
MONALONIINI, Carvalho, 1952
|
publication ID |
https://doi.org/10.1111/zoj.12311 |
|
persistent identifier |
https://treatment.plazi.org/id/142A4050-DE5B-FFD0-91E7-E4EDFC40FF3C |
|
treatment provided by |
Marcus |
|
scientific name |
MONALONIINI |
| status |
|
TRIBE MONALONIINI
Monalonionaria Reuter, 1892: 398 (division nov.);
Eucerocoraria Kirkaldy, 1902: 294 (division nov.); Reuter, 1910: 123 (disc.)
Monaloniaria Reuter, 1910: 123 (disc.);
Odoniellaria Reuter, 1910: 123 (disc.); Oshanin, 1912: 70 (as tribe, cat.)
Monaloniini Carvalho, 1952: 33 , 35, 40, 41, 59 (as tribe, disc., key to tribes, cat.); Carvalho, 1955: 16, 38 (key to tribes, key to gen.); Carvalho, 1957: 131 (cat.); Odhiambo, 1962: 313 (review of some African genera); Schmitz, 1968: 7 (descr., disc., key to Ethiopian fauna); Schuh, 1975: 9, 17 (trichobothria); Carayon, 1977: 21 (key to tribes); Lavabre, 1977a: 57 (desc., review of cocoa pest genera); Carvalho, 1981: 5, 7 (list of spp. for Papua New Guinea, key to gen.); Kerzhner, 1988a: 792 (key to spp. of Far East USSR); Cassis and Gross, 1995: 141 (cat.); Kerzhner and Konstantinov, 1999: 122 (male genitalia).
Odoniellini Carvalho, 1952: 33, 35, 40, 41, 60 (disc., key to tribes, cat.); Carvalho, 1955: 15, 40 (key to tribes, key to gen.); Miller and China, 1957: 430 (key to gen.); Carvalho, 1957: 143 (cat.); Odhiambo, 1962: 271 (generic review, in part); Schuh, 1975: 9, 17 (trichobothria); Carayon, 1977: 21 (key to tribes); Lavabre, 1977a: 48 (descr., key to gen.; review of cocoa pest genera); Carvalho, 1981: 5, 6 (list of spp. for Papua New Guinea, key to gen.); Kerzhner and Konstantinov, 1999: 122 (male genitalia).
Monaloniina Schuh, 1976: 23 (as subtribe, pretarsus, disc.); Schuh, 1995: 508 (cat.); Schuh and Slater, 1995: 176 (disc.)
Odoniellina Schuh, 1976: 32 (as infratribes, pretarsus, disc.); Schuh, 1995 (cat.); Schuh and Slater, 1995: 176 (disc.)
