
Axinella nayaritensis, Carballo & Bautista-Guerrero & Cruz-Barraza, 2018
|
publication ID |
https://doi.org/10.11646/zootaxa.4482.1.4 |
|
publication LSID |
lsid:zoobank.org:pub:2193C3BE-6D10-4DAE-9A06-85B875D28E6D |
|
DOI |
https://doi.org/10.5281/zenodo.5979224 |
|
persistent identifier |
https://treatment.plazi.org/id/11792157-B318-714C-8897-21439097FE23 |
|
treatment provided by |
Plazi |
|
scientific name |
Axinella nayaritensis |
| status |
sp. nov. |
Axinella nayaritensis View in CoL n. sp.
Etymology. The species is named after the state of Nayarit (Mexican Pacific), the first area where the species was found. Nayarit was the name of one of the most famous King of the Cora people that lived in this state in the past. Nayarit means that is ‘between the sun and the sky’.
Material examined: Holotype: LEB-ICML-UNAM-61, Las Monas, isla Isabel ( Nayarit), 21°50’59’’N, 105°52’46’’W, 24 m depth, 11/21/1999 GoogleMaps , Paratypes: LEB-ICML-UNAM-581, Tecuchitán ( Nayarit), 20°43’53’’N, 105°24’44’’W, 25 m depth, 10/05/2002 GoogleMaps . LEB-ICML-UNAM-866, Chacala ( Nayarit), 21°09’57’’N, 105°13’38’’W, 7 m depth, 06/12/2003 GoogleMaps . LEB-ICML-UNAM-1003, Cerro Pelón, isla Isabel ( Nayarit), 21°51’21’’N, 105°53’33’’W, 21 m depth, 10/12/2003 GoogleMaps . LEB-ICML-UNAM-1095, El Bajo ( Oaxaca), 15°40’16’’N, 96°28’11’’W, 7 m depth, 07/ 12/2003 GoogleMaps .
Description: The holotype is an erect branched sponge, with numerous cylindrical branches in different planes and fused for most of their length ( Fig. 2A View FIGURE 2 ). It measures 17 cm high x 18 cm wide (from side to side). Young specimens can vary from simply branched, or only bifurcated at the end, to individuals with several branches mostly fused, or almost coalescent for most of their length ( Fig. 2B,C,D View FIGURE 2 ). It has been also observed specimens up to 25 cm in high with only three or four erect branches. The branches are not flexible, and they measure from 0.5 to 1.8 cm in diameter. They are generally tapering and fused for part of their length. Stem is cylindrical, of the same diameter as the branches or slightly wider, and sometimes expanded below as a woody base. Aquiferous system is superficial and conspicuous, formed by a system of surface canals and meandering ridges which run around the branches ( Fig. 3A,B View FIGURE 3 ). A dermal membrane covers the surface of the sponges, where oscules, slightly elevated are visible (see arrows in Fig. 2A View FIGURE 2 ). The sponge surface is irregularly convoluted and corrugated ( Fig. 3A,B View FIGURE 3 ). The consistency is stiff and very difficult to tear. The colour in life is orange, turning brown in alcohol. It characteristically dyes dark orange in alcohol.
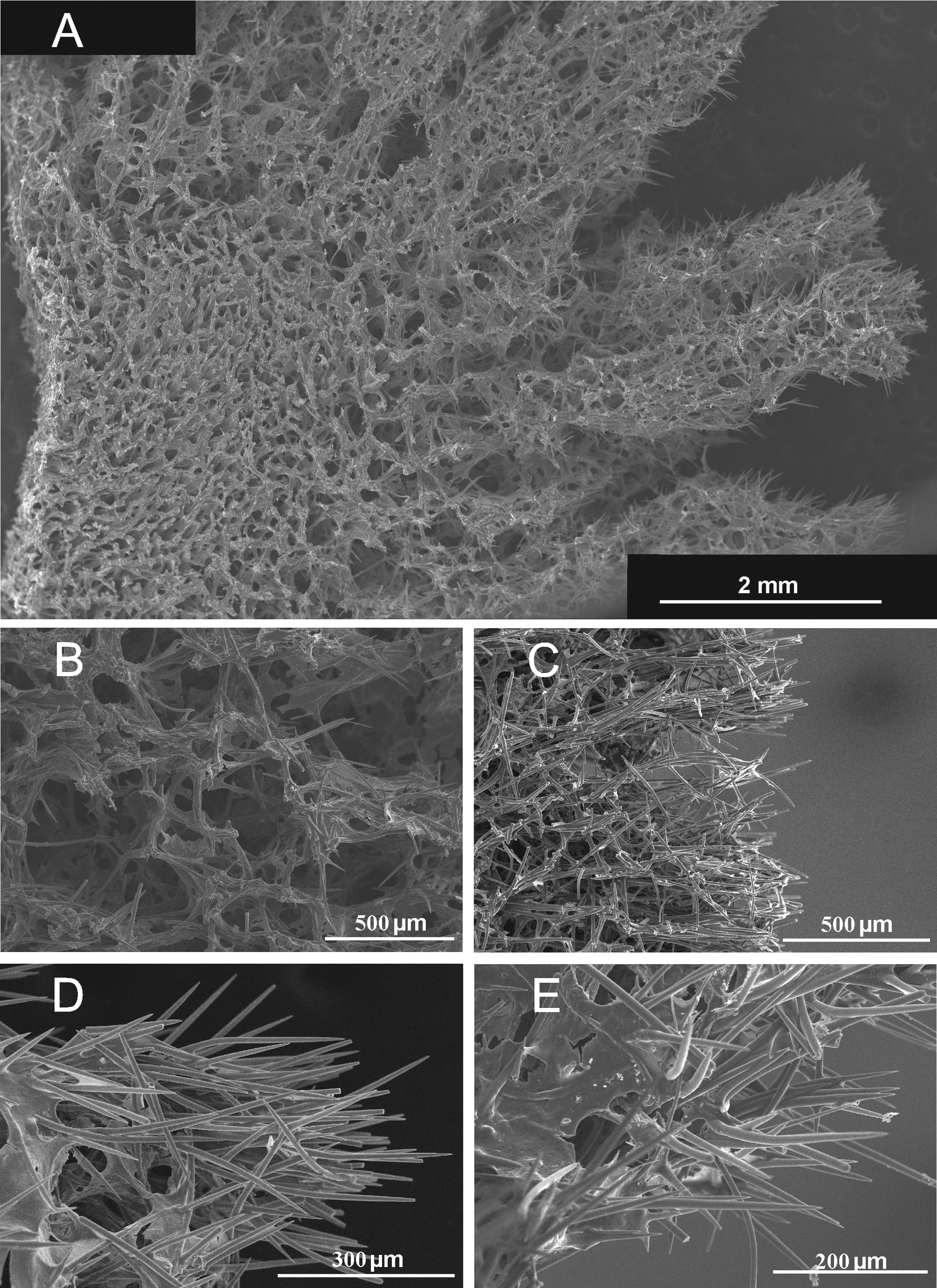
Skeleton: In cross section is observed a dense central axial condensation which forms a reticulated skeleton from which radiates to the surface an extra axial plumoreticulated skeleton ( Fig. 4A View FIGURE 4 ). The extra axial skeleton is interconnected by scattered spicules or fibers in a reticulated pattern ( Fig. 4B View FIGURE 4 ). Extra axial megascleres usually protrude through the surface where sometimes they form fan-shaped spicule brushes, whose tips also project beyond the surface ( Fig. 4D,E View FIGURE 4 ).
The spicules are oxeas (from 185 to 590 µm in length); centrally flexed, and thin or stout styles (from 175 to 475 µm in length), sometimes curved in the proximal third. Rarely with both rounded ends ( Fig. 5 View FIGURE 5 , Fig. 6 View FIGURE 6 , Table 1).
Molecular results: The phylogenetic analysis for 18S rRNA gen was reconstructed with 38 sequences (1627 selected sites, 213 variable and 165 informative) and the CO1 mtDNA gene was reconstructed with 32 sequences (576 selected sites, 256 variable and 233 informative). The ML and BA topology are represented in Figures 7 View FIGURE 7 and 8 View FIGURE 8 . Independently of the phylogenetic method and loci used, the genus Axinella is split into three distinct clades labelled as AXI 1, AXI 2 and AXI 3 as previously recovered by Gazave et al. (2010). Both 18S and COI nested/ clustered near to Axinella nayaritensis n. sp. in AXI 1.
Discussion. Axinella nayaritensis n. sp. is a species typical of the genus in its spiculation, skeletal arrangement, and external growth form. There is no similar species in the eastern Pacific to compare the new species. The geographically closest species is Axinella profunda Ridley & Dendy, 1886 , a deep-sea species collected at abyssal depths from the central Pacific, which is very different in growth form (branches flattened), size (up to 5 cm high), and overall in the form of the megascleres since the latter has styles with minutely spined bases ( Ridley & Dendy 1886).
In the Atlantic coast of north America, the most similar species is Axinella polycapella de Laubenfels, 1953 , widely distributed in the Gulf of Mexico, Florida and east coast of USA. There are also important differences in morphology since A. polycapella is a flexible branched sponge up to 50 cm high, and the new species is smaller and with rigid branches. The colour in A. polycapella varies from orange to red, purple and brown, whereas the new species is always orange. The skeleton is similar in both species; axial skeleton vaguely reticulated to condensed, and extra axial skeleton plumoreticulated, becoming halichondrioid near the surface in A. polycapella . In the new species spicule tracts arise from the inner central portion, ascending and diverging radially towards the surface, forming an extra axial plumoreticulated skeleton. There are also differences in the form of spicules because in addition to oxeas, there are strongyloxeas and styles in A. polycapella (de Laubenfels 1953) .
The systematic position of Axinella nayaritensis n. sp. was confirmed with two independent loci: cytochrome c oxidase subunit 1 gene (CO1 mtDNA) and ribosomal gene (18s rDNA). The 18S region of the new species was 99 % similar (BLAST identity) to Axinella polycapella and Axinella cf. polypoides Schmidt, 1862 . Evidently, the sequence of A. cf. polypoides recorded in GeneBank (accession code EF 092269 View Materials ) and distributed in Indian Island, Australia is not probably a true A. polypoides , which is distributed exclusively from the Mediterranean. Rather, it could be an undescribed species, similar to A. polypoides .
Regarding COI, the most closed species is Axinella aruensis ( Hentschel, 1912) , a species with a very variable form (flabellate, erect, fan-shaped or narrow long and flat digits), beige or pale yellow with, vaguely reticulated to halichondrioid skeleton, characteristics not found in the new species. There are also important differences regarding the spicule composition between the two species, since A. aruensis has oxeas with blunt, pointed or telescoped tips with transitional forms ( e.g. styloids, strongyles), and the true styles are scarce. The different morphotypes of A. aruensis occur sympatrically throughout northern Australia, Indonesia and Papua New Guinea, where it was detected that up to 29 % of intra-genomic polymorphism (ITS) could correspond to hybrid species ( Alvarez et al. 2007).
We also established the phylogenetic relationships of the new species within the family Axinellidae , recovering the same clades obtained by Gazave et al. (2010). The new species was included in the clade AXI 1 (clade named Axinella by Gazave et al. 2010) (for both 18S and COI), together with eight Axinella species: A. polycapella , A. cf polypoides (probably an undescribed species), A. vaceleti , A. polypoides , A. flustra , A. infundibuliformis , A. pyramidata , A. aruensis . All exhibit a great diversity of shapes but the choanosomal skeleton is often differentiated between axial and extra-axial regions, which is particularly evident in the arborescent forms, as occur in the species of Axinella of this clade.
In addition to these nine typical arborescent Axinella -species, the same clade also grouped species of Pseudoaxinella Schmidt, 1875 (a junior synonym of Axinella ), Dragmacidon Hallmann, 1917 , and Myrmekioderma Ehlers, 1870 . Dragmacidon (family Axinellidae ) are massive or massive encrusting axinellids with a multispicular plumoreticulate choanosomal skeleton, but undifferentiated between axial and extra-axial regions, which is one of the most important differences with Axinella ( Alvarez & Hooper 2002) . Instead, Mymekioderma is a very different genus that belongs to the Family Heteroxyidae Dendy, 1905 . They are species with a detachable ectosomal skeleton consisting of smaller (acanth-)oxeas in brushes perpendicular or paratangential to the surface, supported by larger choanosomal (acanthoxeas, strongyles or styles forming a slightly compressed halichondrioid reticulation in the central portion of the choanosome and a more cavernous peripheral skeleton of oblique spicule tracts, with wispy trichodragmata of raphides (Hooper 2002).
Further work is required to conveniently interpret these results, but the topology obtained in clade 1 after the inclusion of the new species supports the polyphyly of Axinella , according to the presence in the same clade of species of the genus Myrmekioderma and Dragmacidon , as it had already previously suggested ( Alvarez et al. 2000; Gazave et al. 2010).






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |