAS, COLOMBIA; IBSP 9196). New synonymy. [Atractus] limitaneus —SAVAGE 1960: 81.
Atractus collaris collaris— DIxON & SOINI 1977: 34 (IN PART).
Atractus alphonsehogei— MARTINS & OLIVEIRA 1993: 24 (FMT 450, A MALE SPECIMEN). Atractus collaris —CARRILLO & ICOCHEA 1995: 13.
Holotype. Adult male housed in the Museo Regionali di Scienza Naturali di Torino ( MRSNT) from Cononaco River (c.a., 0 1 ° 31’S, 75 ° 36’W; 190m asl), province of Pastaza, Ecuador. Specimen apparently lost (see Andreaone & Gavetti 2007) ( Fig. 4 View FIGURE4 ). GoogleMaps
Diagnosis. Atractus collaris is distinguished from all congeners, except for those species of the A. collaris species group by having one (usually) or two (rarely) apical pits on dorsal scales from both sexes and supracloacal tubercles in the cloacal region of mature males (Passos et al. 2013b). Additionally, the following combination of morphological characters is unique of the species and distinguishes it from any other species of Atractus: (1) dorsal scale rows 17/17/17 with apical pits in both sexes and supracloacal tubercles in males; (2) postoculars two; (3) moderately long loreal, contacting first three supralabials; (4) temporals 1+2; (5) seven supralabials, third and fourth contacting eye; (6) seven infralabials, first three contacting chinshields; (7) five (rarely) or six maxillary teeth; (8) gular scale rows in four series; (9) preventrals usually four; (10) ventrals 167–186 in females, 145–178 in males; (11) subcaudals 18–24 in females, 22–33 in males; (12) in preservative, dorsum brown to grayish black, with cream occipital collar incomplete and small paired black spots usually cream bordered along the body, first dorsal scale rows with lighter center and brown lateral lines; (13) in preservative, venter cream except for two lines (one from each side of belly) in the lateral margins of ventral scales (paraventral region); (14) small body size, females reaching 300 mm SVL, males 218 mm; (15) moderately tail length in females (8.2–10.9.3% SVL) and males (10.9–14.5% SVL); (16) hemipenis moderately bilobed, non-capitate, and non-calyculate.
Comparisons. Atractus collaris differs from all members of the A. collaris species group, except A. alphonsenhogei and A. gaigeae , in having first supralabial contacting loreal (vs. first supralabial not contacting loreal in A. caxiuana , A. hoogmoedi , A. surucucu , and A. zidoki ). Atractus collaris differs from A. alphonsehogei in having descalcified alary spines and hemipenial lobes centrolineally oriented, dorsum with conspicuous spots, and yellow supralabials (vs. calcified alary spines and hemipenial lobes centrifugally oriented, dorsum uniformly dark brown or black lacking spots, and cream supralabials; from A. gaigeae in having 167–186 ventrals in females, 146–178 in males and lacking the vertebral line (vs. 200–214 ventrals in females, 184–198 in males).
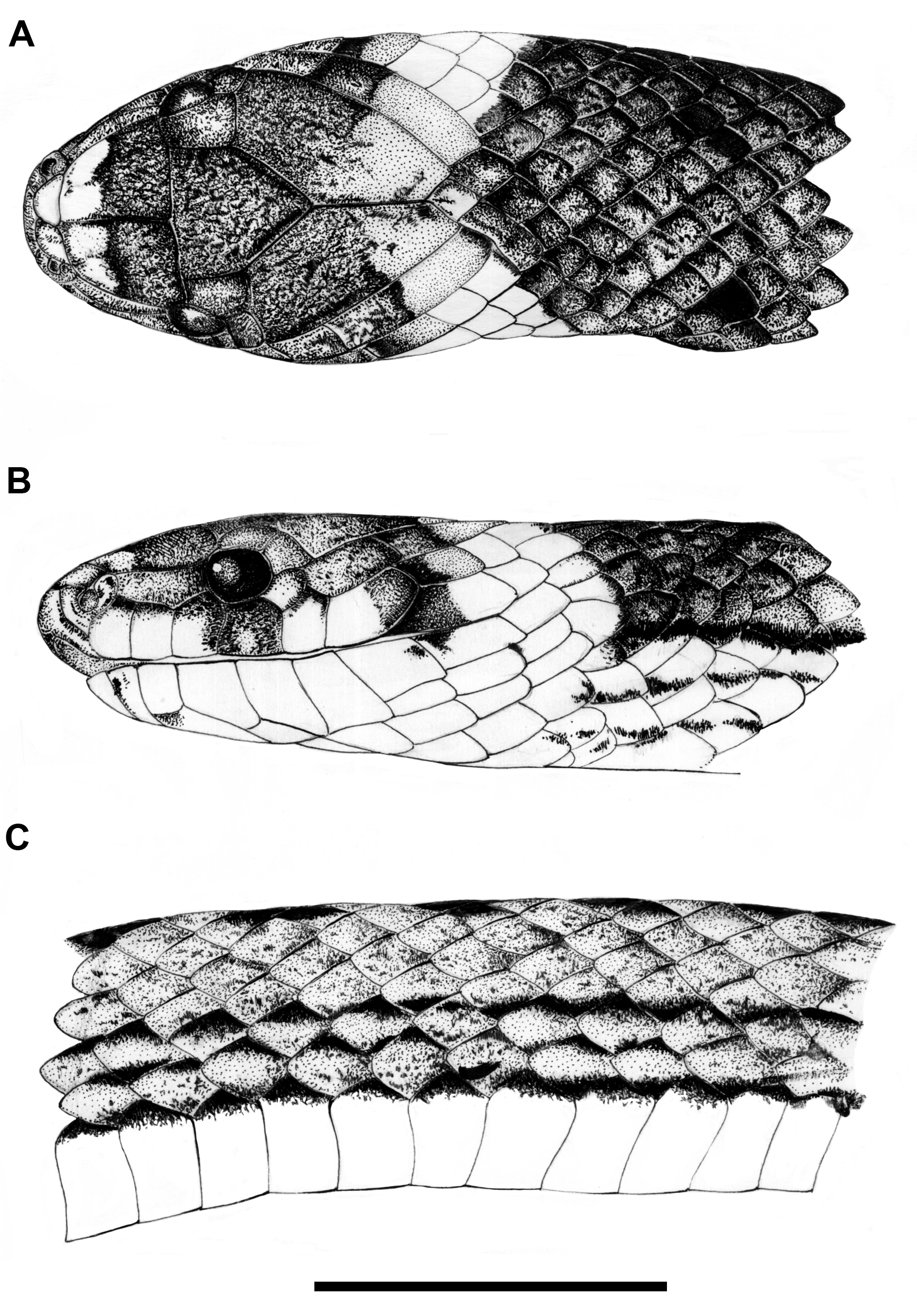
Description. Head slightly distinct from body, twice as long as wide, and arched in lateral view; snout rounded in dorsal view and truncated in lateral view; rostrum-orbit distance about half of head length; nostril-orbit distance as long as prefrontal length; interorbital distance as lon as parietal length; rostral subpyramidal in frontal view, wider than high, and barely visible in dorsal view; internasals as wide as long; internasal suture sinistral with respect to prefrontal suture; prefrontal as wide as long; supraocular subtrapezoidal in dorsal view, about three times as long as wide; frontal subpyramidal, as long as wide; parietal about twice as long as wide; nasal divided; nostril between prenasal and postnasal; prenasal about twice as high as long; postnasal shorter than prenasal, as high as long; loreal long, contacting first three supralabials; pupil round; two postoculars similar in height; upper postocular slightly longer than lower postocular; temporals 1+2; first temporal about three times as long as high; upper posterior temporals usually fused in a single shield, about five times as long as wide; supralabials seven, third and fourth contacting eye; first four supralabials nearly equal in height; sixth supralabial taller and seventh supralabial longer than remaining supralabials; symphysial semicircular, about three times as wide as long; fist pair of infralabials preventing symphysial/chinshields contact; infralabials seven, first three contacting chinshields; chinshields three times as long as wide; gular scales in four series; preventrals usually four; dorsal scale rows 17/ 17/17; dorsal scales usually with one apical pit (rarely two) on both sexes and supracloacal tubercles in mature males; terminal spine large (longer than last subcaudal), approximately robust and conical.
