
Derallus paranensis Oliva, 1981
|
publication ID |
https://doi.org/ 10.1649/744 |
|
persistent identifier |
https://treatment.plazi.org/id/140C87D2-FFBF-E760-FE30-2A35324FFC2A |
|
treatment provided by |
Tatiana |
|
scientific name |
Derallus paranensis Oliva, 1981 |
| status |
|
Derallus paranensis Oliva, 1981 View in CoL
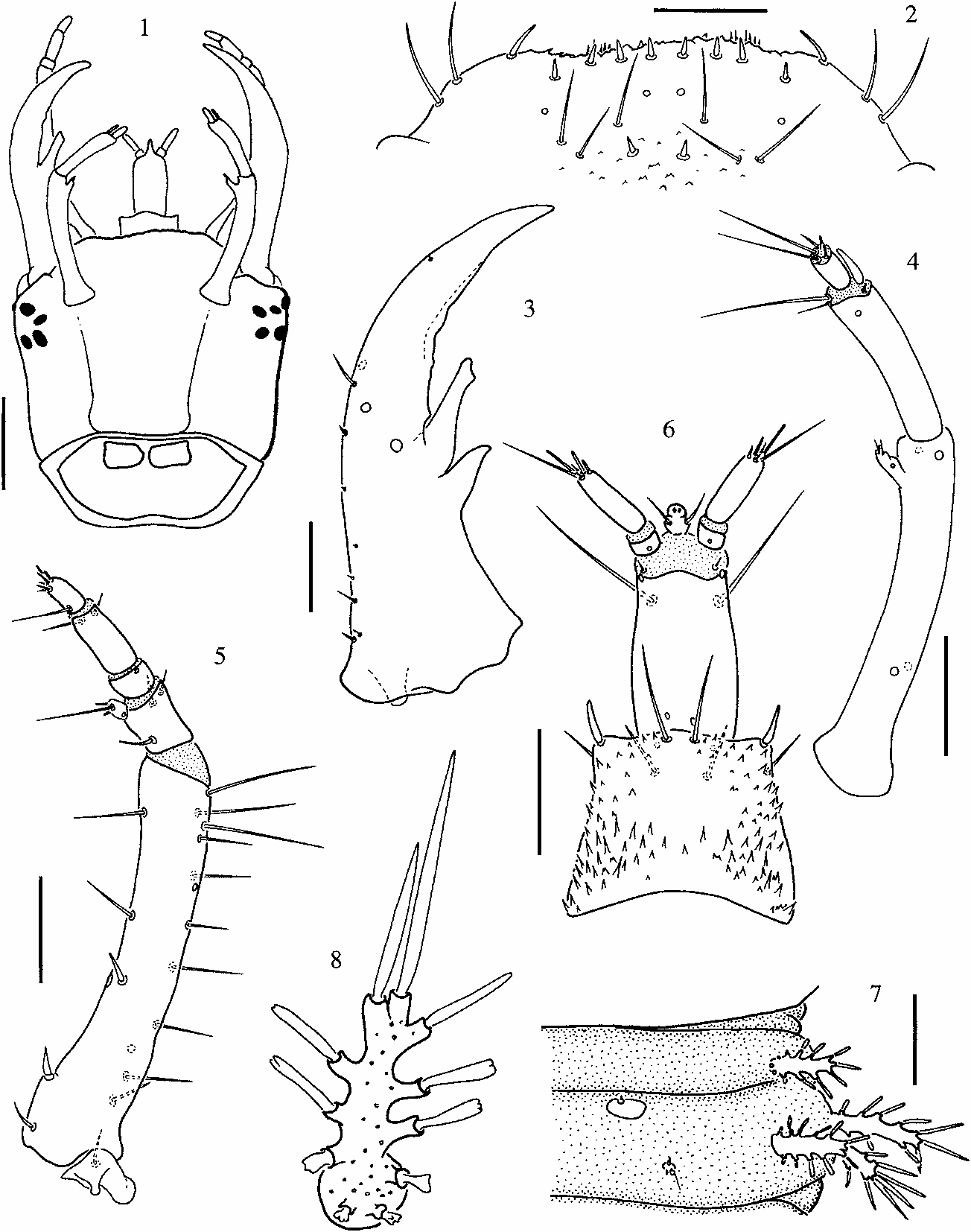
( Figs. 1–9 View Figs View Fig )
Material Examined. ARGENTINA, Buenos Aires province: Partido de Ensenada,
Punta Lara N. von Ellenrieder and L. A. Fernández coll. (44 larvae) .
Description. Egg case. Attached to the lower surface of Azolla filliculoides Lam. leaves. Flat and round similar to the egg case of D. angustus (Archangelsky and Durand 1992) . The only egg case reared in the laboratory contained seven eggs.
Third Instar Larva. Length: 3.8 to 5.3 mm, wider at midlength (between metathorax and second abdominal segment). Color light brown, with sclerotized parts darker; non-sclerotized integument covered by cuticular asperities, denser and rougher on dorsal side.
Head capsule subquadrate ( Fig. 1 View Figs ); occipital foramen wide, dorsal part of cervix with two subquadrate cervical sclerites. Frontoantennal lines parallel, reaching base of head capsule widely separated; coronal line absent (in third instar larvae the ecdysial line is vestigial). Vertex of head with vestiture of small and flat, tooth-like cuticular projections pointing anteriad. Six stemmata on each anterolateral corner of head capsule, close to base of antennae.
Clypeolabrum symmetrical ( Fig. 2 View Figs ). Nasale with several tooth-like cuticular projections and eight short setae; ventral side of nasale with short spicules. Lateral lobes of epistome rounded, not projecting farther than nasale, each with three slender setae.
Mandibles symmetrical ( Fig. 3 View Figs ), with two large inner teeth; distal tooth irregularly shaped, with blunt apex, basal tooth smaller, with sharper apex.
Antennae three-segmented ( Fig. 4 View Figs ); basal segment longer than other two combined, bearing four campaniform sensilla and one inner preapical projection bearing three distal setae and one campaniform sensillum or pore. Second segment slightly slender, with three distal setae, two on inner margin, one on outer margin; distal sensory appendage present on outer apical margin, almost as long as last antennal segment. Third segment the smallest, usually with five apical setae or sensoria, two long and three short ones (in a few cases the number was four or six setae or sensoria).
Maxillae five-segmented ( Fig. 5 View Figs ), longer than antennae; cardo small, irregularly shaped, with long outer seta. Stipes the longest segment, with row of five inner setae; outer margin with ten long setae (in some specimens the number of setae on the outer margin is lower, ranging between seven and nine). Palp four-segmented, second and fourth segments the shortest; basal segment the widest, bearing three slender setae and one short inner process with three distal setae; second segment short, lacking setae; third segment with two subapical setae on ventral side; last segment bearing one long basal seta, projecting mediad, and six short distal setae.
Labium well developed ( Fig. 6 View Figs ). Submentum large, subpentagonal, wider than mentum. Mentum subquadrate, with strong cuticular spines at base and margins; dorsal anterior margin with two pairs of setae, ventral side with two pairs of subapical setae. Prementum rectangular, longer than wide, with two short setae close to base of palps on dorsal side, two long setae on ventral side. Ligula short, easily seen in dorsal view, with two setae at midlength; labial palpi long, twosegmented; basal segment short, bare; second segment long, bearing five apical setae.
Prothorax wider than head capsule; pronotal shield formed by two large plates separated by fine sagittal line; prosternum subpentagonal, with short incomplete sagittal line on basal part. Mesonotum with two pairs of dorsal sclerites, anterior pair small and subtrianguar, posterior pair large, subtriangular; metanotum with two pairs of subrectangular sclerites, anterior pair larger. Pleural areas membranous, those of meso- and metathorax with setiferous projections, three on each pleura. Legs, five-segmented, visible in dorsal view; all three pairs similar in shape, coxae subtriangular, widely separated and elongate, trochanter short, femur as long as coxa but slender, tibiotarsus shorter and slightly slender than femur, pretarsal claw half the length of tibia, with small basal toothlet.
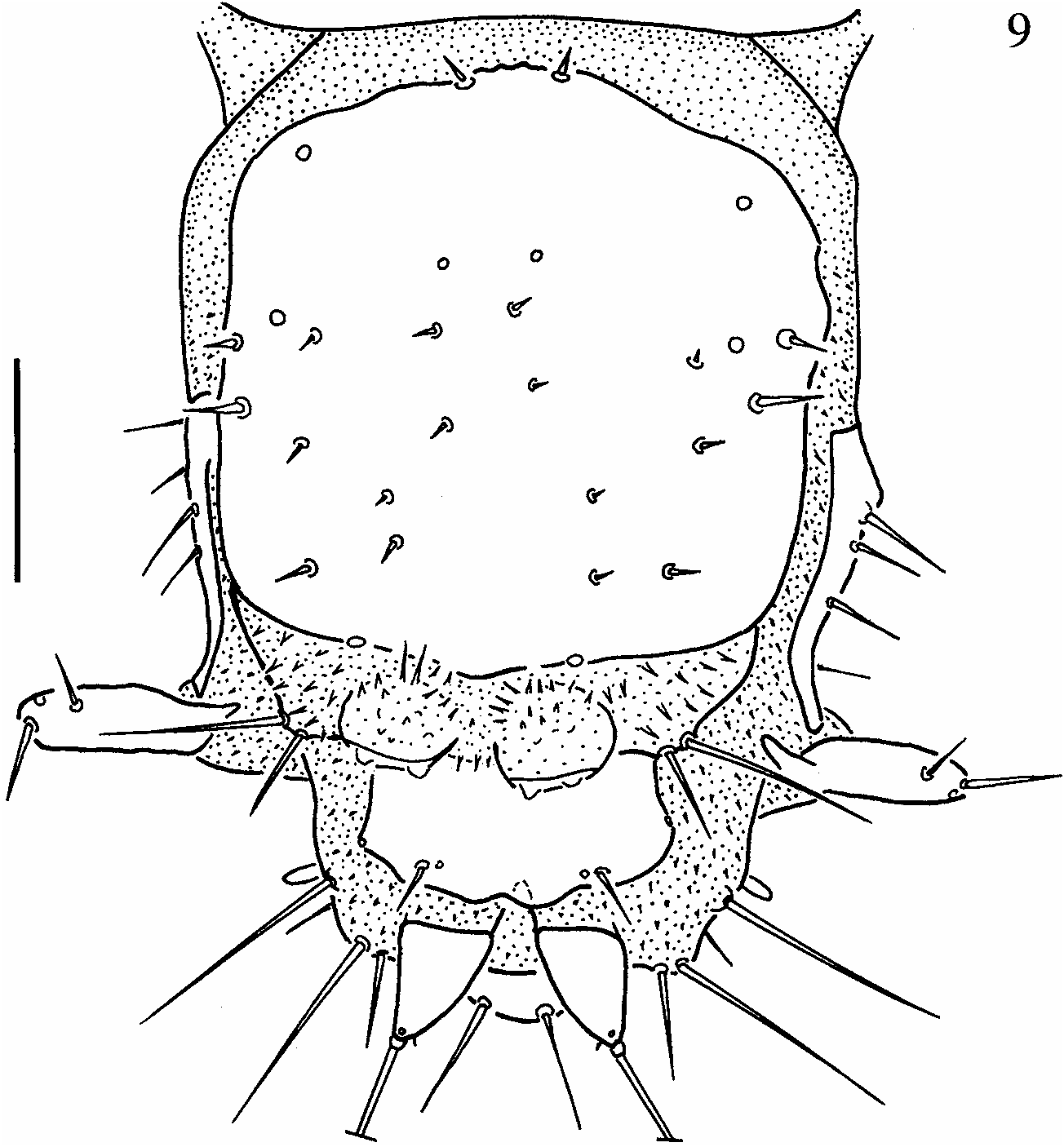
Abdomen ten-segmented, tapering towards posterior end, segments VIII and IX forming spiracular atrium ( Fig. 9 View Fig ), segment X reduced. Segments I to VII similar in size and shape, each with one pair of small sclerites and one small sensorium-bearing plate posterior to sclerite; segments II to VIII subdivided by one transverse fold. Segment I with three pairs of lateral setiferous projections; segments II to VII with four pairs of setiferous projections, one on anterior fold, three on posterior fold ( Figs. 7–8 View Figs ). Segment VIII ( Fig. 9 View Fig ) with large dorsal subquadrangular plate and a pair of short, one-segmented lateral appendages (paracerci); posterior to tergal plate two pairs of tubercles, inner pair spinose, outer pair with two long setae. Segment IX trilobed, partially covered by eighth, with a pair of short, one-segmented, strongly sclerotized urogomphi.
Nine pairs of spiracles, one on mesothorax and eight abdominal. Thoracic and first seven abdominal pairs of spiracles non-functional. Last abdominal pair enclosed within spiracular atrium.
Comparative Notes with First and Second Instar Larvae. First instar larvae show several differences, besides the size ( Table 1), with third instars: 1) nasale without tooth-like cuticular projections; 2) antennae with basal segment shorter than other two
.
combined; 3) mentum with strong cuticular spines, but without the two pairs of setae on anterior margin and the two pairs of subapical setae on ventral side; 4) anterior margin of pronotum without setae; 5) setiferous projections shorter, in relation to body.
Second instar larvae similar to third instar; the main difference between them is the size ( Table 1).
Bionomical Notes. One egg case of D. paranensis was collected and carried to the laboratory. It contained seven eggs; larvae emerged from all of them. They were fed with cladocerans and copepods. Duration of the first instar ranged between 11 and 12 days, that of the second instar between 11 and 17 days; all died as third instars.
Larvae and adults of D. paranensis were collected in three different stations, all three of them covered by floating vegetation composed by Lemna sp. , Wolfiella sp. , Azolla filliculoides Lam. , Hydrocotile ranunculoides L., Alternanthera sp. ; littoral vegetation was represented in one collecting site by Iris pseudacorus L., and in the two others by Scirpus giganteus Kunth. Together with D. paranensis , sporadically two other species of Derallus were present (only as adults): D. argentinensis , common more to the north, in the Paraná Delta, and D. angustus , characteristic of permanent environments of the gallery forest along the shore of the Río de La Plata estuary (usually found on the aquatic fern Salvinia sp. ). The aquatic Coleoptera community at the collecting sites is described in detail in von Ellenrieder and Fernández (2000).
Considering the three sampling stations, adults and larvae were present all year round. First instar larvae were collected between November and March, second instar larvae between November and June, those of third instar were collected all year round. Two of the collecting sites were not permanent (von Ellenrieder and Fernández 2000). One of the collecting sites dried up during the summer of 1997 (between January and April); no adults were collected until October (1997), and larvae did not appear until November of that year. The second sampling station dried up during the months of January and February 1997; adults did not recolonize it until August of that year, and larvae did not appear until November. Finally, the third station dried briefly in January 1997, but the bottom remained moist and larvae were present all year round (although a decay in density was observed during the dry season). In 1998 none of the stations dried up since the ‘‘El Niño’’ phenomenon generated abundant rains all through the summer. In all three sites adults were most numerous between December and March.
Comparative Notes with Other Derallus Larvae, and Larvae of Related
Genera. The larvae of two other Derallus species have been described, D. rudis and D. angustus ( Spangler 1966; Archangelsky and Durand 1992; Archangelsky 1997). There are several differences between the larvae of the three species, mostly related to the head capsule and mouthparts ( Table 2). These differences will allow the identification of the larvae of the three species.
As for differences between Derallus and the related genera Allocotocerus and Regimbartia , very few could be found ( Table 3). These three genera are very distinctive and share many characters that clearly separate them from the other known Berosini genera, Hemiosus and Berosus . Watts (2002) reached a similar conclusion when he described the larvae of the Australian Allocotocerus and Regimbartia . Other researchers, either based on larval or adult morphology, have also concluded that these three genera form a uniform clade, clearly distinct from Berosus and Hemiosus ( Spangler 1966; Bertrand 1972; Oliva 1992).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.