
Kalophrynus cryptophonus, Vassilieva, Anna B., Galoyan, Eduard A., Gogoleva, Svetlana S. & Poyarkov, Nikolay A., 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3796.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:FE38169D-6687-42DF-B29B-444A954D1E08 |
|
DOI |
https://doi.org/10.5281/zenodo.5674283 |
|
persistent identifier |
https://treatment.plazi.org/id/18098786-FFF8-9618-EDC1-750CE4242811 |
|
treatment provided by |
Plazi |
|
scientific name |
Kalophrynus cryptophonus |
| status |
sp. nov. |
Kalophrynus cryptophonus View in CoL sp. nov.
Holotype. ZMMU A- 4944 (field number ABV-00222), adult male from Loc Bao, Lam Dong Province, Vietnam (coordinates 11°44' 17" N, 107° 42' 25" E, elevation 800 m. a.s.l.), collected by E.A. Galoyan and A.B. Vassilieva on 22 April 2013.
Paratypes. ZMMU A- 4858, one adult female (individual field number ABV-00136) collected on 12 of April 2013, and three adult males (individual field numbers ABV-00224, ABV-00225, ABV-00226) collected on 22 April 2013 in the same area as holotype; ZMMU A- 4859, one adult male (individual field number ABV-00223), collected on 22 April 2013 in the same area as holotype.
Diagnosis. The species is allocated to Kalophrynus based on the following characters considered diagnostic for the genus ( Parker 1934; Bourret 1942; Inger 1966): one or more transverse dermal ridges across the palate anteriorly to the oesophagus; tips of digits not dilated to disks; no spine-like projections at heel or elbow; snout short, pointed; tympanum visible; pupil horizontal; inner metatarsal tubercle low, not shovel-like; skin thick and glandular. The new species is distinguishable from its congeners by a combination of the following features: (1) small size, SVL 27.9–30.4 mm in males, 23.4 mm in female; (2) snout pointed, slightly sloping in profile; (3) canthus rostralis indistinct; (4) males with large sharp conical spines on the skin covering the jaws; (5) males with finely asperous nuptial pads on the dorsal surface of the fingers I–III; (6) tympanum distinct, smaller than eye in diameter; (7) toe webbing moderate; (8) outer metatarsal tubercle present; (9) light dorsolateral line absent; (10) dark ocellus in the inguinal region usually present, small, without light bordering; (11) anterior palatal dermal ridge short, restricted to medial part of the palate.
Description of holotype. Adult male ( Fig. 3 View FIGURE 3 AB) with well-developed oblong testes; measurements are given in Table 4.
Habitus stout, with body widest in lumbar area. Head relatively short (HL/SVL 0.31), slightly wider than long (HL/HW 0.88), triangular. Snout pointed, short (SL/HL 0.36), sloping in profile ( Fig. 3 View FIGURE 3 C) and distinctly projecting beyond lower jaw; canthus rostralis indistinct, smooth; nostrils rounded, situated closer to snout tip than to eye (SND/ END 0.65) and oriented rostrolaterally. Eyes bulging, medium-sized (ED/SVL 0.11), lateral, well visible from below, with oval horizontal pupil; upper eyelid with fine tubercles on margin, UEW/IOD 0.45. Tympanum very close to eye, smaller than eye (TD/ED 0.69, TD/SVL 0.08), roughly round, distinct, bordered from above by sharply prominent supratympanic fold running from posterior corner of eye along upper edge of tympanum and then down to the point of forelimb insertion. Maxillary and vomerine teeth absent; tongue with even rounded edge posteriorly; transversal crenulated dermal ridge on palate posterior to eyes level, short low dermal fold anterior to it, restricted to medial part of palate; postchoanal dermal ridges distinct. Small notch in the rostral part of skin covering of upper jaw and small prominence on the rostral part of lower jaw, fitting together. Single subgular vocal sac; wide slit-like vocal openings between jaw articulation and tongue insertion.
Numerous tubercles with wide base and sharp, conical, semitransparent apical spine being clearly distinct on skin covering maxilla and mandible ( Fig. 3 View FIGURE 3 D). Spines on mandible largest, bordering the lower jaw in one to three rows, from rostral end to mouth corner and spreading on the area below and behind tympanum; on upper jaw spines smaller and occupying the entire area below snout tip, nostrils and eyes. Several tiny, hardly distinct similar spines present on each tympanum. Small scattered spines spread on axillar area, on anterior forelimb surface and on breast between forelimbs, but not on gular region.
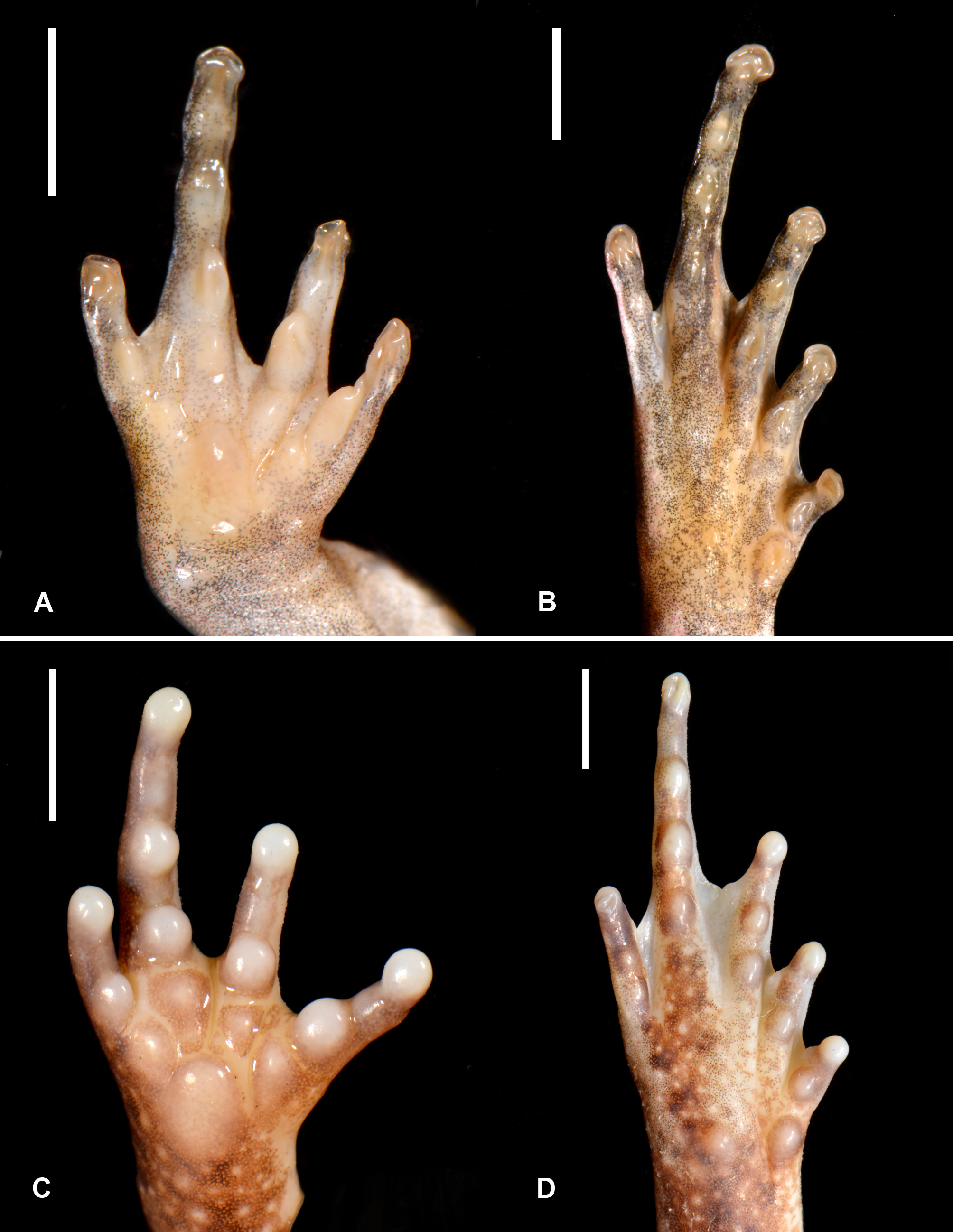
Forelimbs slender, more than half of body length (FLL/SVL 0.71); hands with rudimental webbing. Tips of fingers rounded, without dilatation. Relative length of fingers I<IV<II<III; palmar tubercle moderately large, prominent, oval in shape; subarticular tubercles prominent, rounded; one on each fingers I, II and IV and two on finger III; smaller additional metacarpal tubercles between palmar tubercle and each finger base ( Fig. 4 View FIGURE 4 A).
Nuptial pads on dorsal hand well-defined, finely asperous, thickened areas of skin covering fingers I–III from base to proximal end of terminal phalange ( Fig. 4 View FIGURE 4 C).
Hind limbs slender, relatively short (HLL/SVL 1.36); when legs adpressed to body laterally, heel reaches interval between eye and tympanum. Tibia slightly shorter than thigh (TFL/ThL 0.96) and significantly shorter than tarsus+foot (TFL/TarFL 0.66). Toe tips rounded, without dilatation; relative length of toes I<II<V<III<IV; foot webbing formula I 1–2 ½ II ½–2½ III 1 ½ –3½ IV 3 ¾–1½ V; subarticular tubercles moderately prominent on toes I–IV, oval in shape; one on each toe I and II, two on toe III and three on toe IV; distal subarticular tubercle on toe V distinct, proximal one less conspicuous, rudimentary; inner metatarsal tubercle prominent, oval; outer metatarsal tubercle smaller than inner one (OMTL/IMTL 0.8), low, rounded ( Fig. 4 View FIGURE 4 B).
Skin on dorsum finely shagreened from interorbital level to vent, with singular small tubercles topped with tiny apical spine; smooth on the snout; roughly granular on belly, lower flanks, thighs undersides and in cloacal region, smooth on gular area, inner leg surfaces and forelimbs except for scarce, tiny spines in outer and axillar surfaces. Two pairs of whitish glandular tubercles on the pectoral region, each with tiny sharp spines on apex. Dorsolateral boundary sharp, finely glandular.
Coloration. In life,( Fig. 5 View FIGURE 5 AB) ground coloration on upper head, trunk and limb surfaces variable, ranging from chocolate-brown at night to pinkish-beige during the day time. Sharp edge of dorsal coloration extending from snout tip through upper eyelid to groin contrasted by darker, pinkish-brown flanks and head sides. Diffuse pinkish-gray pattern in the shape of inverted “ Y ” on the dorsum lasting from interorbital area towards inguinal region and edged with light-beige color. " Y "-shaped pattern followed laterally by three less distinct, parallel, intervening stripes forming somewhat reticulate pattern. Pale rounded spot visible on parietal area, similar diffuse spots scattered on dorsum. Three faint brownish crossbands on thigh and shin; brown marmorate pattern on elbow and knee, on foot underside and on anterior surface of lower forearm and hand. Cloacal region brownish, separated from dorsal surface by thin pale edging line. Inguinal ocelli represented by small (smaller than tympanum, ISD/TD 0.77) rounded black spot in each inguinal area, without light edges; three asymmetric small dark dots on left side of back. Venter pale, yellowish-pink, unpatterned except for gular, chest and upper belly areas displaying diffuse brownish-gray spotting; palmar and plantar surfaces pink. Iris golden.
In preservative coloration fades to gray on dorsum and flanks and yellowish-gray on venter; other features remain without significant change.
Variation. Individuals of the type series are similar in morphology, body size and proportions (Table 4; Fig. 3 View FIGURE 3 E), except for the female that is smaller in size and lacks the spines and the asperous patches on the hand. The spines in males vary slightly in abundance and size; in all individuals they are the largest on mandible margins, especially in the subtympanal area, and may be less conspicuous on the upper jaw. Nuptial pads on fingers I–III are similar in all males. Variations in the coloration include the inverted “ Y ” or hourglass-shaped pattern on dorsum, which may be almost indistinct; three paratypes have a uniform stellar, slightly paler pattern on back. The inguinal ocelli vary in size, but are always smaller in diameter than tympanum (ISD/TD 0.77±0.05); in the female this spot is present only on the left side of body. Supernumerary asymmetric small dark spots may occur in the inguinal area, on back and on upper thigh and shin surface. The female ( Fig. 5 View FIGURE 5 CD) additionally differs in undersides coloration by having a more pronounced dark marbling or spotting on the upper belly and lower part of flanks, and by having a light medial band on the gular surface bordered with diffuse brown blotches. The ovaries of the female contain relatively small quantity (50–60 per ovary) of developing unpigmented eggs variable in size (diameter 0.3–0.9 mm).
Natural history. All specimens were collected at night time on the limited parcel of secondary, disturbed mountain high polydominant evergreen tropical forest with the abundance of bamboo Phyllostachys sp. ( Poaceae ) ( Fig. 6 View FIGURE 6 ), where Kalophrynus cryptophonus sp. nov. occurs syntopically with Kurixalus sp., Theloderma bambusicola Orlov, Poyarkov, Vassilieva, Ananjeva, Nguyen, Sang & Geissler , Microhyla fissipes Boulenger , Limnonectes limborgi (Sclater) , and Ingerophrynus galeatus (Günther) . The female was found on the ground; all males were collected while calling from inside dead bamboo stems with diameters of about 30–40 mm ( Fig. 5 View FIGURE 5 E), sometimes as close to each other as 5–10 m; up to four males were heard simultaneously from one point. Tadpoles were collected on 22 April 2013 in a decaying bamboo stem with diameter about 35 mm. In total, 11 early-stage larvae (Gosner Stage 25) were recorded in one partly perforated internode, containing 70–80 ml of rainwater. A calling male was found in the adjacent internode of the same stem.
Tadpole description. Tadpoles were allocated to the species based on the following evidence: (1) morphological features characteristic for microhylid larvae in general and other phytotelm breeding Kalophrynus species in particular (see Discussion); (2) specific site of collection (water-filled decaying bamboo internode) which was close to calling male found in the same bamboo stem; (3) all other phytotelm breeding frogs known from the region (members of the genera Theloderma and Rhacophorus , Rhacophoridae ) differ in tadpole morphology and cannot be mistaken with larvae of the new species.
Standard tadpoles measurements (mean±SD, n=4, Stage 25): TL 9.14±0.77; BL 2.50±0.05; BW 1.47±0.03; SVL 3.02±0.16; SSp 1.89±0.15; IOD1.05±0.03; SED 0.98±0.03; ED= 0.16±0.01; ODW= 0.94±0.06; TaL= 6.6±0.72; TH= 0.89±0.1.
In dorsal view ( Fig. 7 View FIGURE 7 A), body oval or slightly pyriform, with maximum width at eye level (BW/BL 0.59±0.01); snout long (SED/BL 0.39±0.01), broad, blunt. In lateral view ( Fig. 7 View FIGURE 7 B), body compressed dorsoventrally (BH/BW 0.66±0.05), especially anteriorly, flattened above and convex below. Tail long (TaL/ BL2.70±0.25), lanceolate; almost equal in height along its length; tail tip bluntly acuminate, without terminal filament; muscular part moderately developed; at vent level, tail muscular portion about three times higher than lower fin. Upper fin originating slightly behind body-tail junction, much shorter than the lower fin proximally and nearly equal in height to it distally. Eyes dorsolateral, not visible from below, small (ED/BL 0.06±0.002), pupils oriented dorsolaterally. Nostrils not opened at the stages under description. Spiracle ventral, medial, with short free flap, opening at 0.80±0.06 of distance from snout tip to body end. Vent tube medial, long, directed obliquely backward, with opening at ventral edge of lower fin. Lateral line system not visible. Mouth terminal, oral disk wide (ODW/BW 0.64±0.05), without keratinized elements and obvious papillae. Upper labium widely arched, slightly overhanging Ω-shaped lower labium.
Live tadpoles appeared almost unpigmented, whitish, with darker gut and reddish gills clearly visible through translucent skin. In preservative, scattered melanophores visible on the dorsal surface of body and tail. Belly and tail fins totally transparent. Eyes black. Gut forming three wide loops.
Call characteristics. The advertisement call of Kalophrynus cryptophonus sp. nov. recorded at 25ºC is represented by rhythmic, non-modulated, tonal calls ( Fig. 8 View FIGURE 8 A).
Call duration varied from 42 to 114 ms (79±13ms, n=105). The interval between successive calls in one individual varied from 5.91 to 25.57 s (11.76± 6.57 s, n=10). In two individuals calling simultaneously at the distance 5 m apart from each other the interval between calls uttered by different males varied from 0.009 to 2.37 s (1.06± 0.56 s, n=31). The call repetition rate in individuals (n=11) varied from 0.016 to 0.42 calls per second (0.13±0.12, n=105 calls). The call repetition rate in choruses (n=3) consisting of 4–5 individuals varied from 0.21 to 0.74 calls per seconds (0.44±0.27). The initial (1020±100 Hz) and final fundamental frequency in a single call (920±130 Hz) were almost equal (n=38). Two lower harmonics were distinguishable on the majority of call spectrograms. The maximum amplitude frequency in individual calls (1100±370 Hz, n=38) coincided with the fundamental frequency. The entire call was pulsed, with2–5 pulses (3.270.69, n=52) per call and a mean pulse period of 24±0.3 ms. In all calls recorded the pulse rate varied from 30.3 to 55.6 pulses per second (42.51±6.29 pulses/s, n=52).
Comparison. In southern Vietnam, Kalophrynus cryptophonus sp. nov. co-occurs with Kalophrynus interlineatus , which inhabits diverse kinds of forested biotopes at elevations up to 900 m a.s.l. The latter was reliably recorded from the same province and district as Kalophrynus cryptophonus sp. nov., and both species may potentially occur syntopically with each other. From southern Vietnamese K. interlineatus , the new species differs morphologically by the following features: a smaller size (SVL 23.4–30.4 mm vs. 38.6–45.4 mm); the presence of skin spines on head and nuptial pads on fingers in males (absent in K. interlineatus ); smooth canthus rostralis (vs.
sharp in K. interlineatus ); skin on gular area smooth (roughly tuberculated in K. interlineatus ); longer legs (HLL/ SVL 1.35±0.05 vs. 1.16± 0.07 in K. interlineatus ), in adpressed leg heel reaching tympanum (vs. not reaching tympanum in K. interlineatus ); the presence of rudimental webbing on fingers (absent in K. interlineatus ) and slightly less extensive webbing on toes (webbing formula I 1–2 ½ II ½–2½ III 1 ½ –3½ IV 3 ¾–1½ V vs. I ½ –2 II 1–2 ½ III 1–3 IV 3 ¾– 1 V in K. interlineatus , Fig. 10 View FIGURE 10 CD), and reduced anterior palatal dermal ridge (well developed, arch-shaped in K. interlineatus ). In coloration, Kalophrynus cryptophonus sp. nov. differs from K. interlineatus by having a very small, rudimentary black spot without light edging in inguinal area (smaller than tympanum, ISD/TD 0.77±0.05 vs. larger than tympanum, ISD/TD 1.60±0.36), and white-edged inguinal ocellus in K. interlineatus . In reproductive characteristics, Kalophrynus cryptophonus sp. nov. differs from K. interlineatus by having unpigmented eggs (vs. pigmented in K. interlineatus ). In vocalization, Kalophrynus cryptophonus sp. nov. differs from K. interlineatus by calling from inside bamboo stems (vs. from forest floor, often in dense choruses around breeding ponds), as well as call duration, inter-call interval, frequency and other acoustic parameters ( Fig. 8 View FIGURE 8 AB, Table5).
From its other congener, described herein from southern Vietnam, Kalophrynus honbaensis sp. nov. (see below), Kalophrynus cryptophonus sp. nov. differs morphologically by having a smaller body size in males (SVL 27.9–30.4 mm vs. 26.7–36.8 mm in Kalophrynus honbaensis sp. nov.); large skin spines on head (vs. only microscopic spines in Kalophrynus honbaensis sp. nov.), nuptial pads on fingers in males present (absent in males Kalophrynus honbaensis sp. nov.); a smooth canthus rostralis (vs. sharp in Kalophrynus honbaensis sp. nov.); reduced anterior palatal dermal ridge (vs. well developed, parallel to posterior ridge in Kalophrynus honbaensis sp. nov.), and more extensive webbing on toes (webbing formula I 1–2 ½ II ½–2½ III 1 ½ –3½ IV 3 ¾–1½ V vs. I 1–2 II 1 ½ –3 III 2–3 ¾ IV 4–2 V). In coloration, Kalophrynus cryptophonus sp. nov. differs from Kalophrynus honbaensis sp. nov. by having a very small, rudimentary black spot in inguinal area (smaller than tympanum, ISD/ TD 0.77±0.05, vs. larger than tympanum, ISD/TD 1.47±0.09).
Kalophrynus cryptophonus View in CoL sp. nov. differs from other congeners distributed in South Asia, China, the Malay Peninsula and the Malay Archipelago by the presence of sharp prominent spines on the head skin (not recorded in any other species) in males. Moreover, Kalophrynus cryptophonus View in CoL sp. nov. has substantially smaller body size (SVL 23.4–30.4 mm) than K. baluensis Kiew, 1984 View in CoL (SVL 34.8–39 mm), K. intermedius Inger, 1966 View in CoL (SVL 35–40 mm in females), K. orangensis Dutta, Ahmed & Das, 2000 View in CoL (SVL up to 38 mm), K. palmatissimus Kiew, 1984 (SVL 31–39 mm) and K. pleurostigma Tschudi, 1838 View in CoL (SVL 35–50 mm in males, 38–58 mm in females). Additionally, from these species Kalophrynus cryptophonus View in CoL sp. nov. differs by having a single, small black inguinal ocellus not edged in white on each side of sacrum (vs. one or two yellow ocelli in K. baluensis View in CoL , absence of inguinal spot in K. intermedius View in CoL , white-edged black ocelli in K. pleurostigma View in CoL ), the presence of rudimental webbing on fingers (absent in K. orangensis View in CoL ) and moderate toe webbing (vs. extensive webbing in K. palmatissimus ), and from K. baluensis View in CoL and K. pleurostigma View in CoL by call characteristics (i.e. call duration, dominant frequency and modulation) (Table 5).
Kalophrynus cryptophonus View in CoL sp. nov. differs from K. barioensis Matsui & Nishikawa, 2011 View in CoL (SVL 17.5–19.8 mm in males, 20.5 mm in females), K. nubicola Dring, 1983 View in CoL (SVL 14.4–24.4 mm in males) and K. robinsoni Smith, 1922 View in CoL (SVL 17 mm in males, 18 mm in females) in having a significantly larger body size (SVL 27.9–30.4 mm in males, 23.4 mm in female); additionally, from these species Kalophrynus cryptophonus View in CoL sp. nov. differs by the presence of finely asperous nuptial pads on fingers (absent in K. barioensis View in CoL and K. nubicola View in CoL , few large spines in K. robinsoni View in CoL ), presence of outer metatarsal tubercle (vs. absent in K. barioensis View in CoL and K. nubicola View in CoL ), coloration features (presence of a light lateral stripe in K. barioensis View in CoL , no inguinal ocellus in K. nubicola View in CoL and K. robinsoni View in CoL ), and from K. barioensis View in CoL and K. nubicola View in CoL by call characteristics, namely a lower dominant frequency (Table 5).
From Kalophrynus View in CoL species, which are similar in size, Kalophrynus cryptophonus View in CoL sp. nov. clearly differs by the following important features: from K. bunguranus Günther, 1895 View in CoL (SVL 22.2–23.4 mm in males, 25.5–26.7 mm in females) by having small inguinal spots not edged in white (vs. dark ocelli surrounded by a light area in K. bunguranus View in CoL ); from K. calciphilus Dehling, 2011 View in CoL (SVL 29.7–30.1 mm in males, 35.5–38.8 mm in females) by the absence of dentition on the upper jaw (vs. weak dentition present in K. calciphilus View in CoL ), by finger IV being longer than I (vs. IV shorter than I in K. calciphilus View in CoL ), by coloration features (black dorsum and chest, white dorsolateral line, red iris in K. calciphilus View in CoL ), by call characteristics, in particular call duration and dominant frequency (Table 5); from K. eok Das & Haas, 2003 View in CoL (SVL 26.3 mm in male), by coloration features (dark interorbital bar and chevron on shoulders, inguinal spots absent in K. eok View in CoL ); from K. heterochirus Boulenger, 1900 (SVL 26 mm in males, 30 mm in female) by normally developed fingers (vs. strongly shortened fingers I, II and IV in K. heterochirus ) and small black inguinal spot (vs. few large round yellowish-white spots in the lumbar region in K. heterochirus ); from K. limbooliati Matsui, Nishikawa, Belabut, Norhayati & Yong, 2012 View in CoL (SVL 26.2–28.7 mm in males) by the presence of subarticular tubercles under toe V and the outer metatarsal tubercle (absent in K. limbooliati View in CoL ), the absence of light dorsolateral line (present in K. limbooliati View in CoL ) and call characteristics, in particular dominant frequency and frequency modulation (Table 5); from K. menglienicus Yang & Su, 1980 View in CoL (SVL 19.75–23.4 mm) by moderate webbing on toes (vs. foot webbing absent in K. menglienicus View in CoL ) and the absence of horny granules on back and belly (present in K. menglienicus View in CoL ); from K. minusculus Iskandar, 1998 View in CoL (SVL 25 mm in males, 35 mm in females) by coloration features (brownish black coloration with black banded pattern in K. minusculus View in CoL ); from K. punctatus Peters, 1871 View in CoL (SVL 22–27 mm in males) by the presence of outer metatarsal tubercle (absent in K. punctatus View in CoL ); from K. stellatus Stejneger, 1908 View in CoL (SVL 24 mm) by reduced anterior palatal ridge (vs. developed arched ridge in K. stellatus View in CoL ) and inguinal spots smaller than tympanum and not edged in white (vs. black inguinal spots as large as tympanum and surrounded with light dots in K. stellatus View in CoL ); from K. subterrestris Inger, 1966 View in CoL (SVL 23 mm in males, 27 mm in females) by indistinct canthus rostralis (distinct in K. subterrestris View in CoL ), reduced anterior palatal ridge (vs. developed angular, strongly notched ridge in K. subterrestris View in CoL ) and subarticular tubercles on toe V present (absent in K. subterrestris View in CoL ); from K. tiomanensis Chan, Grismer & Grismer, 2011 View in CoL (SVL 25.8–26.3 mm in males, 21.4–25.8 mm in females) by the presence of outer metatarsal tubercle (absent in K. tiomanensis View in CoL ), small inguinal spot (vs. large spot in K. tiomanensis View in CoL ) and other coloration features (spotted lower back in K. tiomanensis View in CoL ); from K. yongi Matsui, 2009 View in CoL (SVL 28.8–31.0 mm in males) by the absence of strongly developed humeral spine (present in males K. yongi View in CoL ), finely asperous nuptial pads on fingers I–III (vs. sharp white asperities on nuptial pads in males K. yongi View in CoL ), tadpole features (see Discussion) and call characteristics, namely call duration, dominant frequency and modulation (Table 5).
Distribution. To date the species is known only from type locality, the Loc Bao Forestry Enterprise in Lam Dong Province.
Etymology. The species name is a noun in the nominative case, derived from the Greek cryptos, meaning “to hide”, “mystery”, “cryptic”, and phonus meaning “voice” in reference to our history of discovering the species. During the expedition of the JRVTRTC in April–May, 2013 in Loc Bao forest, Lam Dong Province, we recorded unusual frog calls which were hard for us to locate. After two weeks of searching we found a male Kalophrynus cryptophonus sp. nov. calling from an empty bamboo stem.
Recommended vernacular name: The recommended common name in English is Bamboo sticky frog, referring to the unusual breeding habitat of the new species.






| ZMMU |
Zoological Museum, Moscow Lomonosov State University |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Kalophrynus cryptophonus
| Vassilieva, Anna B., Galoyan, Eduard A., Gogoleva, Svetlana S. & Poyarkov, Nikolay A. 2014 |
K. limbooliati
| Matsui, Nishikawa, Belabut, Norhayati & Yong 2012 |
K. barioensis
| Matsui & Nishikawa 2011 |
K. calciphilus
| Dehling 2011 |
K. tiomanensis
| Chan, Grismer & Grismer 2011 |
K. yongi
| Matsui 2009 |
K. eok
| Das & Haas 2003 |
K. orangensis
| Dutta, Ahmed & Das 2000 |
K. minusculus
| Iskandar 1998 |
K. baluensis
| Kiew 1984 |
K. palmatissimus
| Kiew 1984 |
K. nubicola
| Dring 1983 |
K. menglienicus
| Yang & Su 1980 |
K. intermedius
| Inger 1966 |
K. subterrestris
| Inger 1966 |
K. robinsoni
| Smith 1922 |
K. stellatus
| Stejneger 1908 |
K. heterochirus
| Boulenger 1900 |
K. bunguranus Günther, 1895
| Gunther 1895 |
K. punctatus
| Peters 1871 |
K. pleurostigma
| Tschudi 1838 |