Maxilla arched in dorsal view with usually with four prediastemal teeth and one postdiastemal tooth; first two teeth moderately spaced (smaller than tooth size, measure taken from the base of the tooth to its apex); spaces among each one of third and fourth teeth longer (similar to the tooth size); prediastemal teeth, posteriorly curved, angulars, robust at base and narrowed in the apices; teeth decreasing posteriorly in size; diastema longer than size of postdiastemal tooth; postdiastemal tooth half-sized of predistemal ones; lateral maxillary process little developed lacking posterior projection.
Color pattern variation in preservative. Dorsum of head dark brown to black with cream spots usually covering snout region (rostral, internasals and anterior portion of prefrontals) and displaying an incomplete occipital collar; incomplete occipital collar extending from posterior area of parietals to lower temporal region, frequently connected with wide cream gular region; lateral surface of head brown to dorsal edges of supralabials; supralabials cream except for dorsal area of third and fourth supralabials mostly dark brown; eventually fourth supralabial entirely dark brown; posterior region of sixth and fully seventh supralabials usually dark brown; infralabials, chinshields, gular region and preventrals usually uniformly cream; occasionally, suture region among infralabials, symphysial, chinshields and other scales from mental region with dispersed and irregular dark brown dots or spots; ventral surface of body and tail uniformly cream, except for dark brown to black lateral margins of ventral scales; lateral pigmentation of ventrals longitudinally arranged forming paraventral lines; occasionally, ventral surface of tail with irregular brown dots or spots; dorsal background of body brown to dark brown, usually with a series of small paired black spots (one scale long and one to two scale wide) along paravertebral region (seventh to eighth scales rows) and continuous dorsolateral black stripes covering dorsal edges of first two or three scale rows; more rarely, individuals displaying a conspicuous vertebral stripe; paired black spots edged anteriorly, laterally (sometimes) and posteriorly by light (pale brown or beige) spots (one scale long and wide); stripes usually located on the dorsal edges of first three scale rows but sometimes restricted to first two scale row; lower half of first two scales rows sometimes light pigmented given an impression of a light stripe due to the contrast with the dark brown dorsolateral stripes; central portion of first dorsal scale row usually cream pigmented contrasting sharply with paraventral and dorsolateral dark lines on the first three scale rows. Paravertebral spots and dorsolateral lines are usually barely distinct from brown background in old preserved or melanistic specimens.
Color pattern variation in life. Dorsum of head dark brown with cream to orange spots covering snout region; eye with red to reddish brown iris; center of supralabials and infralabials cream to yellow pigmented; incomplete occipital collar cream to creamish red; belly salmon to red colored, except for lateral margins of ventral scales black pigmented; ventral surface of tail salmon to red with dispersed black dots or spots, sometimes concentrated on the scales sutures; dorsal background of body red to dark brown, with black spots and stripes; paravertebral spots bordered by cream to creamish red; light dorsolateral stripes cream to beige ( Fig. 3A–C View FIGURE 3 ).
Microdermatoglyphics. Apical borders of basal portion cells 2–4 µm apart from each other; borders of denticulations slightly triangular not exceeding 1 µm high; cells surface composed by small and barely conspicuous pores; at midapical portion, spinules from cell borders longitudinally narrow with 1.5–2.0 µm height. Microdermatoglyphic pattern at this portion of the scale is, essentially, the same for the A. alphonsehogei and A. gaigeae ( Fig. 9A View FIGURE 9 ).
Hemipenis morphology. Retracted organ extends at level of seventh to eighth subcaudal and bifurcates at level of sixth to seventh subcaudal (n = 2). Fully everted and almost maximally expanded hemipenis (n = 1, MNRJ 24959) renders a moderately bilobed, non-capitate and non-calyculate; lobes barely centrifugally oriented distally and slightly distinct from hemipenial body; distal portion of lobes clavate with nearly attenuated apices; lobular region is not distinct from the hemipenial body, both in ornamentation and by lacking the capitular groove; lobes and hemipenial body uniformly covered by alary spines (sensu Passos et al. 2013d), except for lobular portion on the asulcate side with scarce and spaced spines; alary spines with reduced size above lobular region, forming irregular barely defined flounces at distal portion of hemipenial body on the asulcate side; alary spines spaced widely at their bases on the intrasulcar region and at medial portion of hemipenial body; sulcus spermaticus bifurcates on the proximal third of organ; each sulcus spermaticus branch with centrolinear orientation, extending to lobe apices; sulcus spermaticus moderately laterally expanded before division and narrowed after bifurcation; margins bordered lacking spinules along almost all entire extension; basal naked pocket indistinct; basal region of hemipenis with longitudinal plicae and small spinules irregularly distributed ( Fig. 10A View FIGURE 10 ).
Meristic and morphometric variation. Largest male SVL 218 mm, TL 26 mm; largest female SVL 300 mm, TL 25 mm; tail 10.9–14.2% SVL (mean = 12.3; SD = 0.4; n = 21) in males, 8.2–10.9% SVL (mean = 9.0; SD = 1.0; n = 6) in females; ventrals 145–178 (mean = 158.7; SD = 10.6; n = 21) in males, 167–186 (mean = 176.2; SD = 7.2; n = 9) in females; subcaudals 22–35 (mean = 27.5; SD = 3.1; n = 21) in males, 20–26 (mean = 22.5; SD = 2.4; n = 8) in females; dorsals at the level of second subcaudal 7–9 (mean = 8.5; SD = 0.6; n = 60 sides); midbody diameter 2.0– 4.4 mm (mean = 3.6; SD = 0.8; n = 9); maxillary teeth 5 (n = 8 sides) or 6 (n = 28 sides).
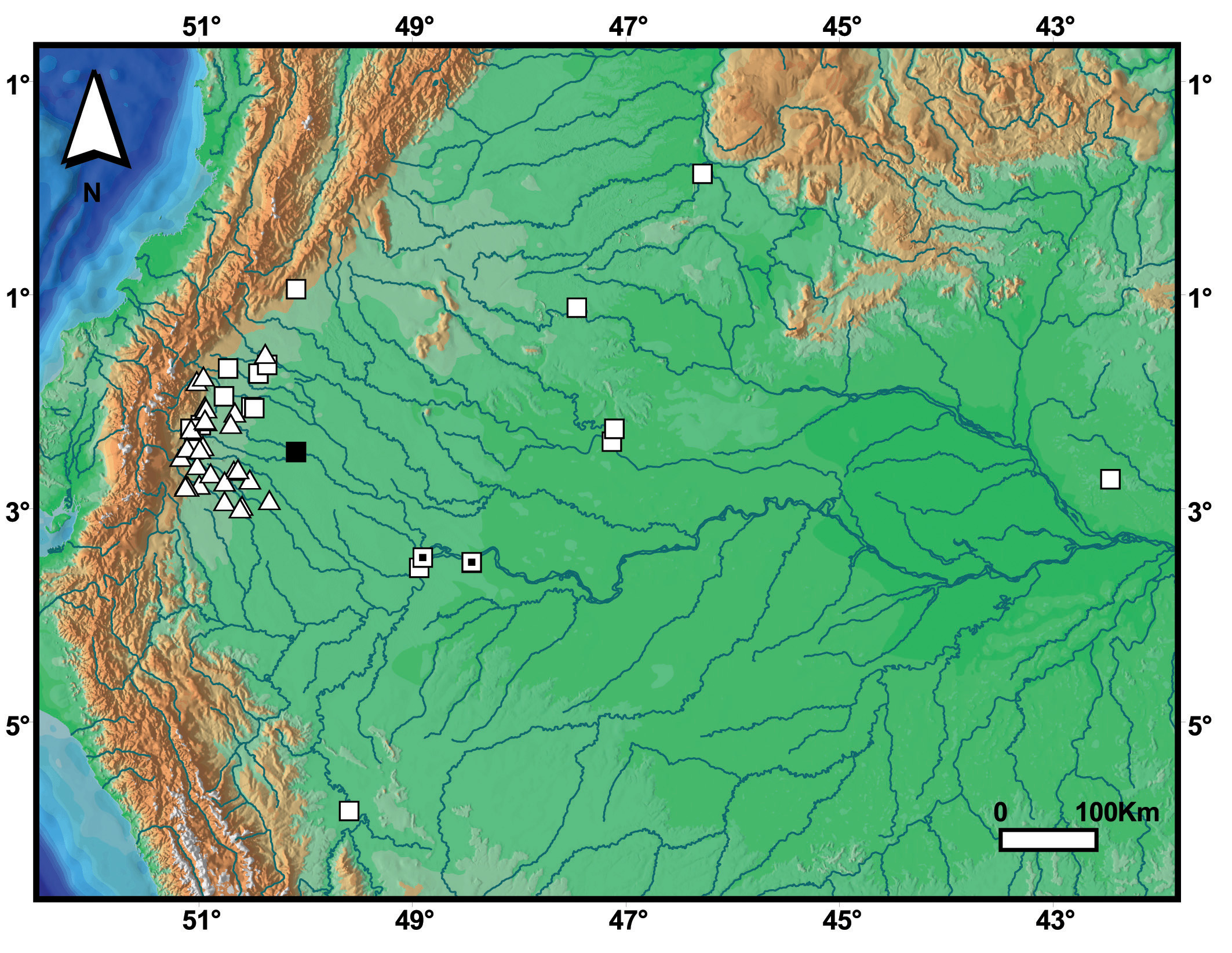
Distribution. Amazon rainforest eastern of the Andes from Inírida (03 ° 48’N, 67 ° 50’W), department of Guainía, Colombia, south to Coronel Portillo (08 ° 23’S, 74 ° 35’W) department of Ucayali, Peru; and east to Urucú River (ca. 0 4 ° 31’S, 64 ° 23’W), state of Amazonas, Brazil. Atractus collaris occurs from 100–700 m elevation ( Fig. 12 View FIGURE 12 ). Martins & Oliveira (1993) report a specimen of A. alphonsehogei from Presidente Figueiredo (01 ° 26’S, 60 ° 01’W), state of Amazonas, Brazil. Although this specimen (a male, FMT 405) was not available to us, we have examined other specimen from Presidente Prudente (MPEG 17545) and confirmed its identification as Atractus collaris . Therefore, based on the data available in the Martins & Oliveira (1993) and also on the above species record, we tentatively consider FMT 405 as A. collaris .
Remarks. Recently, Wallach et al. (2014) pointed out that the holotype of Leptocalamus limitaneus— as well as many other types of Neotropical snakes taxa — was lost in the tragic fire accident that consumed most of the herpetological collection of the Instituto Butantan (see Kumar 2010; Warrel et al. 2010; Franco 2012). However, this and other types of Atractus remain intact and available to examination because such specimens were loaned to the senior author when the accident happened. In addition, we emphasize that other type specimens kept on the metallic cabinets survived (with some degree of damage) from the fire due to oxygen consumption inside the compacted racks (P. Passos pers. observ.). Therefore, authors of future studies must make sure to consult the curator in charge of the Instituto Butantan collection regarding the whereabouts of the types of interest for comparisons before proposing neotype designations based on the provisions of ICZN (1999).






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |