
Lysmata malagasy, Ashrafi & Baeza & Ďuriš, 2021
|
publication ID |
https://doi.org/ 10.5852/ejt.2021.774.1535 |
|
publication LSID |
lsid:zoobank.org:pub:E5039C89-D341-4F71-9738-F4878F9E5E50 |
|
DOI |
https://doi.org/10.5281/zenodo.5572657 |
|
persistent identifier |
https://treatment.plazi.org/id/62A66440-6F8E-4BD3-94E3-5CAD9D2D336E |
|
taxon LSID |
lsid:zoobank.org:act:62A66440-6F8E-4BD3-94E3-5CAD9D2D336E |
|
treatment provided by |
Felipe |
|
scientific name |
Lysmata malagasy |
| status |
sp. nov. |
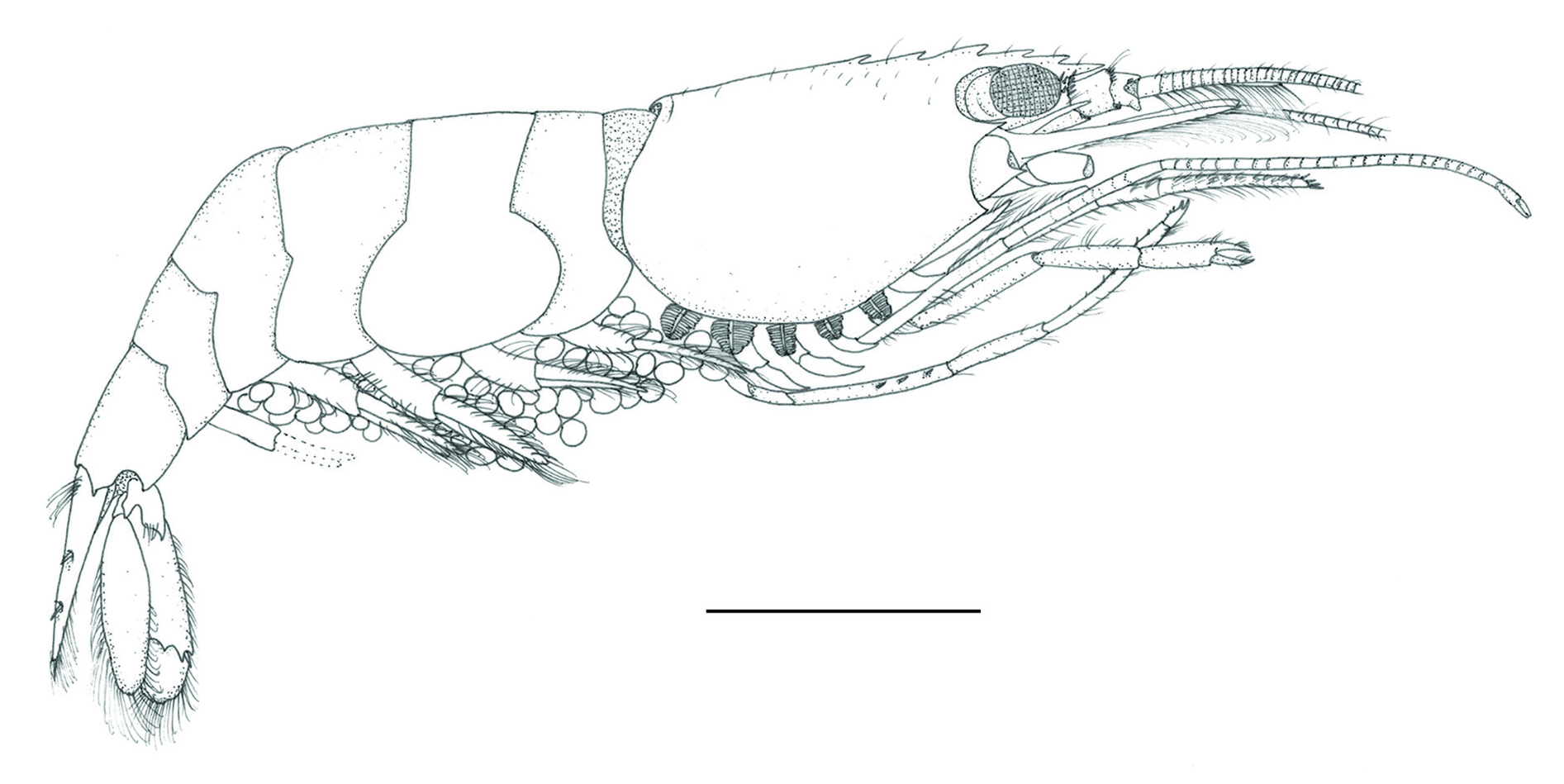
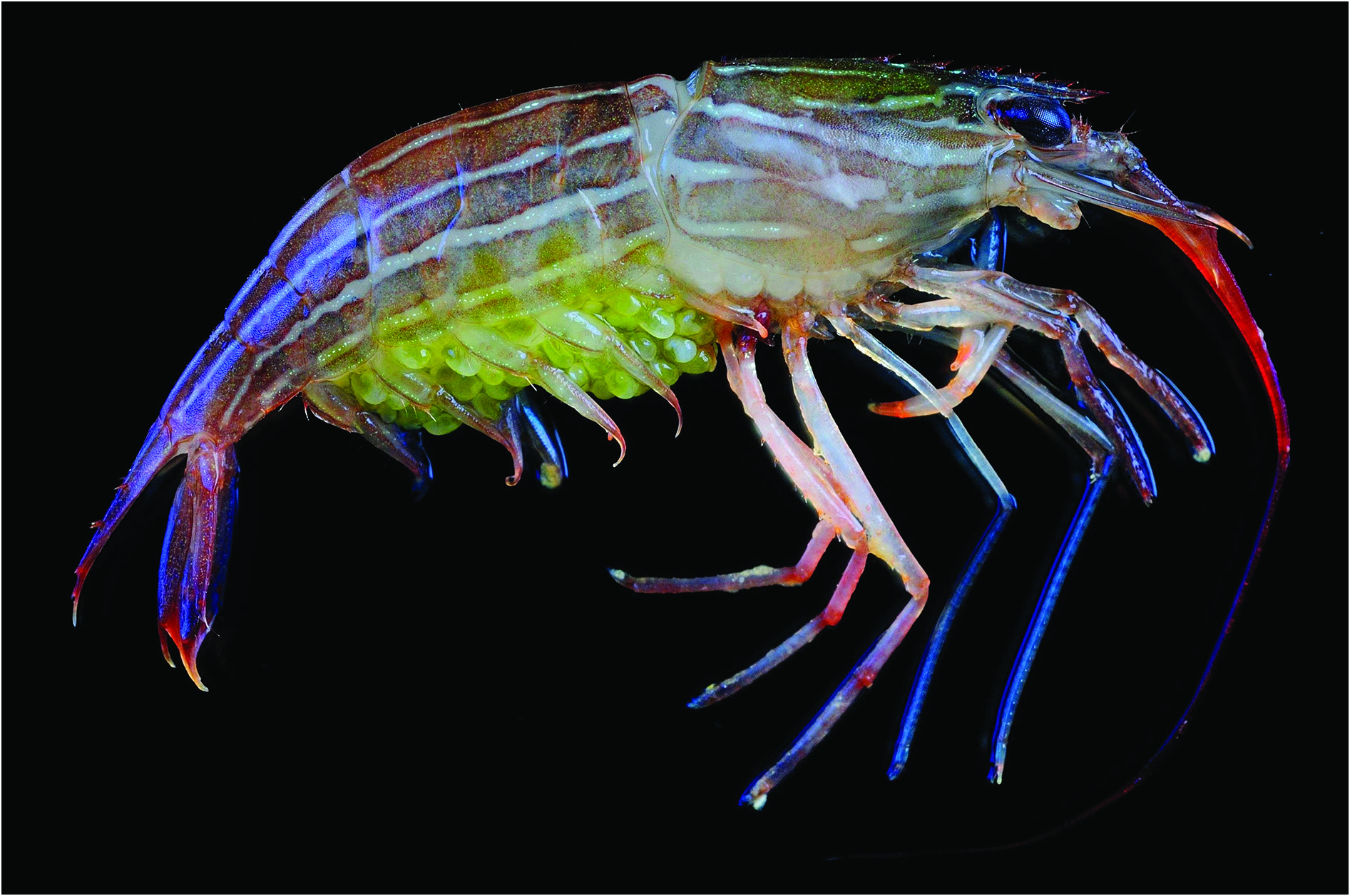
Lysmata malagasy sp. nov.
urn:lsid:zoobank.org:act:62A66440-6F8E-4BD3-94E3-5
Figs 3 7 View Fig View Fig View Fig View Fig View Fig
Etymology
The new species is named after ‘Malagasy’, the native inhabitants of Madagascar where the specimen was collected; the term ‘Malagasy’ is English version of the word ‘Malgache’ in French used for both people and their language in Madagascar; used as a noun in apposition.
Type material
Holotype MADAGASCAR • 1 ovigerous hermaphrodite (CL 8.8 mm); southern Madagascar ; 24°58′4.8″ S, 47°06′8.4″ E; depth 3–8 m; 6 May 2010; MNHN exped.; Atimo-Vatae; Stn TR05 1; GenBank MW798244 View Materials ; MNHN-IU-2010-4890 . GoogleMaps
Description
Medium-sized lysmatid shrimp
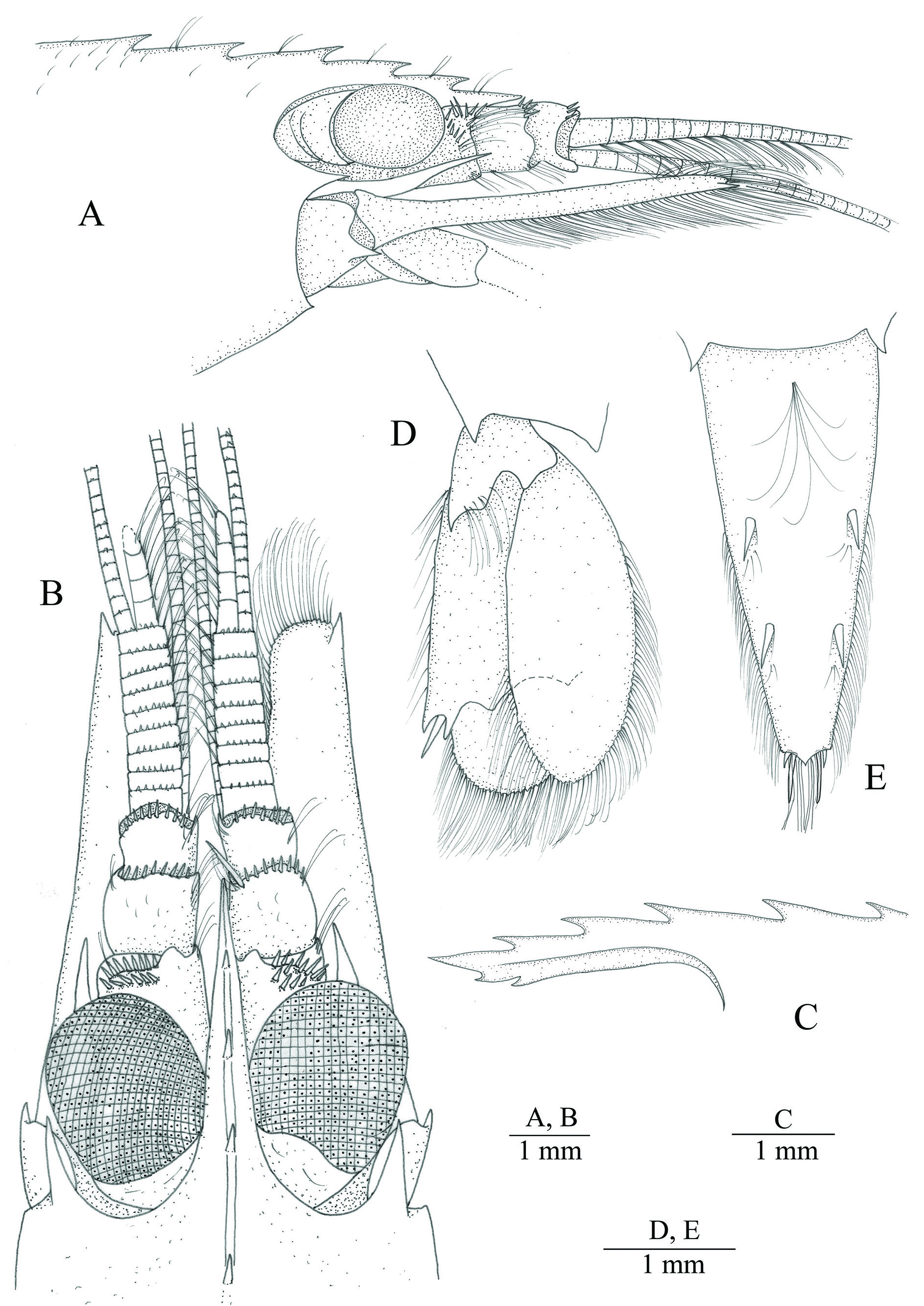
CARAPACE ( Figs 3 View Fig , 4A–C View Fig ). Smooth with scattered setae on dorsal margin, antennal tooth strong, pterygostomial margin slightly produced, pointed; with two postrostral dorsal teeth located anteriorly at about 0.2 and 0.4 of carapace length. Rostrum elongated, approximately half of CL, reaching to about
end of second antennular article, straight in lateral view, bearing evenly distributed three teeth dorsally; ventral margin with two teeth at distal-third.
PLEON ( Fig. 3 View Fig ). Smooth; first pleuron obtusely angulate anteriorly, rounded postero-ventrally; second and third pleura rounded antero- and postero-ventrally; fourth pleuron angulated postero-ventrally, not pointed; fifth pleuron produced postero-ventrally, subacute; sixth pleuron with triangular projections postero-laterally.
TELSON ( Fig. 4E View Fig ). Elongated, slightly more than twice as long as broadest part, tapering posteriorly; dorsal surface with antero-median tuft of long setae and two pairs of spines about 0.1 of telson length and situated at 0.5 and 0.75 of telson length; distal half of lateral margins furnished with setae; posterior margin broadly triangular, projecting medially into acute apex, with two pairs of slender submedian setae and two pairs of laterally placed spines, mesial spines long, about 1.2 times as long as dorsal spines and more than three times as long as lateral ones, latter small, about 2.5 times as short as dorsal spines.
ANTENNULE ( Fig. 4A–B View Fig ). With peduncle stout; basal peduncular segment longer and stouter than others, reaching about to base of most anterior dorsal rostral tooth, with group of spinules antero-dorsally; stylocerite slender, acute, falling short of reaching to middle of second antennular segment; second peduncular segment stouter and longer than third one, with few spinules along antero-dorsal margin; third segment about 0.7 as long as second one, with row of spinules furnishing distal margin. Lateral (= upper) flagellum biramous, fused part consisting of eight or nine articles (right and left antennule in holotype, respectively), each article of fused part with row of small spinules situated on anterior margin, short free ramus (= accessory ramus) comprising of four elongated articles; aesthetascs present from distal end of first article of fused part to end of short free ramus. Longer free ramus of upper flagellum filiform, each article with one or two small spinules on anterior margin. Lower antennular flagellum filiform, long.
ANTENNA ( Fig. 4A–B View Fig ). With scaphocerite about 0.8 as long as CL, elongated, five times as long as broad, lateral margin straight, distolateral tooth acute, slightly overreaching blade, slightly overreaching fused part of lateral antennular flagellum, basicerite with acute inferior tooth, falling short of middle of cornea, superior margin rounded; carpocerite small, reaching to about end of first antennular segment.
EYES. With cornea about 0.2 of CL, stalk about a third of cornea.
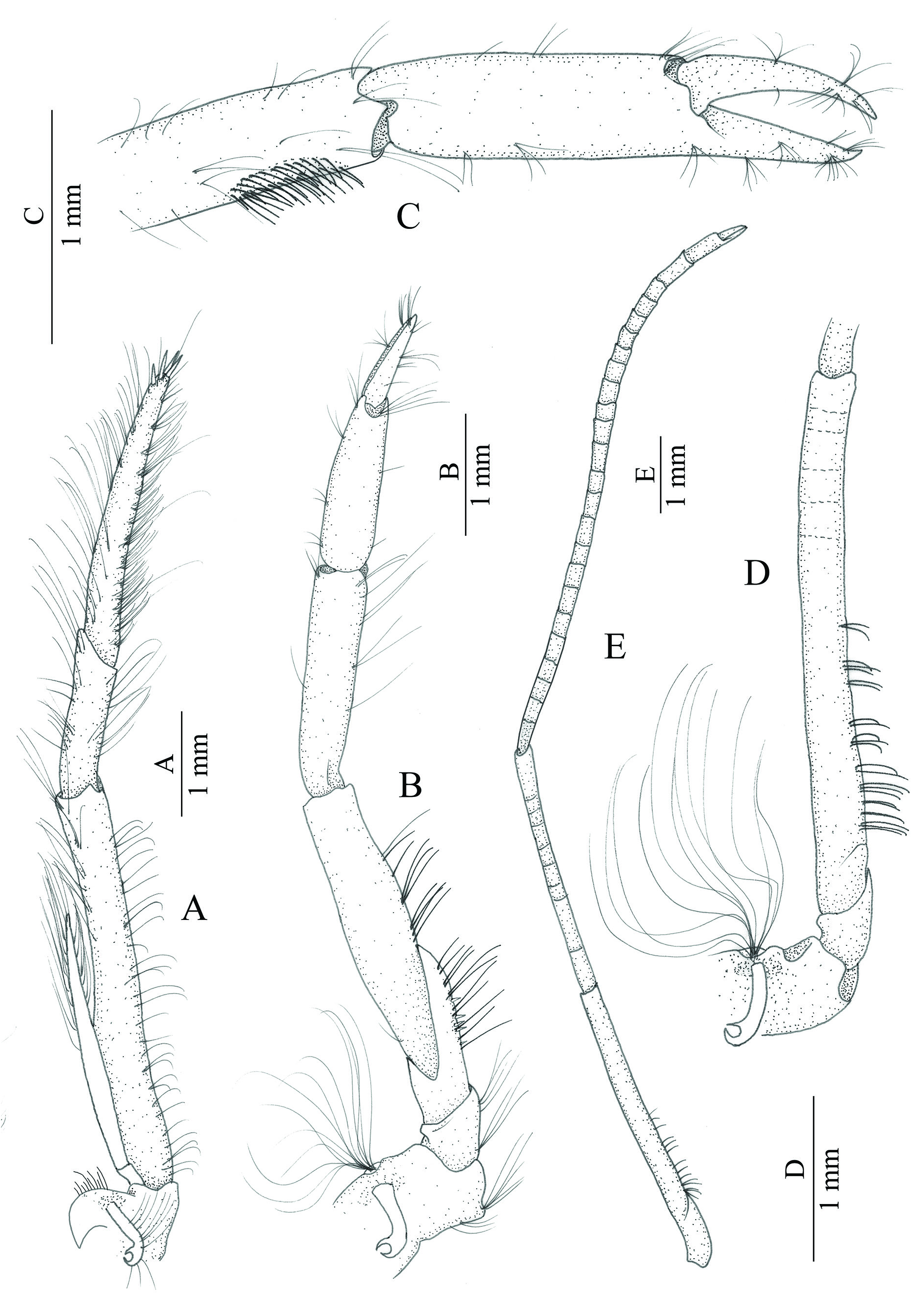
MOUTH PARTS. Not dissected except for third maxilliped. Second maxilliped with podobranch. Third maxilliped ( Fig. 5A View Fig ) pediform, bearing broadly hooked lateral plate, group of short distolateral setae, strap-like epipod, and arthrobranch. Endopodite stout, with numerous slender serrulate setae dorsally as well as ventrally; antepenultimate segment long, slender, more than nine times as long as deep; penultimate segment about 0.4 as long as antepenultimate one, four times as long as deep; ultimate segment long, slender, slightly tapering distally, approximately 0.75 as long as antepenultimate segment, bearing three terminal spinules and single similar subterminal one. Exopodite feebly developed, reaching to distal two thirds of antepenultimate segment, distal half with numerous slender marginal setae.
FIRST PEREIOPOD ( Fig. 5B–C View Fig ). As in other lysmatid shrimps, similar, equal, smaller than other pereiopods, slightly overreaching scaphocerite when extended; coxa with strap-like epipod and tuft of numerous long setobranchs; basis small, with no special structures; ischium about three times as long as basis, with several small spinules and several thick setae on ventral margin; merus nearly twice as long as ischium, about 5.5 times as long as broadest part in midlength, with few thick setae on mid-ventral margin; carpus about 0.8 times as long as merus, more than three times as long as broad, furnishing with setal brush disto-ventro-mesially; chela with palm elongated, more than 1.5 times as long as fingers, lacking thick setae; fingers simple, without gap when closed.