Diagnosis: The Monaloniini differ from other bryocorine suprageneric groups by the unique structure of the thoracic pleura, with the opening of the scent glands obscure, ventrally positioned and without evaporative bodies (fig. 14A in Namyatova et al., in press; see also Cassis, 1986 and Cassis & Schuh, 2012 where this character was initially mentioned), the suture between mesopleuron and metapleuron incomplete (fig. 14A in Namyatova et al., in press), and the metepimeron in many species with a distinct lobe ( Fig. 13A–C View Figure 13 , fig. 14A in Namyatova et al., in press). Other important diagnostic characters for the Monaloniini are: LSI–II more than twice as long as wide (fig. 9A in Namyatova et al., in press); LSIV usually the longest, rarely subequal to LSIII (fig. 8A-E in Namyatova & Cassis, 2013b; fig. 2F in Namyatova & Cassis, 2014; fig. 19A in Namyatova et al., in press), collar weakly separated, usually delimited laterally, hemelytron membrane with single cell, with cell often elongate, surpassing apex of cuneus; basal tooth on claw present (fig. 9A, B in Namyatova & Cassis, 2013b; fig. 21F, I Namyatova et al., in press); parempodia symmetrical (fig. 10 in Namyatova & Cassis, 2013b; fig. 20F in Namyatova et al., in press); unguitractor plate with three contiguous rows of tiles, tiles of middle row straight (fig. 10B in Namyatova & Cassis, 2013b; fig. 20F in Namyatova et al., in press). See also diagnosis of Monaloniini in Namyatova et al. (in press).
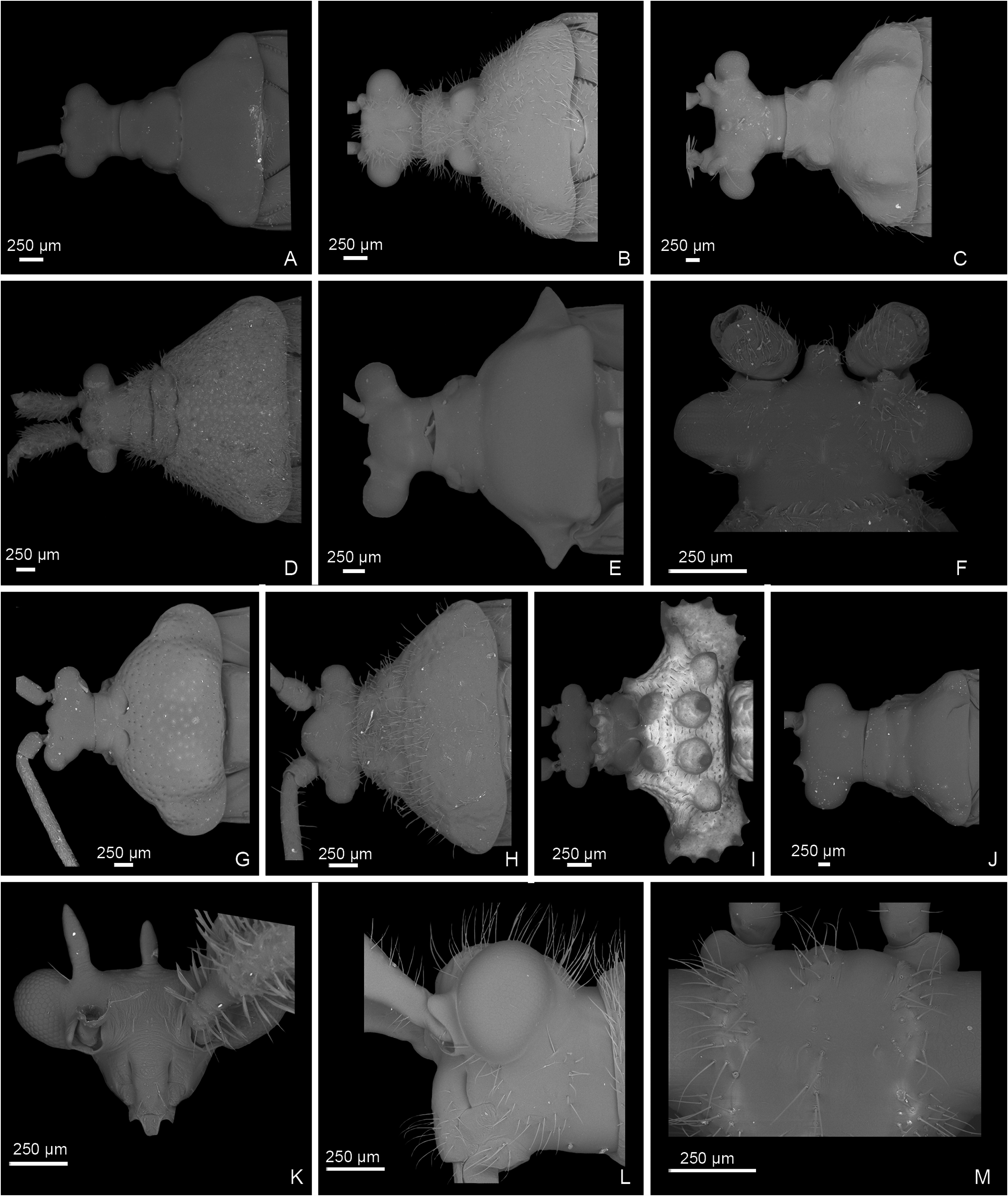
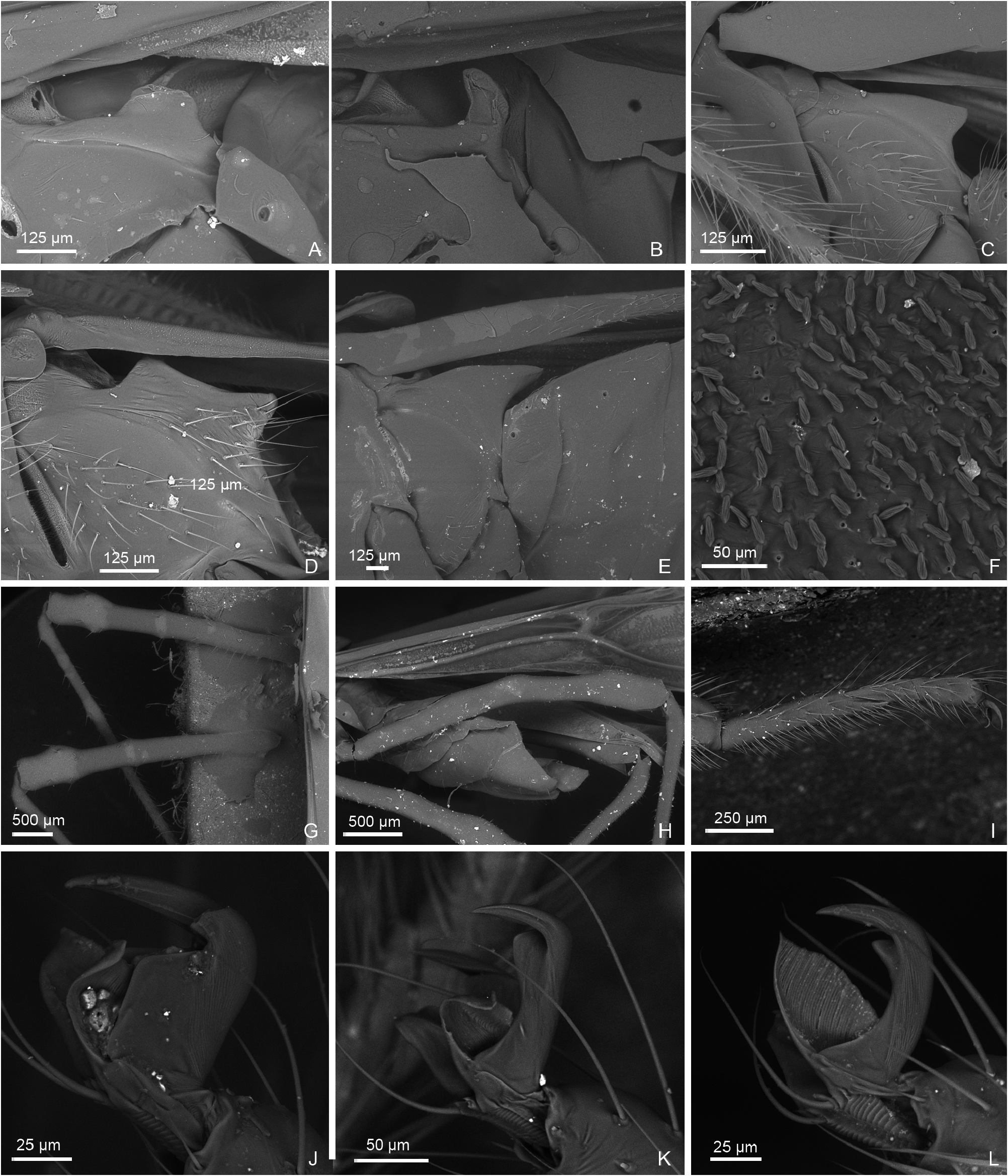
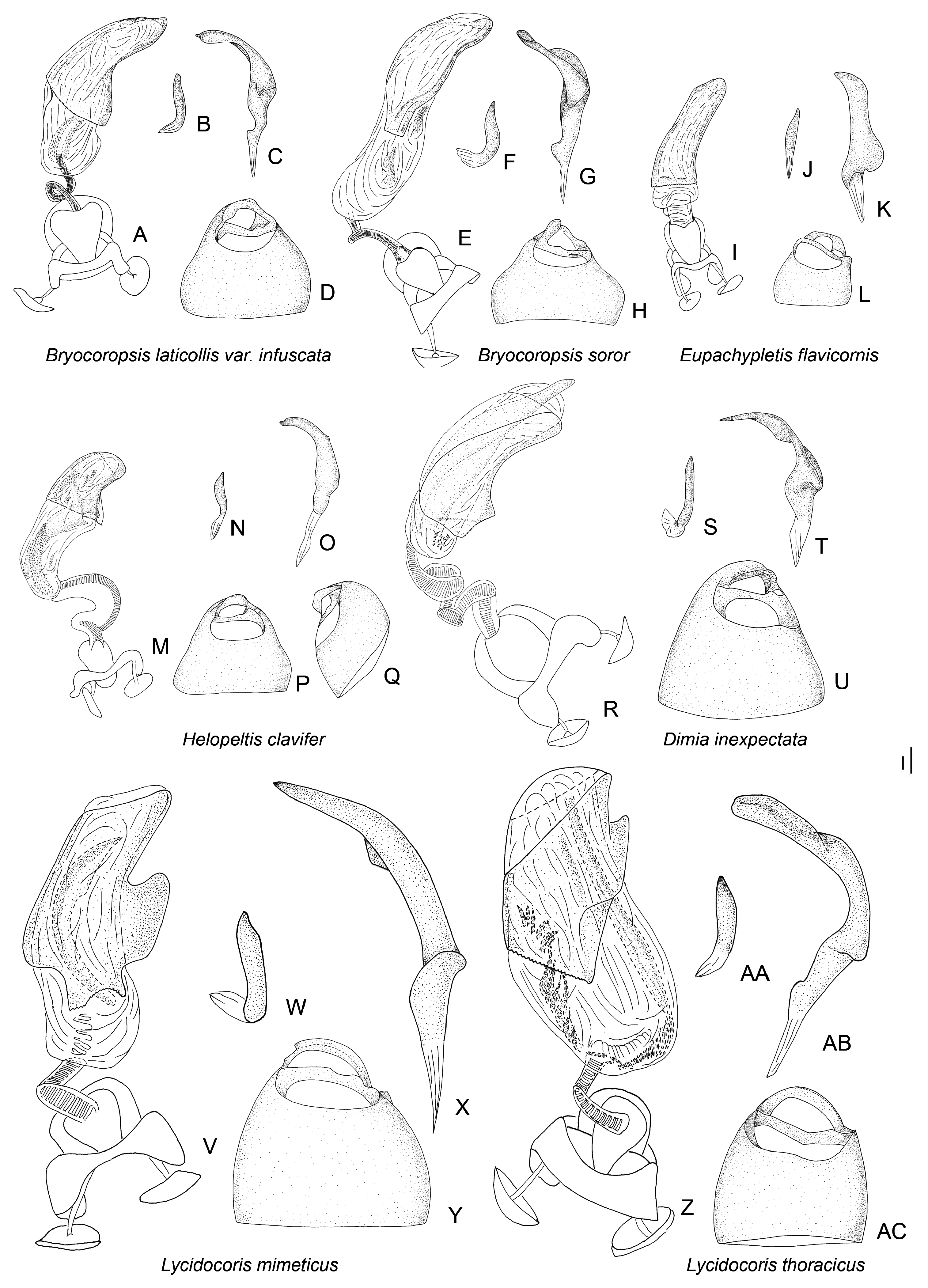
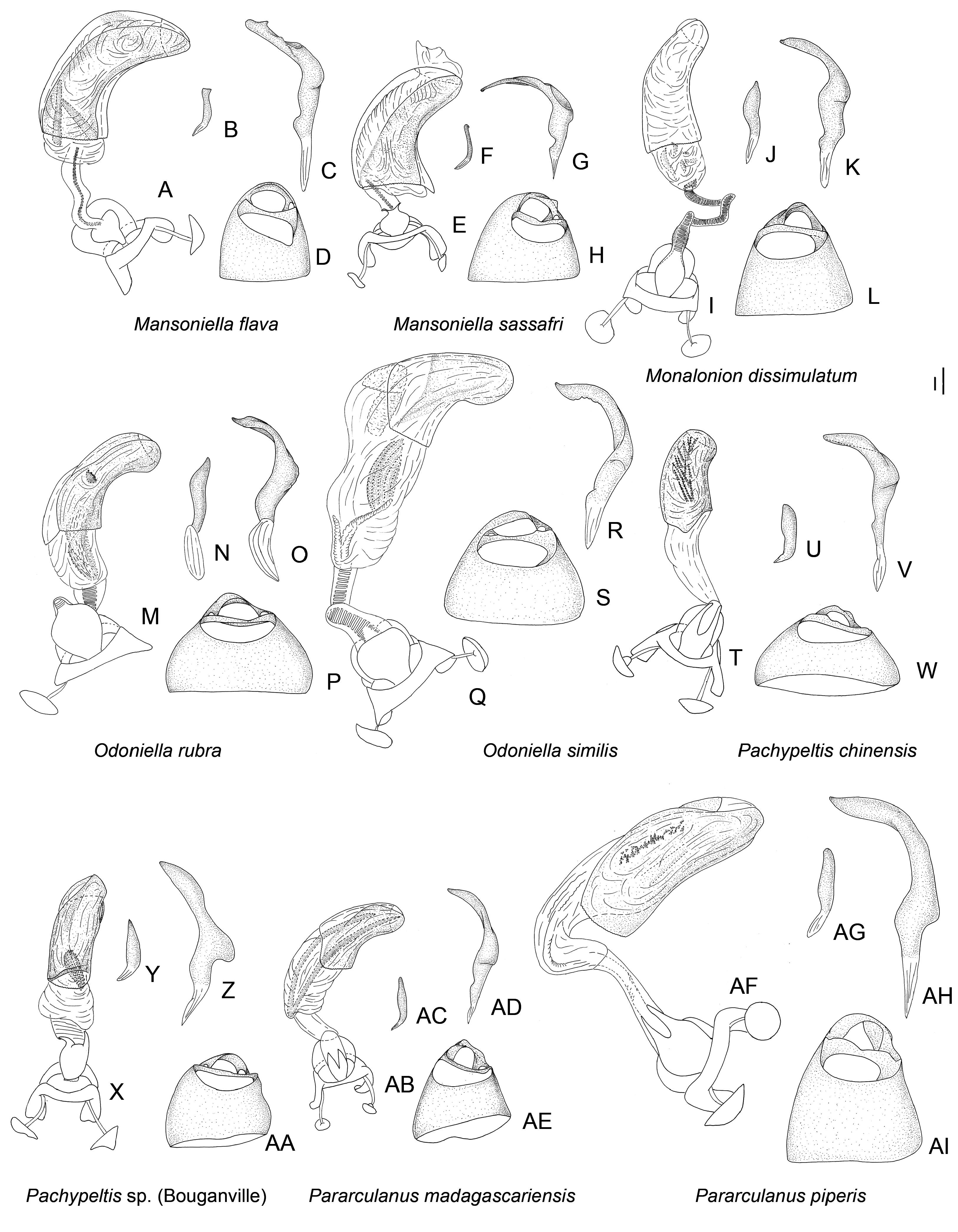
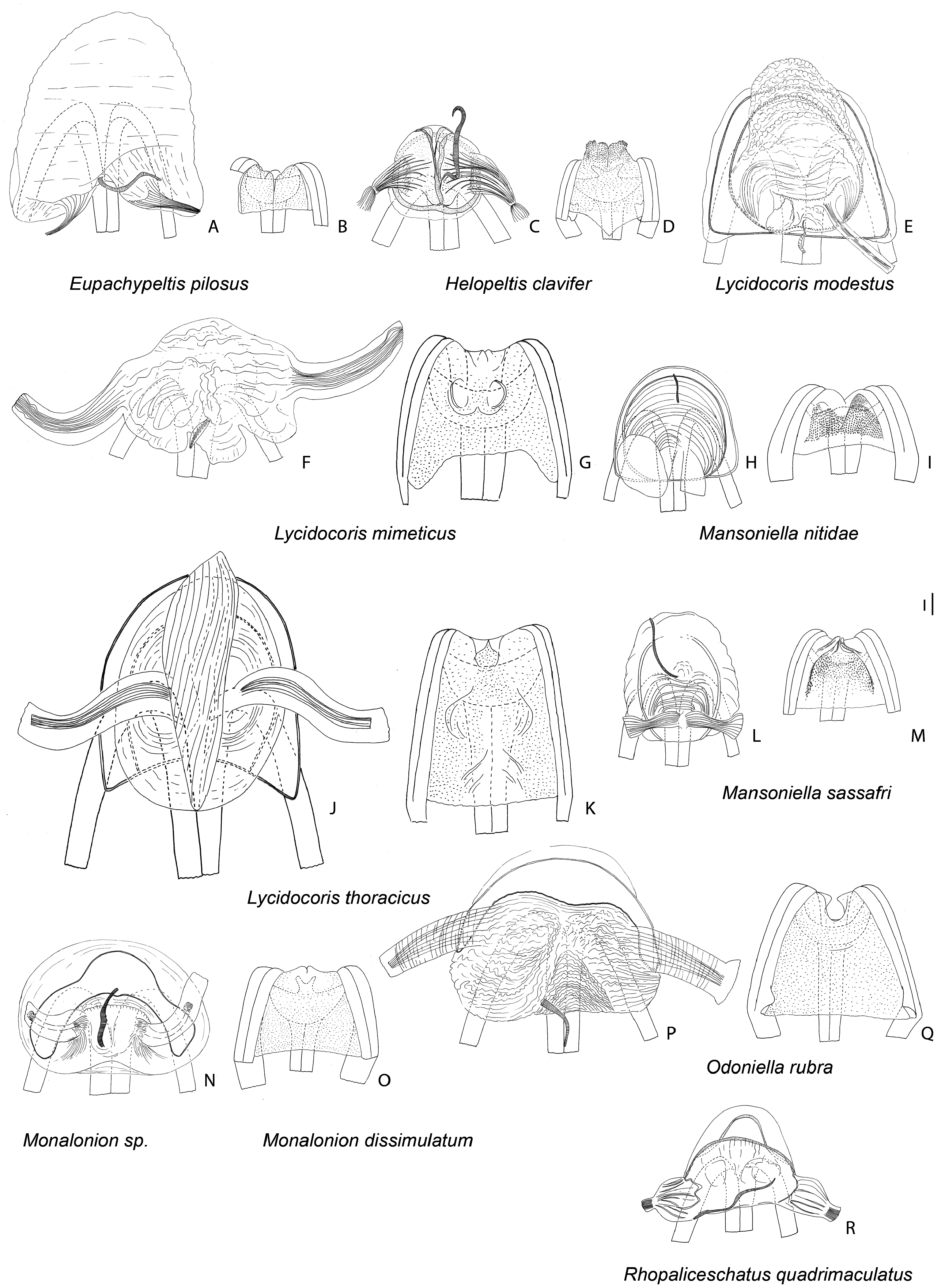
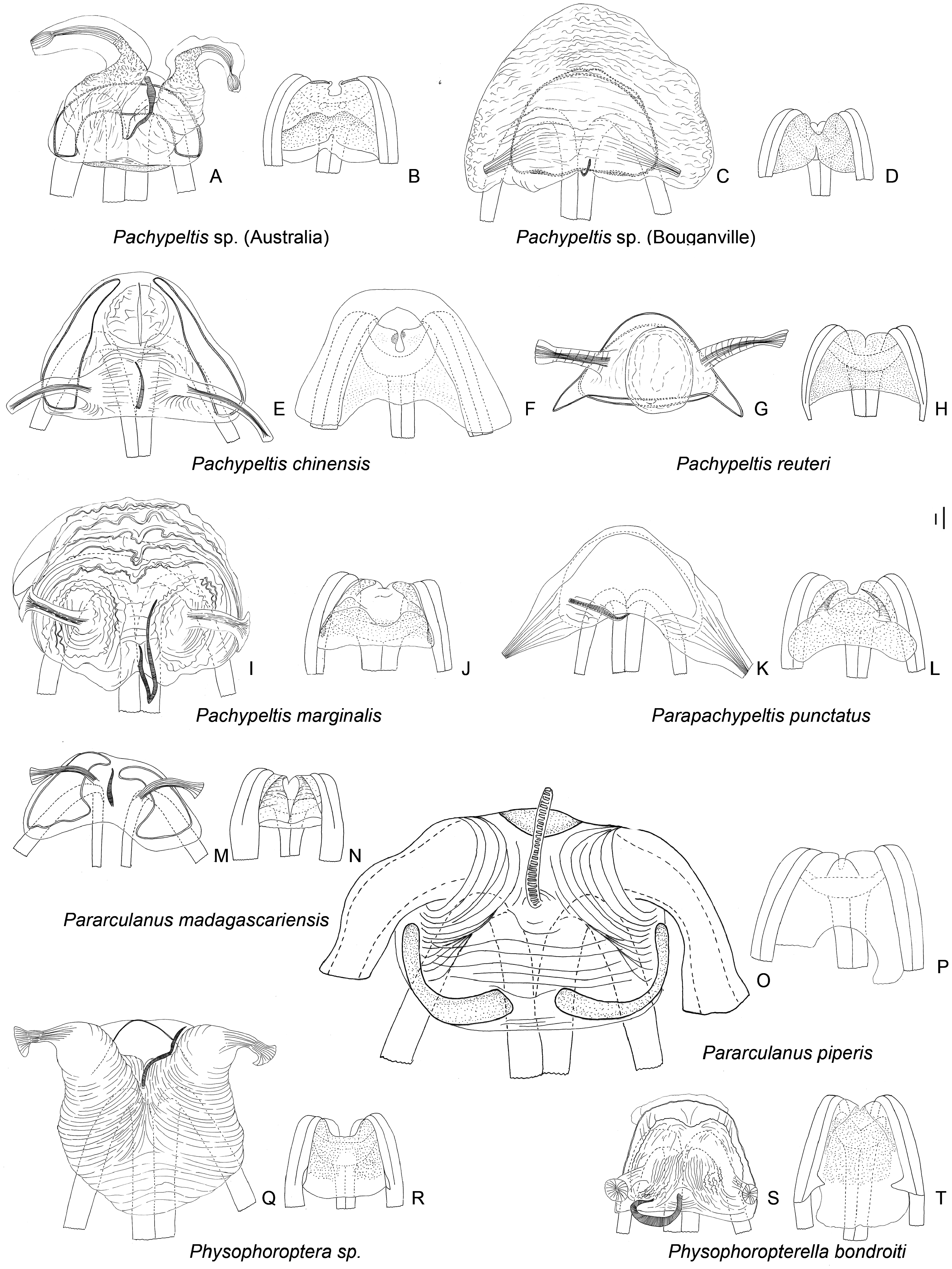
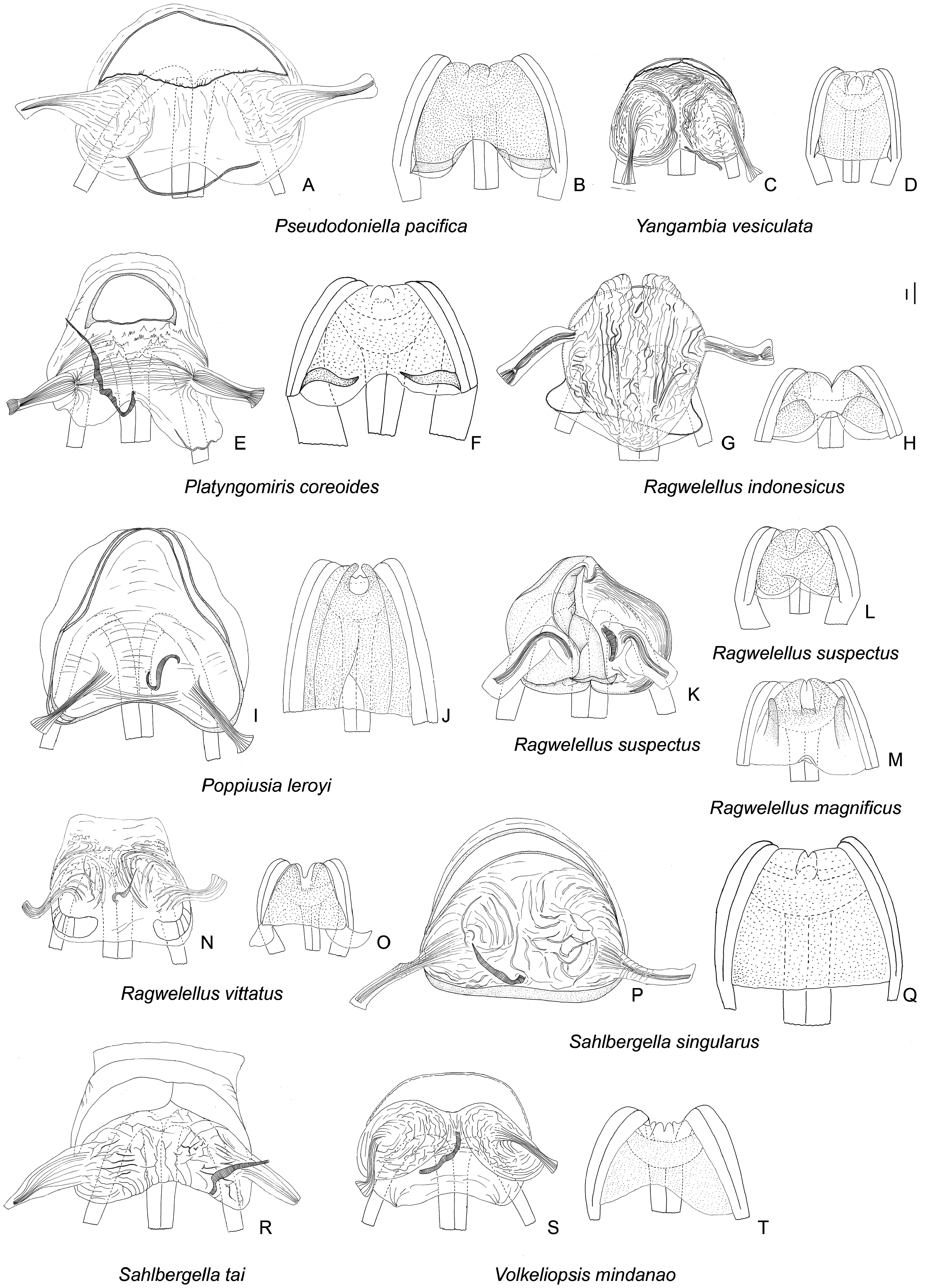
Description: Body size 4.5–14 mm, usually ranging between 8 and 11 mm. COLORATION ( Figs 6–9 View Figure 6 View Figure 7 View Figure 8 View Figure 9 , figs 2– 4 in Namyatova & Cassis, 2013b). Colour usually bright, varying from uniformly pale yellow (e.g. Yangambia vesiculata ) to dark brown (e.g. Sahlbergella singularis ) or bright red (e.g. Physophoroptera mirabilis ). Body usually multicoloured with markings and spots, sometimes with braconid-mimicking (e.g. Rayieria basifer ) or bee-mimicking ( Platyngomiris apiformis ) colour pattern. TEXTURE. Head impunctate, usually with short medial sulcus, which sometimes is very indistinct; some genera with flattened area on vertex near or behind each eye (fig. 4B, C in Namyatova et al., in press); pronotum and scutellum impunctate or punctuate, sometimes punctures mixed with wrinkles or only wrinkles present; pair of punctures on depression delimiting calli and pair of punctures between mesoscutum and scutellum absent or present (fig. 10A in Namyatova et al., in press); small tubercles or tumescences on pronotum and scutellum absent or present ( Figs 10I View Figure 10 , 11D, F–H, J, L View Figure 11 , fig. 4B, C in Namyatova et al., in press); lateral margin of scutellum usually with row of punctures or striations (fig. 11C, D in Namyatova et al., in press), except genera from the Monalonion -complex (see discussion and node 35); sulcus between mesoscutum and scutellum with pair of medial punctures in many genera, except the Monalonion -complex and Odoniella - complex (fig. 11C,D in Namyatova et al., in press); hemelytron generally impunctate, but in many genera bearing rows of punctures on clavus and R + M (fig. 11C, D in Namyatova et al., in press); pleura smooth, impunctate. VESTITURE. Body often clothed with simple setae; in many species of Chamus and some genera of the Odoniella -complex ASI and hemelytra mostly or entirely with flattened setae, sometimes those setae darkened (many genera of the Odoniella -complex); sometimes vestiture scarce, pronotum, scutellum and hemelytron almost without setae; rarely vestiture dense; legs sometimes covered with very long and dense setae ( Sahlbergella theobroma , Platyngomiris apiformis ); small black setae on tibia usually placed in irregular rows (as in fig. 18D in Namyatova et al., in press), sometimes irregularly distributed, present only apically or absent. STRUCTURE. Head. Dorsal view ( Fig. 10 View Figure 10 A-J, fig. 2D in Namyatova & Cassis, 2013a, fig. 5A–F in Namyatova & Cassis, 2013b, fig. 2A–D in Namyatova & Cassis, 2014, fig. 4A–D in Namyatova et al., in press). Eye often removed from pronotum at a distance at least equivalent to half eye diameter or sometimes placed closer to pronotum; occipital region delimited or not delimited by depression; eye stylate or not stylate; distance between antennal fossa varying from as long as to twice as long as antennal fossa diameter; frons often swollen, sometimes straight, sometimes with paired tubercles or more or less bifurcate outgrowth (many representatives of Odoniella -complex), three outgrowths ( Chamus ), or with three shallow ridges ( Eupachypeltis , Dimia , Poppiusia ). Anterior view ( Fig. 10K View Figure 10 , fig. 2B in Namyatova & Cassis, 2013a, fig. 6 in Namyatova & Cassis, 2013b, fig. 2B, E in Namyatova & Cassis, 2014, fig. 3A, B in Namyatova et al., in press). Varying from as wide as long to almost twice as wide as long; from anterior view eye oval, higher than wide or roundish; eye height varying from distinctly longer to slightly shorter than distance between eye and apex of clypeus; antennal fossa round distinctly shorter than eye height or oval, varying from subequal to half of eye height to almost subequal to eye height; inferior margin of antennal fossa slightly above inferior margin of eye, but sometimes at the same level or below inferior margin of eye; base of clypeus placed below, slightly above or at half eye height, often distinctly delimited basally, sometimes not delimited. Lateral view ( Fig. 10L View Figure 10 , fig. 2A in Namyatova & Cassis, 2013a, fig. 7 in Namyatova & Cassis, 2013b, fig. 2C in Namyatova & Cassis, 2014, fig. 6A-C in Namyatova et al., in press). Head swollen of almost flat; in lateral view eye oval, margin of eye surpassing clypeus, but not reaching maxillary plate; maxillary and mandibular plates subrectangular; buccula shortened, almost as long as wide or elongate; gula usually longer than buccula or shortened, straight or convex. Labium (fig. 8A, E in Namyatova & Cassis, 2013b, fig. 9A in Namyatova et al., in press). Length varying from very short, slightly surpassing anterior margin of prosternum to reaching abdominal segments III–IV; LSI–III usually longer than wide, more or less subequal in length, sometimes LSIII shortened, LSIV usually elongate, longer than each of previous segments; sometimes LSIV as long as LSIII or all segments shortened, almost as long as wide. Antenna. Shape and length varying from short and wide, distinctly shorter than body, to long and filiform, distinctly longer than body. Thorax. Pronotum ( Fig. 10 View Figure 10 A- E, G-J, fig. 2D in Namyatova & Cassis, 2013a, fig. 5A– E in Namyatova & Cassis, 2013b, fig. 4A–D in Namyatova et al., in press). Collar often delimited only laterally, fused with callosite area medially, sometimes delimited posteriorly with shallow depression, flat or swollen; calli varying from flat to distinctly raised, sometimes fused with each other, delimited or not delimited by sulcus posteriorly; humeral angles of pronotum dilated or flat, sometimes acute; posterior margin of pronotum varying from almost straight to distinctly concave. Scutellum ( Figs 11 View Figure 11 A-K, 12, fig. 9H in Namyatova & Cassis, 2013b, figs 11C, D, G, 12E, F in Namyatova et al., in press). Varying from flat to distinctly swollen of different shape, sometimes with longitudinal depression, in some with elongate vertical process ( Afropeltis , Helopeltis , Physophoropterella ). Pleura ( Fig. 13A–E View Figure 13 , fig. 9C in Namyatova & Cassis, 2013b, fig. 3d in Namyatova & Cassis, 2014, fig. 14A in Namyatova et al., in press). Mesothoracic apodeme round, open. Metathoracic spiracle oval, open, without evaporative bodies bounding it. Metathoracic gland ostiole placed ventrally, indistinct, evaporative area absent; suture between meso- and metapleuron incomplete; metepimeron often enlarged or narrow with lobe or elongate projection, sometimes angulate, rarely rounded. Posterior margin of metasternum rounded (fig. 17B in Namyatova et al., in press) or with medial projection on to abdomen (fig. 17A in Namyatova et al., in press). Hemelytron ( Figs 6–9 View Figure 6 View Figure 7 View Figure 8 View Figure 9 , figs 2–4, fig. 9A, B, D–G in Namyatova & Cassis, 2013b, figs 12C– F, 13E, F in Namyatova et al., in press). Costal margin straight or concave; hemelytron sometimes tapering towards apex; claval commissure of different length, its margins straight or curved; R + M distinct, reaching or not reaching posterior margin of corium; medial fracture distinctly inclined towards midline or sub- parallel to costal margin of hemelytron; corium rarely with swelling posteriorly ( Physophoroptera and Physophoropterella ) ( Fig. 13F View Figure 13 ); length of cuneus varying from 2 to 6 times as long as its base, medial margin of cuneus straight or concave; membrane with single cell, cell often surpassing apex of scutellum, rarely only reaching or almost reaching apex of cuneus, of different length, its apex acute or rounded. Legs. Length varying from very short to elongate. Coxae short, length varying from almost as long as wide to twice as long as wide; forecoxae contiguous (fig. 17A in Namyatova et al., in press) or separated (fig. 17B in Namyatova et al., in press); hind and middle coxae separated (fig. 17A, B in Namyatova & Cassis, in press). Femora (13G, H, fig. 18A, C in Namyatova et al., in press). Straight or curved, sometimes only hind femur slightly curved, sometimes with swellings apically and medially. Tibiae straight or slightly curved, as long as or longer than femora, sometimes with swellings. Hind tarsus ( Fig. 13I View Figure 13 , fig. 2C in Namyatova & Cassis, 2013a, fig. 8F in Namyatova & Cassis, 2013b, fig. 3H in Namyatova & Cassis, 2014, fig. 19A, B in Namyatova et al., in press) with segments subequal in length, or segment I longer than others; tarsal segment III incrassate; guard setae long. Pretarsus ( Fig. 13J–L View Figure 13 , fig. 20F in Namyatova et al., in press, fig. 2F in Namyatova & Cassis, 2013a, fig. 10 in Namyatova & Cassis, 2013b, fig. 3f, i in Namyatova & Cassis, 2014); unguitractor plate with three contiguous rows of tiles, with lateral rows wider than middle row; claw with basal tooth short triangular or subrectangular, sometimes concave or subdivided into basal and apical parts; parempodia present, symmetrical; pseudopulvilli present, as long as or shorter than claw. Male genitalia ( Figs 14– 17 View Figure 14 View Figure 15 View Figure 16 View Figure 17 , fig. 3A–D in Namyatova & Cassis, 2013a, figs 11– 13 in Namyatova & Cassis, 2013b, fig. 4 in Namyatova & Cassis, 2014, fig. 22A–H in Namyatova et al., in press). Genital capsule variable in shape, sometimes ventral wall shortened anteriorly; supragenital bridge present; left paramere 2–4× as long as right paramere, r-shaped or almost straight; right paramere reduced; phallobase sclerite of primary gonopore of different shape, sometimes with outgrowth in front of ductus seminis attachment place ( Helopeltis ) or with pair of outgrowths supporting ductus seminis; length of ductus seminis variable, with or without coils, attached medially or on left-hand side; ductus seminis sometimes with sclerite around secondary gonopore, rarely base of secondary gonopore also sclerotized ( Monalonion ); phallotheca of aedeagus distinctly sclerotized dorsally and membranous laterally and ventrally, sometimes only very narrow area of phallotheca sclerotized; endosoma membranous, not subdivided, often with elongate spicules or fields of small spicules. Female genitalia ( Figs 18–21 View Figure 18 View Figure 19 View Figure 20 View Figure 21 , fig. 3E–F in Namyatova & Cassis, 2013a, fig. 14 in Namyatova & Cassis, 2013b, fig. 5 in Namyatova & Cassis, 2014, fig. 23H in Namyatova et al., in press). DLP with one or two sclerotized bands, sometimes with medial sclerotized circle or with paired sclerotized rings, sometimes also with sclerotized ridge medially or small additional sclerites medially, sometimes entirely membranous; DLP sometimes with dense striations, especially around places of attachment of lateral oviducts; attachment of lateral oviducts varying; spermathecal gland usually attached medially, in anterior or posterior part of DLP, rarely at midpoint, sometimes shifted to right- or left-hand side; posterior wall of bursa copulatrix often with small tubercles, sometimes with outgrowths or sclerotization, rarely with posterior wall entirely membranous; base of second valvula concave, straight or convex, sometimes with bifurcated outgrowth; ventral wall of bursa copulatrix with or without sclerotization bounding vulva.
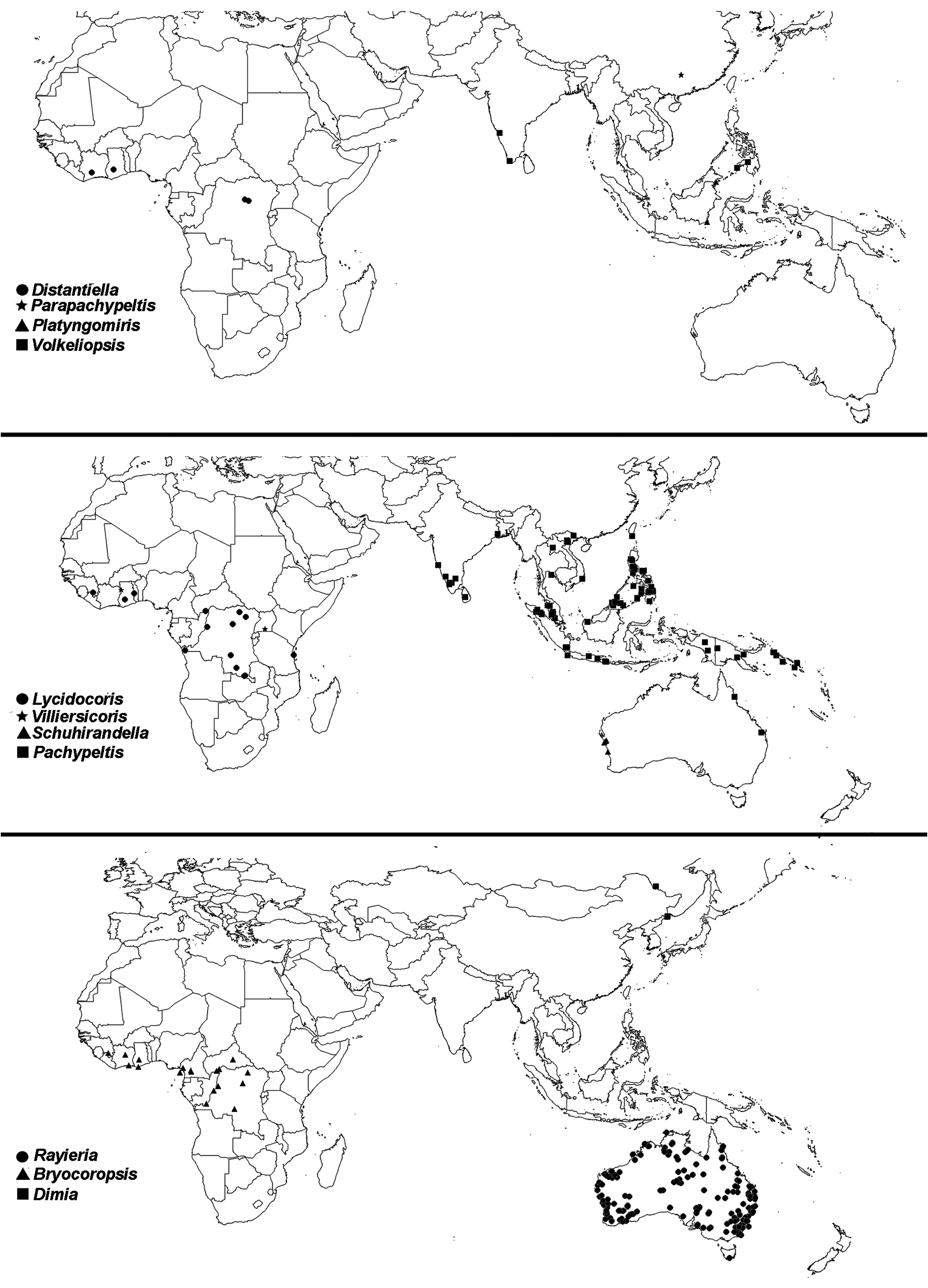
Distribution: Circumtropical. Most diverse in Africa and Indo-Pacific, with some genera known from Australia, Eastern Palaearctic and South America ( Figs 22–24 View Figure 22 View Figure 23 View Figure 24 ).
Host plants: Monaloniines are known to feed on a wide range of plants, with some of them being pests of cocoa, tea, cashew and other cultivated plants (e.g. Schmitz, 1968; de Abreu, 1977; Lavabre, 1977a,b; Piart, 1977; Hill, 1983; Stonedahl, 1991).
Discussion: The position of the Monaloniini within the Bryocorinae was discussed by Namyatova et al. (in press). Previous to this study, the Monaloniini includ- ed 21 genera, all of which were listed in Schuh (1995: 1995–2013), aside from our recent description of Schuhirandella ( Namyatova & Cassis, 2013a) . Felisacus was transferred to the tribe Felisaciini by us (Namyatova et al., in press).
We have removed Onconotellus Knight, 1935 and Pachypeltopsis Poppius, 1912 from the Monaloniini . Knight (1935) initially placed Onconotellus within the subfamily Dicyphini . Cassis (1986) tentatively transferred Onconotellus to the tribe Monaloniini based on the original description. Knight’s illustrations showed two membrane cells, and convergent flattened parempodia without pseudopulvilli. These characters are common for the subfamily Orthotylinae , and we transfer Onconotellus to it (see Schuh, 1976 for a discussion of the pretarsus). Poppius (1912) described the monotypic bryocorine genus Pachypeltopsis , and Carvalho (1952) placed it in the tribe Monaloniini . We examined the holotype of Pachypeltopsis australicus , and observed a distinct collar, delimited by a deep sulcus, two membrane cells, and setiform parempodia and no pseudopulvilli. On the basis of these characters we transfer this genus to the tribe Saturniomirini . One of us (A.N.N.) also examined the monotypic genus Felisacoris Carvalho, 1956 , and found that it is very similar to Felisacus in external view and genitalia and most probably it is nested within this genus. Based on this observation, we transfer Felisacoris to the tribe Felisacini , and the full revision of the groups will be provided in a subsequent paper.
Our phylogenetic analysis resulted in the recognition of two major clades within a redefined tribe, namely the Monalonion -complex and Odoniella -complex (see Namyatova & Cassis, 2013a,b, Namyatova et al., in press). These complexes comprise more than half of the monaloniine genera, and correspond in part to Carvalho’s (1952, 1955) notions of Monaloniini and Odoniellini (see also nodes 12–14 in Namyatova et al., in press). We have refrained from redefining these subtribes, as it would require us to erect new and less supported subtribes for the remaining monaloniine genera, and we prefer the use of informal groups pending further phylogenetic analysis. These two complexes comprise the following genera:
1) The Monalonion -complex includes Afropeltis , Helopeltis , Monalonion , Physophoroptera , Physophoropterella , Rayieria , Ragwelellus , Physophoroptera , Physophoropterella , and Schuhirandella (node 35). The group of species is characterized by the following set of characters: pronotum and scutellum without punctures or wrinkles, punctures on R + M and clavus absent, forecoxae separated from each other, outgrowth on metepisternum absent. The representatives of this group are usually elongate bugs with long antennae and legs, although some genera are more oval with relatively short appendages ( Schuhirandella , Physophoroptera ).
2) The Odoniella -complex includes Boxia , Bryocoropsis , Distantiella , Odoniella , Platyngomiris , Pseudodoniella , Rhopaliceschatus , Sahlbergella , Volkeliopsis Poppius, 1915 , Volkelius and Yangambia (node 19). Representatives of this group and oval bugs, with more or less swollen scutellum, pronotum and scutellum with distinct punctures or wrinkles, R + M and clavus without punctures, antennal segments III and IV clavate or incrassate apically.










No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
MONALONIINI
| Namyatova, Anna A. & Cassis, Gerasimos 2016 |
Monaloniina Schuh, 1976: 23
| Schuh RT & Slater JA 1995: 176 |
| Schuh RT 1976: 23 |
Odoniellina Schuh, 1976: 32
| Schuh RT & Slater JA 1995: 176 |
| Schuh RT 1976: 32 |
Monaloniini
| Kerzhner IM & Konstantinov FV 1999: 122 |
| Cassis G & Gross GF 1995: 141 |
| Kerzhner IM 1988: 792 |
| Carvalho JCM 1981: 5 |
| Carayon J 1977: 21 |
| Lavabre EM 1977: 57 |
| Schuh RT 1975: 9 |
| Schmitz G 1968: 7 |
| Odhiambo TR 1962: 313 |
| Carvalho JCM 1957: 131 |
| Carvalho JCM 1955: 16 |
| Carvalho JCM 1952: 33 |