
Monalonion, Herrich-Schaeffer, 1850
|
publication ID |
https://doi.org/10.1111/zoj.12311 |
|
persistent identifier |
https://treatment.plazi.org/id/142A4050-DE0C-FFBC-92EE-E17EFB71FCE0 |
|
treatment provided by |
Marcus |
|
scientific name |
Monalonion |
| status |
|
MONALONION View in CoL View at ENA HERRICH- SCHAEFFER
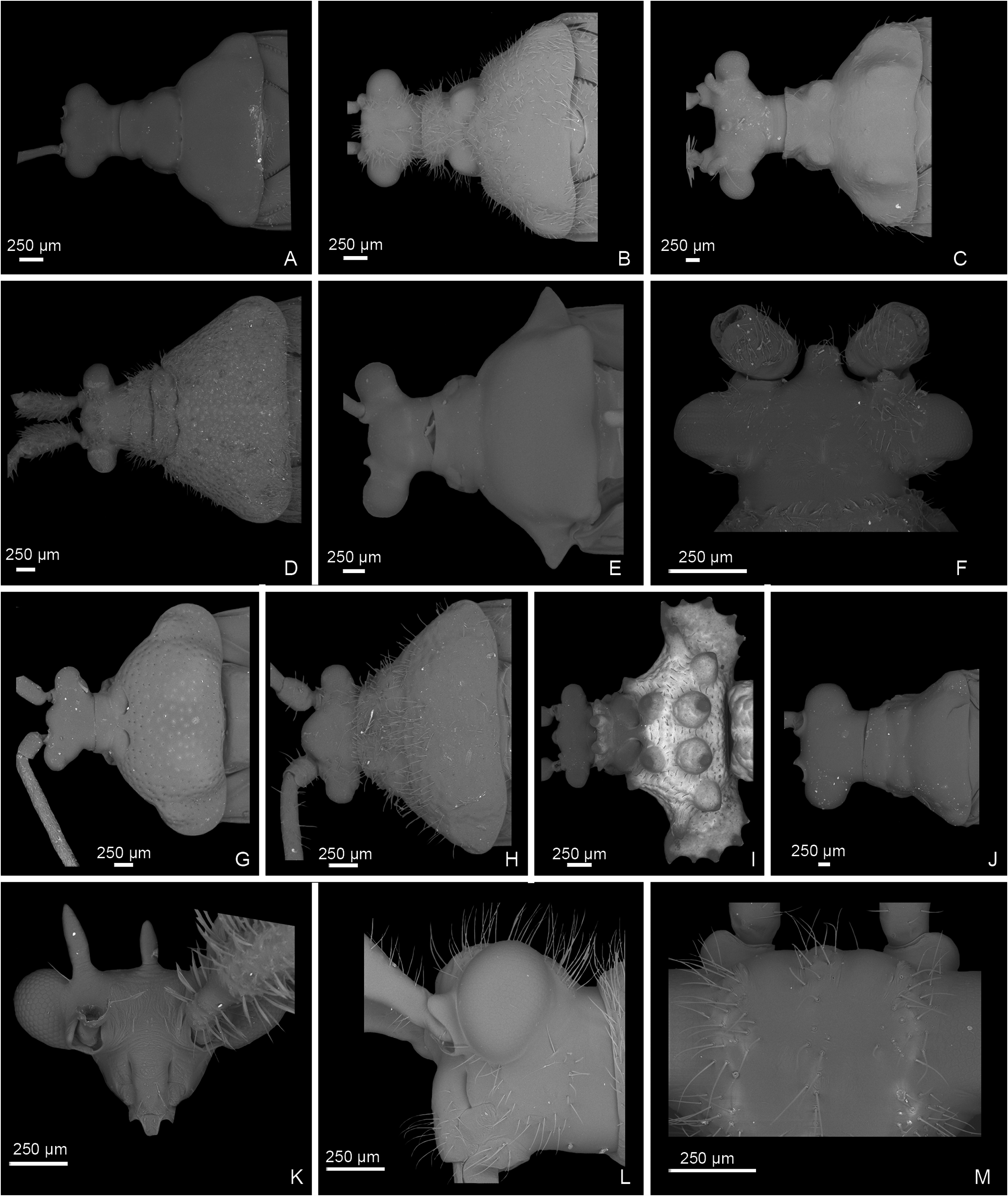
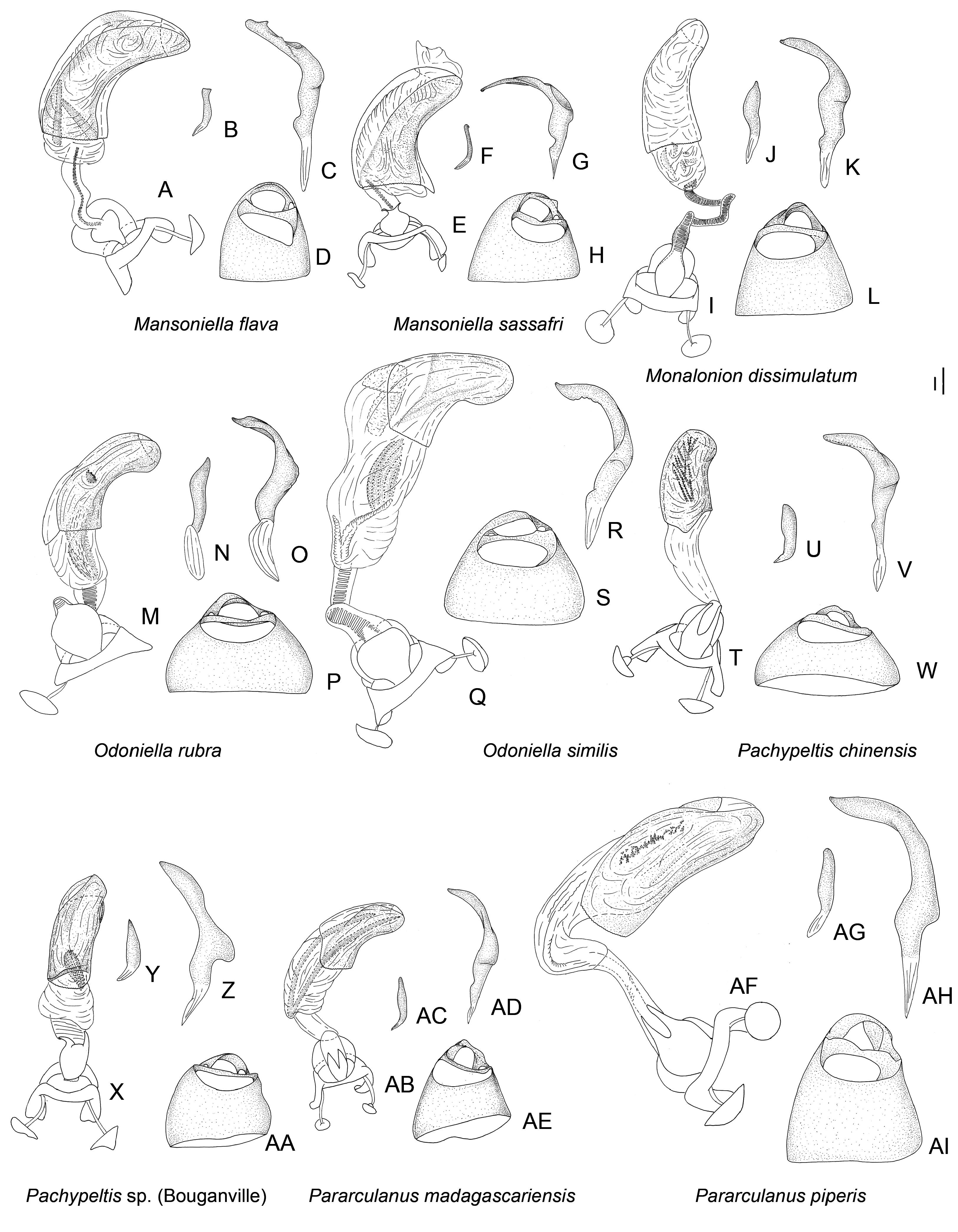
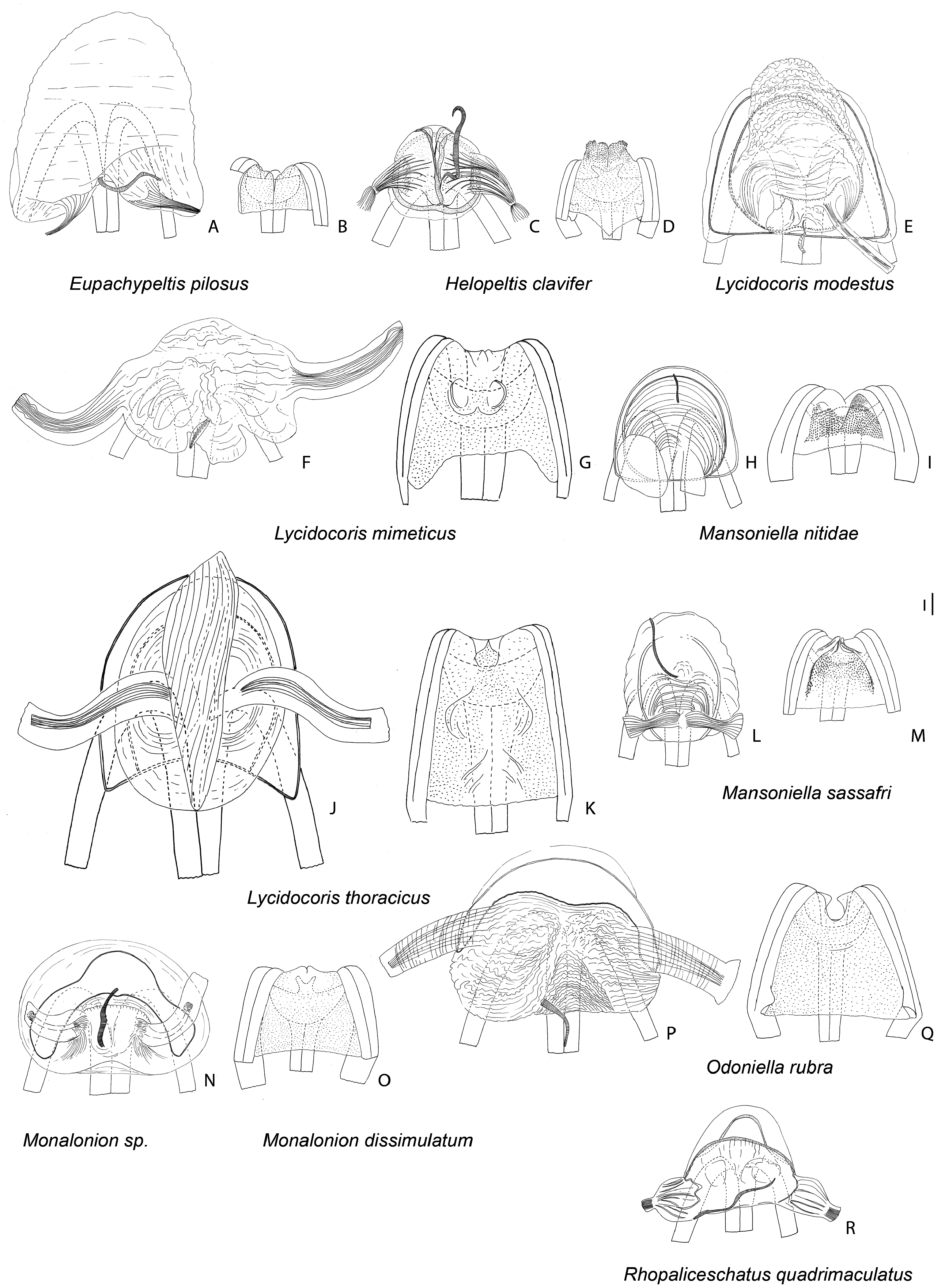
Figures 9 View Figure 9 , 16I–L View Figure 16 , 19N, O View Figure 19 , 24 View Figure 24
Monalonion Herrich-Schaeffer, 1850: 168 View in CoL (gen. nov.; type species Monalonion parviventre Herrich-Schaeffer, 1859 View in CoL by monotypy); Fieber, 1858: 300 (key to gen.); Signoret, 1858: 500 (descr.); Walker, 1873: 161 (cat.); Distant, 1883: 246 (descr.); Atkinson, 1890a: 49 (cat.); Kirkaldy, 1906: 134 (list); Kuhlgatz, 1906: 29 (key to gen.); Reuter, 1908: 150 (descr.); Reuter, 1910: 153 (cat.); Bondar, 1939: 1 (note); Knight, 1939: 226 (note); Carvalho, 1952: 59 (cat.): Carvalho, 1955: 39 (key to gen.); Carvalho, 1957: 138 (cat.); Carvalho, 1972: (diag., key to spp.); Lavabre, 1977a: 65 (diag.); de Abreu, 1977: 85 (desc., distrib., ecol.); Schuh, 1995: 517 (cat.); Schuh, 2002 –2013 (cat.); Namyatova et al., in press (phylogeny).
Diagnosis: Among genera of the Monalonion -group, Monalonion can be separated by: ASI very short, c. 1.5– 3× as long as wide; elongate body ( Fig. 9 View Figure 9 ), ASII–IV filiform; collar distinctly swollen (as in fig. 7D in Namyatova & Cassis, 2013b); scutellum without spine; forecoxae contiguous (as in fig. 17A in Namyatova et al., in press); foretibia almost straight; secondary gonopore surrounded with small sclerite ( Fig. 16I View Figure 16 ); ductus seminis slightly sclerotized basally; and DLP with sclerotized circle ( Fig. 19N View Figure 19 ).
Redescription: Male: Body length 7–12 mm. COLORA- TION ( Fig. 9 View Figure 9 ). Variable, mostly brown to dark brown or yellow, sometimes with reddish areas; antennae and legs usually dark colored, legs sometimes with yellow or pale brown band on femur or mostly yellow. TEXTURE. Body mostly smooth, without punctures, wrinkles and tubercles; vertex without flattened areas; semicircular depression between scutellum and mesoscutum absent; striations on scutellum laterally absent; only small depression on anterior angle of pronotum present (as in fig. 9H in Namyatova & Cassis, 2013b). VESTITURE. Setae on dorsum and thoracic pleura absent; only short simple suberect setae on ASII–IV, legs and abdomen present; setae on ASII sometimes slightly longer than width of hind tibia; setae on tibia and hind femur sometimes very dense, shorter or longer than width of hind tibia; setae on abdomen pale, short and adpressed and very rare; black spinules on femora and tibiae absent. STRUCTURE. Head. Distance between eye and pronotum subequal to half of eye diameter (as in Fig. 10J View Figure 10 ), sometimes subequal to eye diameter; occipital region delimited with distinct depression; longitudinal depression on vertex as long or slightly longer than eye diameter; eyes not stylate, in line with contour of head, c. 0.2–0.3× as long as head width; distance between antennal fossae oval as long as or slightly longer than antennal fossa diameter; frons straight or slightly convex, without ridges, outgrowths or longitudinal depression; anterior view of head c. 1.4– 1.7× as wide as high; eye c. 1.7–2.8× as long as distance between eye and apex of clypeus; antennal fossa oval c. 0.5–0.7× as long as eye height, not raised (as in fig. 6D–F in Namyatova & Cassis, 2013b), inferior margin placed distinctly above inferior margin of eye; position of base of clypeus varying from distinctly below inferior margin of antennal fossa to near halfway of antennal fossa height, not delimited with depression; head swollen in lateral view; length of gula varies from slightly longer to almost twice as long as buccula. Labium. Length varying from reaching middle of mesosternum to slightly surpassing posterior margin metasternum; c. 2–3× as long as wide; LSIII c. 3–4× as long as wide, varying from slightly shorter to distinctly longer than LSI; LSIII c. 2–5× as long as wide, varying from slightly shorter to distinctly longer than LSII; LSV c. 4–8× as long as wide, from slightly longer almost twice as long as long as LSIII. Antenna. Slightly shorter or longer than body; ASI c. 2–3× as long as wide, c. 0.3–0.7× as long as head width, widened basally; ASII c. 4–6× as long as ASI, c. 1.4–2× as long as head and pronotum combined; ASIII c. 0.7–0.9× as long as ASII; ASIV c. 0.2– 0.3× as long as ASIII; ASII–IV filiform. Thorax. Collar distinctly delimited laterally and posteriorly, swollen; calli separated, flat almost indistinct (as in Fig. 10J View Figure 10 ); depression delimiting calli posteriorly absent (as in Fig. 10J View Figure 10 ); humeral angles of pronotum rounded, not dilated (as in Fig. 10J View Figure 10 ); posterior margin of pronotum slightly sinuate (as in Fig. 10J View Figure 10 ); scutellum flat, rounded apically; without outgrowth or medial depression; metepimeron c. 3–4× as high as long, rounded (as in fig. 9C in Namyatova & Cassis, 2013b); metasternum rounded posteriorly, without medial projection on to abdominal segment II (as in fig. 17B in Namyatova et al., in press). Hemelytron. Costal margin concave or almost straight, hemelytron almost not tapering posteriorly; claval commissure c. 2–5× as long as scutellum, concave (as in fig. 11G in Namyatova et al., in press); R + M distinct, reaching posterior margin of corium; medial fracture subparallel to R + M; corium without swelling posteriorly; cuneus c. 3–4× as long as wide, c. 0.7–1.2× as long as pronotum, medial margin distinctly concave (as in fig. 13C in Namyatova et al., in press); membrane cell c. 1.4–2.2× as long as pronotum, rounded or acute apically; auxiliary vein absent; distance between cell and apex of membrane c. 0.2–0.4× as long as cell. Legs. Forecoxae contiguous (as in fig. 17A in Namyatova et al., in press); femora more or less swollen apically (as in fig. 18A in Namyatova et al., in press); forefemur almost straight, middle and hind femora distinctly curved, hind tibia sometimes with medial part widened; foretibia slightly shorter, as long as or slightly longer than head and pronotum combined; tibiae without swellings; segment I of hind tarsus distinctly longer than segment II and subequal to or slightly longer than segment III (as in fig. 19A in Namyatova et al., in press); claw broadly rounded (fig. 10F in Namyatova & Cassis, 2013b); basal tooth on claw triangular, short (as in fig. 10B in Namyatova & Cassis, 2013b). Genitalia ( Fig. 16I–L View Figure 16 ). Genital capsule slightly longer than wide, without outgrowth(s); ventral wall shortened anteriorly; left paramere r-shaped, twice as long as right paramere; phallobase sclerite of primary gonopore suboval, without outgrowths supporting ductus seminis; ductus seminis longer than phallotheca, with coils forming wide tube, sclerotized basally and with narrow sclerite around secondary gonopore, attached to phallobase medially; sclerotized part of phallotheca occupying entire dorsal side, rounded apically, without ridge or outgrowth(s); endosoma with small sclerites or sclerotized areas.
Female: Body length 8–12.5 mm. Coloration, surface, vestiture and structure as in male, generally larger than males and eye usually smaller than in male ( Fig. 9 View Figure 9 ). Genitalia ( Fig. 19N, O View Figure 19 ). DLP with sclerotized ring, sometimes indistinct, mostly without striations or with some medially, especially at base of lateral oviducts; lateral oviducts placed at halfway of or in posterior half of DLP, removed from each other and from lateral margins of DLP; spermathecal gland placed in posterior half, medially; posterior wall of bursa copulatrix with tubercles, without outgrowths or sclerotizations; base of second valvula with bifurcate outgrowth; ventral wall of bursa copulatrix membranous.
Distribution: Widely distributed in Latin America, including Mexico ( Fig. 24 View Figure 24 ).
Host plants: Many species of Monalonion are known to damage cocoa pods ( Distant, 1917; Costa Lima, 1938; Bondar, 1939; Carvalho, 1972; de Abreu, 1977; Entwistle, 1977). Monalonion schaefferi was also recorded from cashew ( Piart, 1977), and M. velenzagi is known from avocado ( Carvalho & Costa, 1988). Species are also known from Ficus sp. , Cercopia adenopsis ( Moraceae ), Hamelia patens (Rubiaceae) and Begonia spp. (Begoniaceae) ( Costa Lima, 1938; Bondar, 1939; Carvalho, 1972; Piart, 1977).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Monalonion
| Namyatova, Anna A. & Cassis, Gerasimos 2016 |
Monalonion
| Lavabre EM 1977: 65 |
| Carvalho JCM 1957: 138 |
| Carvalho JCM 1955: 39 |
| Carvalho JCM 1952: 59 |
| Bondar G 1939: 1 |
| Knight HH 1939: 226 |
| Reuter OM 1910: 153 |
| Reuter OM 1908: 150 |
| Kirkaldy GW 1906: 134 |
| Kuhlgatz T 1906: 29 |
| Atkinson ET 1890: 49 |
| Walker F 1873: 161 |
| Fieber FX 1858: 300 |
| Signoret V 1858: 500 |
| Herrich-Schaeffer GAW 1850: 168 |