SECOND PEREIOPODS ( Fig. 5D–E View Fig ). Subequal, similar, about 2.5 times as long as CL, more slender than other pereiopods, coxa with strap-like epipod and tuft of numerous setobranchs; basis short; ischium long, about 7 times as long as basis, with relatively thick setae on ventral margin at proximal-half, distal half with several weak subdivisions; merus long, about 0.8 as long as ischium, with eight articles; carpus long, more than twice as long as merus, subdivided into 23 articles; chela small, with palm about 1.5 times as long as fingers, fingers simple.
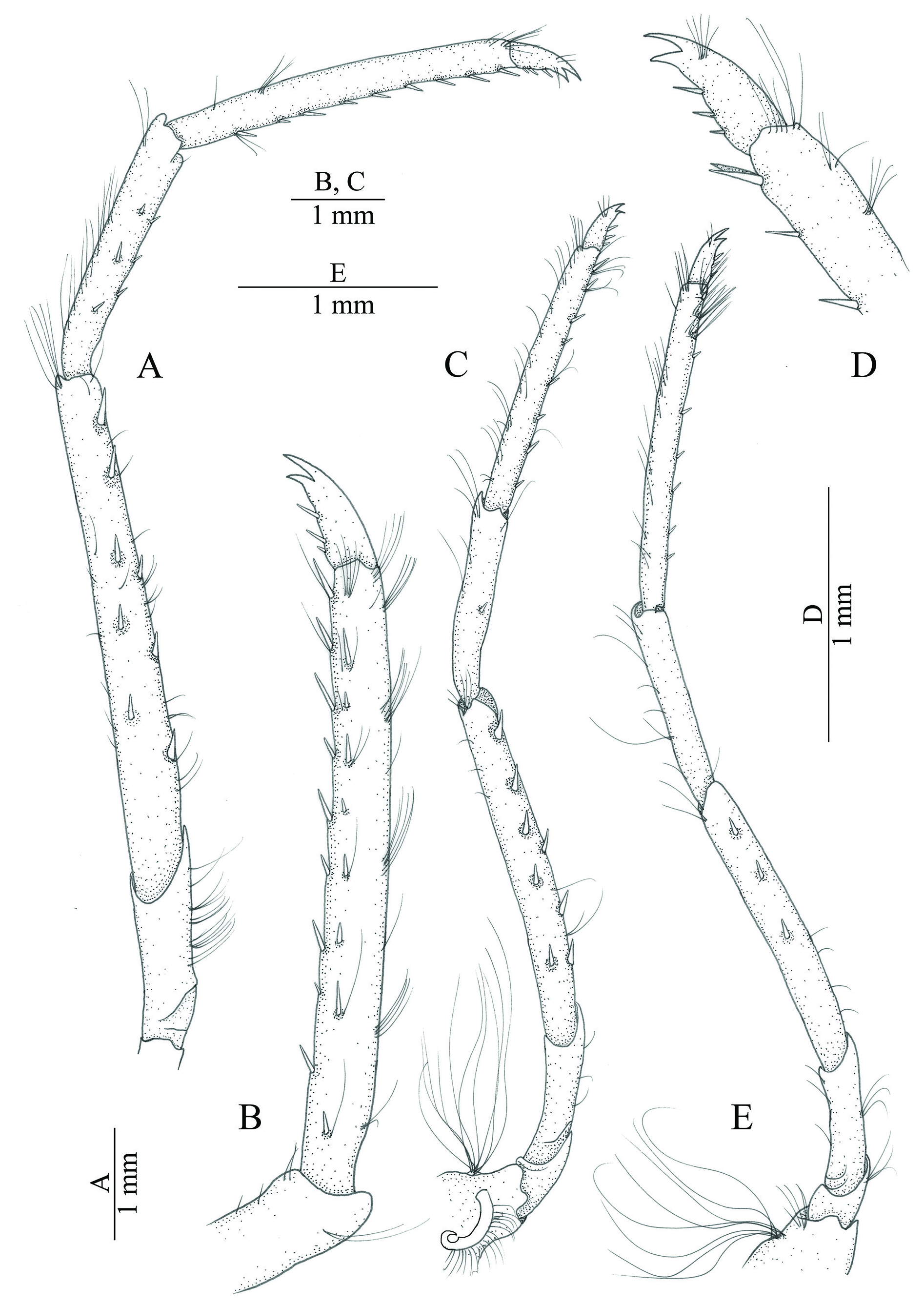
THIRD PEREIOPOD ( Fig. 6A–B View Fig ). Longer than other walking pereiopods; coxa with strap-like epipod and tuft of numerous setobranchs; merus more than three times as long as ischium, with two rows of spines on latero-ventral margin, one row with five and other row with three spines; carpus about 0.5 times as long as merus, with two rows of spines, each row with three spines; propodus about 1.3 times as long as carpus, bearing two rows of spines on ventral margin and one pair distally, each row with eight spines; dactylus biunguiculate, superior unguis slightly longer than inferior, inferior unguis slightly broader at base than superior, flexor margin with three accessory spines.
FOURTH PEREIOPOD ( Fig. 6C–D View Fig ). Generally similar to third one, carpus with only one spines on ventral margin, propodus with single row of seven spines ventrally and one pair distally.
FIFTH PEREIOPOD ( Fig. 6E View Fig ). Generally similar to fourth pereiopod; with coxa bearing setobranchs, without strap-like epipod; merus with only one row of three spines implemented ventrally; carpus unarmed, propodus with two small transverse row of cleaning setae distally on ventro-mesial margin.
UROPOD ( Fig. 4D View Fig ). With dorso-distal end of protopod acute; rami subequal, broadly subovate; exopod approximately twice as long as broad, with lateral margin straight ending distolaterally into ‘trispinose’ arrangement of two strong teeth – acute lateral and subacute mesial one, with slender and longer movable spine between teeth; mesial distolateral tooth situated laterally on diaeresis, latter sinuous, with obtuse median projection and subacute projection mesially.
Color pattern
BODY ( Fig. 7 View Fig ). Transparent to semi-transparent with one broad reddish brown coloured stripe mid-dorsally on carapace and four similar irregular longitudinal stripes on the sides, all continuing posteriorly to end of sixth pleonite; lateral pleonal stripes regular in shape. Reddish-brown broad stripes interspaced with narrow whitish longitudinal paths with series of white dots in midline. Outer antennular flagella bright red, inner flagellum pinkish; third maxillipeds and pereiopods pinkish proximally with gentle gradient toward distal parts, darker in articulations, distal margin of palm and fingers of first pereiopods pinkish red; uropods and telson reddish brown without any stripes; eggs kiwi-fruit-flesh green.
Remarks
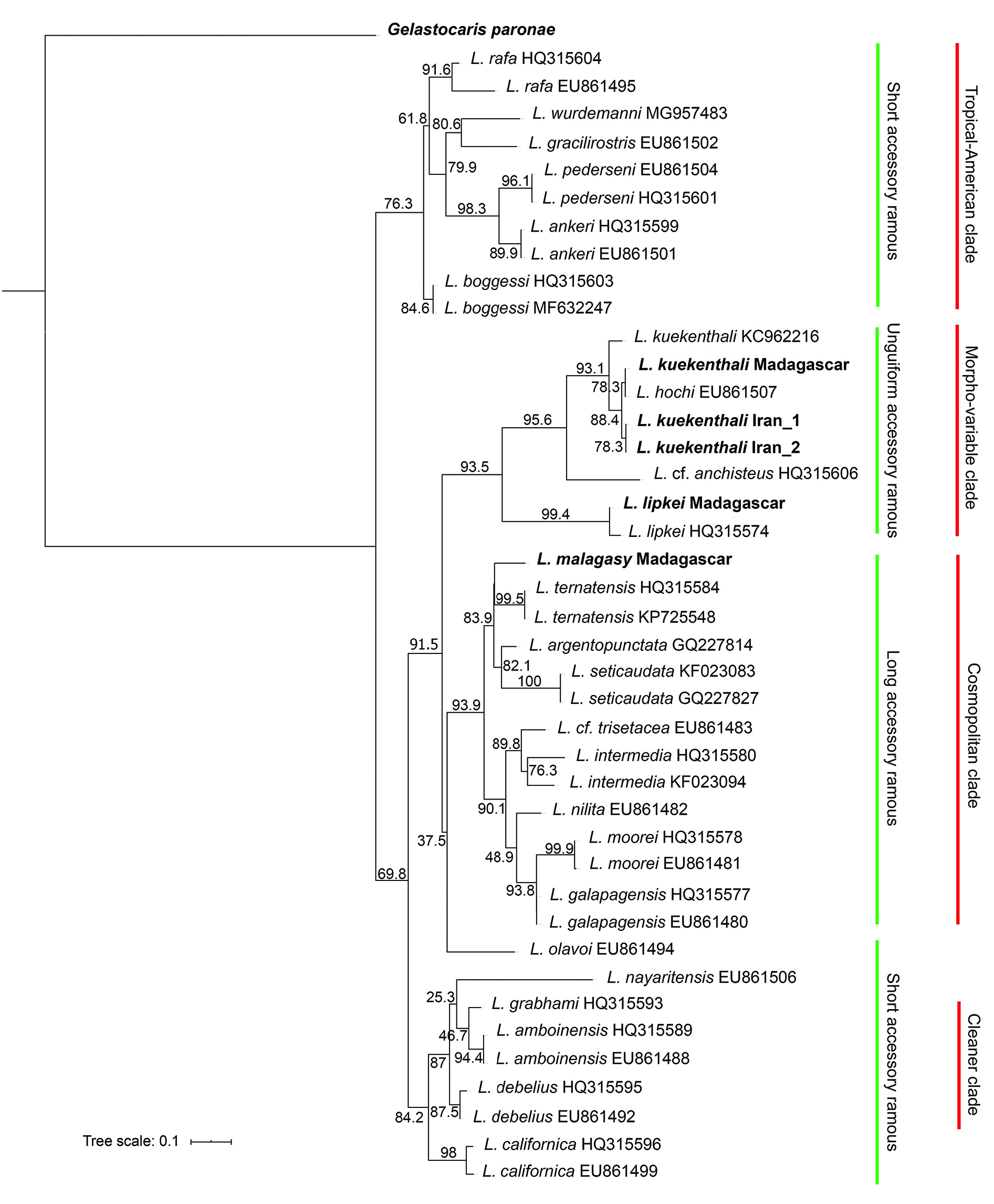
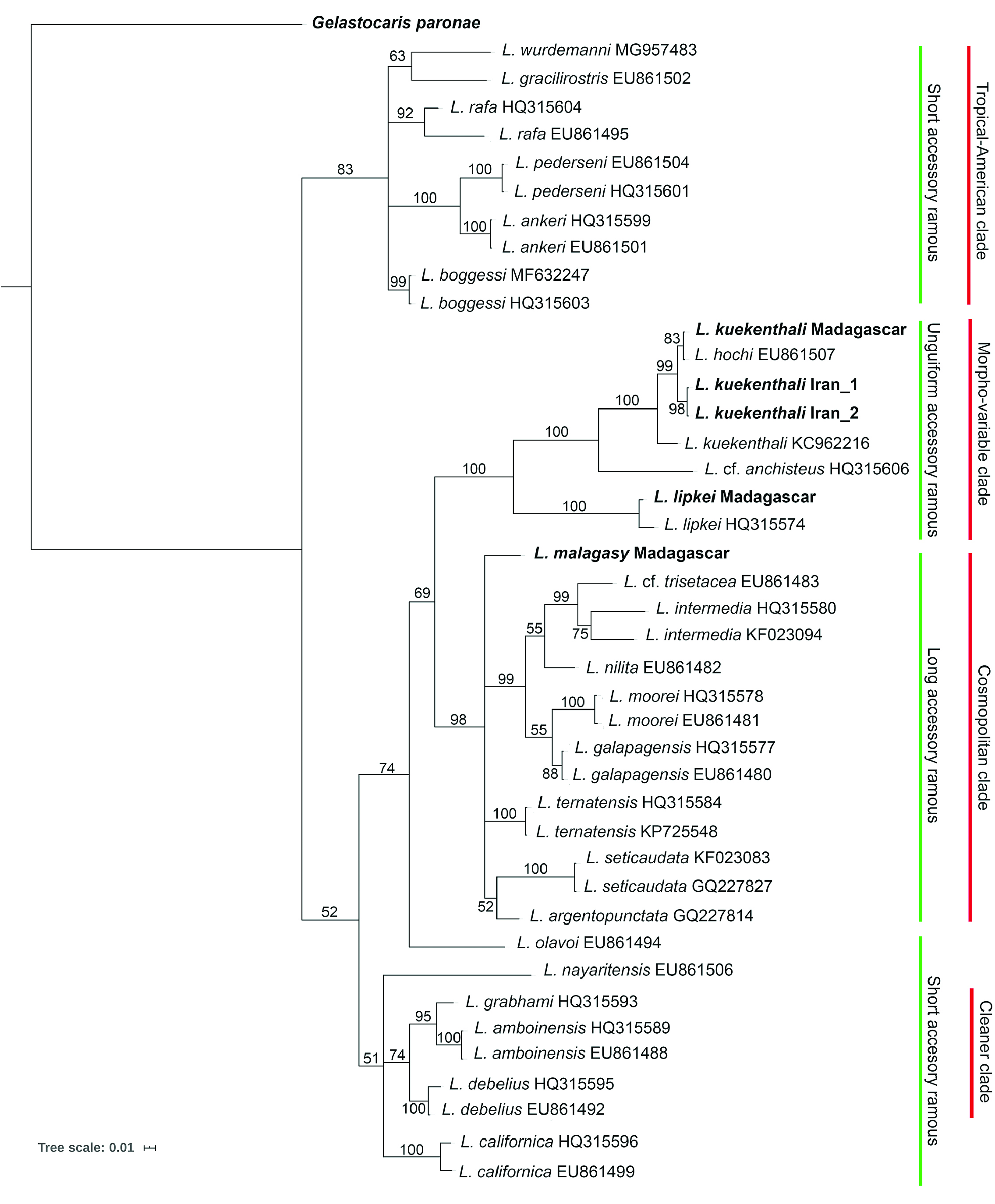
The phylogenetic analysis places the present new species into the “long accessory ramous” clade of the genus Lysmata in both ML ( Fig. 1 View Fig ) and BI ( Fig. 2 View Fig ) analyses. In the ML analysis, L. malagasy sp. nov. forms a subclade; however, not well-supported, together with L. ternatensis , L. argentopunctata and L. seticaudata , with L. ternatensis being the closest species to the new species. In the BI analysis, the relationships of the new species to other species belonging to the “long accessory ramous” clade is not clear as L. malagasy sp. nov. forms no subclade with any other species of the clade.
Considering the division of the genus into three clades morphologically distinguishable by the state of the shorter free (= accessory) ramus of the lateral antennular flagellum i.e., short with one article, unguiform with one finger-like article, or long with multiple articles ( Fiedler et al. 2010), the new species L. malagasy sp. nov. belongs to the third group, i.e., the “long accessory ramous” which represents a well-supported monophyletic clade in previous phylogenetic studies ( Baeza et al. 2009; Baeza 2010; Fiedler et al. 2010; Baeza & Fuentes 2013). In the study of Baeza et al. (2009), Baeza (2010), and Baeza & Fuentes (2013), this clade was named the “cosmopolitan clade”. The Atlantic representatives of this clade are Lysmata intermedia , L. jundalini Rhyne, Calado & dos Santos, 2012 , L. moorei , L. napoleoni De Grave & Anker, 2018 , L. nilita , and L. seticaudata . The eastern-Pacific representatives of this group are L. argentopunctata , L. chica Wicksten, 2000 , L. galapagensis , and L. holthuisi Anker, Baeza & De Grave, 2009 . However, only two species with long accessory ramus have been described from the Indo-West Pacific, L. ternatensis De Man, 1902 , and L. trisetacea , in addition to L. zacae Armstrong, 1941 only being reported from the West Pacific.
Lysmata ternatensis shares with L. malagasy sp. nov. some traits like the rostral armament, i.e., 5–6 teeth on upper (including postrostral ones) and 2–3 on lower margins, the carapace with the pterygostomial tooth, the stylocerite not extending beyond midlength of the second antennular article, and the second pereiopod with the carpus composed of about 24 articles. Although no information about the carpal and meral spinulation of the three last pereiopods was provided by De Man (1902), it seems that these two species are similar in the latter character according to Xu & Li (2015). Importantly, as discussed by De Grave & Anker (2018), and according to personal observations by one of the authors (HA) on other congeners, i.e., L. vittata and L. kuekenthali , characters like the rostral armament, second pereiopod carpal segmentation, and carpal and meral spinulation on three last pereiopods, especially the third one, can be highly variable within a species and consequently untrustworthy for separating taxa. However, based on the original description ( De Man 1902) and other published reports on L. ternatensis (e.g., Chace 1997; Xu & Li 2015; Madhavan et al. 2019), the accessory ramus of the lateral antennular flagellum in L. ternatensis consists of 10 segments (10 to 15 segments were reported for material from the China Sea by Xu & Li in 2015), thus their distinctly higher number clearly distinguishes L. ternatensis from L. malagasy sp. nov. In the latter, the accessory ramus consists of only four elongated articles. This can be the most reliable morphological character applicable to separate these two species. This difference can also be applied for distinguishing L. malagasy sp. nov. from the synonymized species under the name of L. ternatensis , Hippolysmata acicula Rathbun, 1906 , Lysmata affinis Borradaile, 1915 , and Palaemon dentatus De Haan, 1841 . In the description of Hippolysmata acicula, Rathbun (1906) stated nothing about the state of the accessory ramus of the lateral antennular flagellum. However, the holotype material was re-examined by Chace (1997), who reported that the accessory ramus was comprised of 10 segments compared to four in the new species. In the description of Lysmata affinis, Borradaile (1915) only stated that L. affinis is related to L. seticaudata (Risso, 1816) and L. chiltoni Kemp, 1914 ; with no more information about the status of the antennular flagellum. However, Borradaile (1917) provided more information about L. affinis , and he described the accessory ramus of the antennular flagellum being as long as the fused part. This character can be used to distinguish between L. affinis and L. malagasy sp. nov. as in the latter one, the fused part of the antennular flagellum is much longer than the accessory ramus. Holthuis (1947) stated that in L. dentata (originally as Palaemon dentatus ), the fused part and accessory ramus of the antennular flagellum are equal in size as in L. affinis . Thus, the same argument can be used to differentiate between L. dentata and L. malagasy sp. nov.
Two differences are notable between L. trisetacea and L. malagasy sp. nov: 1) the pterygostomial angle is without a tooth in the former species while the latter bears a small acute tooth there; and 2) the accessory ramus of the outer antennular flagellum of L. trisetacea consists of at least eight segments ( Kemp 1914; Chace 1997) and is subequal in length to the fused portion of the flagellum ( Heller 1862; Kemp 1914; Holthuis 1947) whereas in the new species, the accessory ramus includes four elongated segments and is noticeably shorter in length than the fused part. However, it is worth to notice that L. trisetacea Heller, 1861 , and its synonyms L. pusilla Heller, 1862 , and L. chiltoni Kemp, 1914 , should be treated more carefully as there is a considerable difference between the proportion of the length of the free accessory ramus to that of the fused part of the outer flagellum. In the description of L. pusilla, Heller (1862) noted that the accessory ramus is about twice as long as the fused part while Kemp (1914) described L. chiltoni with the fused part and accessory ramus being equal in length.
Armstrong (1941), in the description of L. zacae , only mentioned that the lateral antennular flagellum is biramous. Okuno (1996) re-examined the type material of L. zacae , and yet did not describe the exact status of the lateral antennular flagellum. There are three characters which can be used to differentiate the new species from L. zacae as follows: first, the shape of the pterygostomial angle bearing a small tooth in L. malagasy sp. nov. vs rounded in L. zacae ; second, the very short stylocerite in L. zacae reaching to the proximal third of the basal antennular segment compared to longer one in the new species overreaching the basal antennular segment; and third, the scaphocerite reaching to the end of the antennular peduncle in L. zacae while it distinctly surpassing the antennular peduncle in L. malagasy sp. nov.
Other members of the “long accessory ramous” clade of the genus Lysmata from other geographical regions can be more or less distinguishable from the new species by the number of segments of the fused part and of the accessory ramus of the lateral flagellum; L. intermedia with 17–18 and 3–4 articles ( d’Udekem d’Acoz 2000), L. jundalini 18–24 and 3–4 ( Rhyne et al. 2012), L. moorei 7–13 and 7–16 ( Rathbun 1901; Pachelle et al. 2020), L. napoleoni 6–9 and 10–13 ( De Grave & Anker 2018), L. nilita 21–27 and 4–7 ( Dohrn & Holthuis 1950), L. seticaudata 20–38 and 8–15 ( Dohrn & Holthuis 1950), L. argentopunctata 13–15 and 13–17 ( Wicksten 2000), L. chica 10–11 and 3–6 ( Wicksten 2000), L. galapagensis 6–13 and 5–7 ( Schmitt 1924), and L. holthuisi 17 and 6–7 articles ( Anker et al. 2009), respectively.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |