
Aquilonastra O’Loughlin, 2004
|
publication ID |
https://doi.org/ 10.24199/j.mmv.2004.61.1 |
|
DOI |
https://doi.org/10.5281/zenodo.10870627 |
|
persistent identifier |
https://treatment.plazi.org/id/185387DD-FFB6-FFB8-FF0E-E570FB0949E0 |
|
treatment provided by |
Felipe |
|
scientific name |
Aquilonastra O’Loughlin |
| status |
gen. nov. |
Aquilonastra O’Loughlin View in CoL gen. nov.
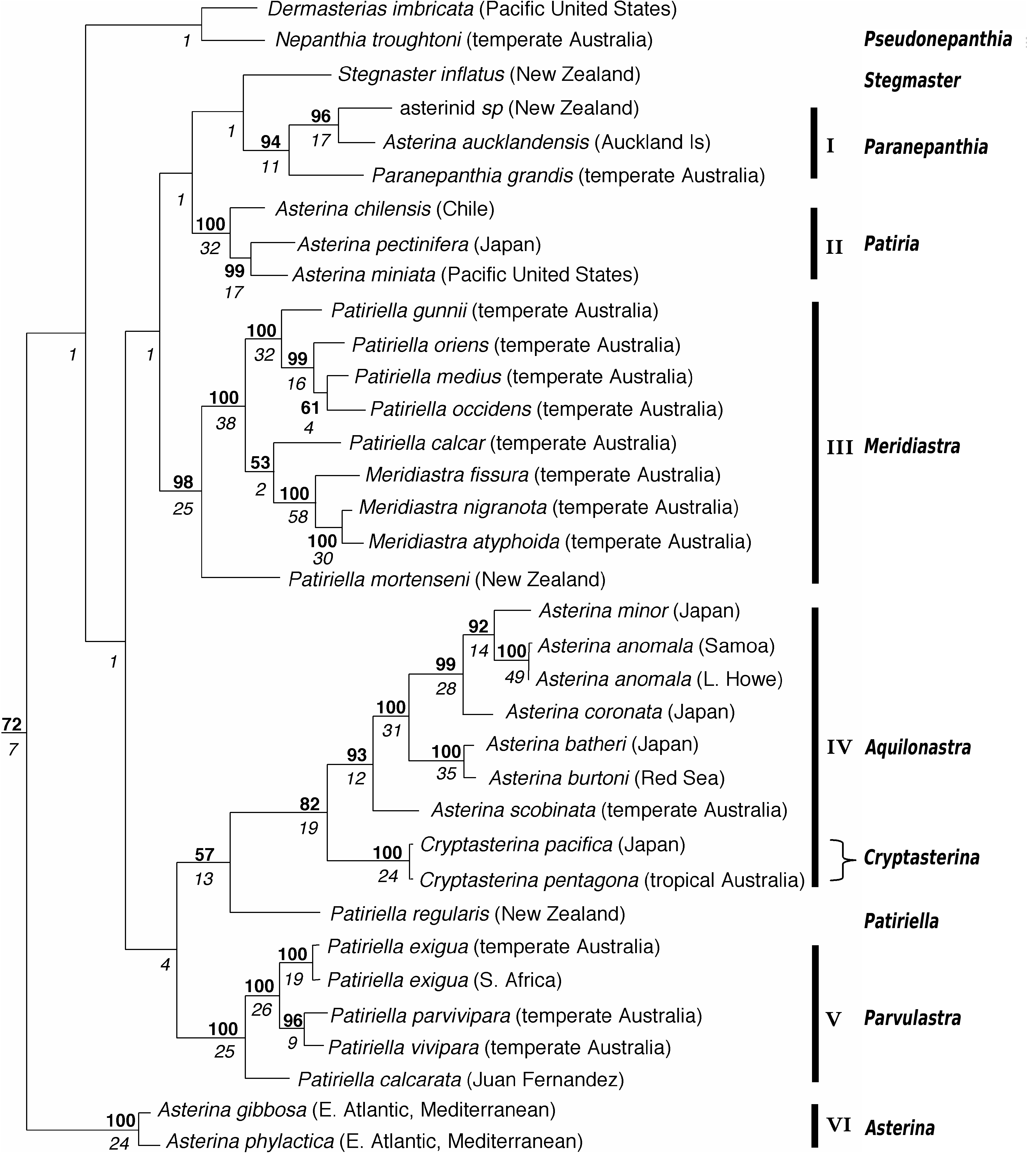
Figures 1 View Figure 1 (clade IV, part), 2b, 9a–f
Diagnosis. Rays 5 (6–8 in fissiparous species); interradial margin deeply incurved, form stellate; rays discrete, broad at base, tapering, rounded distally; flat actinally, high convex abactinally; abactinal plates in longitudinal series, not perpendicular to margin; papulate areas extensive; papulae predominantly single, large, in longitudinal series along sides of rays; abactinal plates with glassy convexities; abactinal spinelets and actinal spines predominantly fine, glassy, conical or sacciform or splay-pointed sacciform, in bands or tufts, numerous (10–40 per plate); actinal plates in longitudinal, not oblique, series; superambulacral plates present for all of ray, sometimes for part of ray or absent in pedomorphic species; superactinal plates present.
Type species. Asteriscus cepheus Müller and Troschel, 1842 .
Other species. A. anomala (H.L. Clark, 1921) ; A. batheri (Goto, 1914) ; A. burtoni ( Gray, 1840) ; A. corallicola ( Marsh, 1977) ; A. coronata (Martens, 1866) (junior synonyms: Asterina coronata fascicularis Fisher, 1918 , possible synonymy raised by H.L. Clark, 1928, formalised by Rowe, 1995; and Asterina spinigera Koehler, 1911 , by VandenSpiegel et al., 1998); A. heteractis (H.L. Clark, 1938) ; A. iranica ( Mortensen, 1940) (raised to species status, this work); A. limboonkengi ( Smith, 1927); A. minor (Hayashi, 1974) ; A. rosea (H.L. Clark, 1938) ; A. scobinata ( Livingstone, 1933) .
Material examined. A. anomala . Caroline Is, WAM Z6845 (2); Christmas I., Z6851 (1); Lord Howe I., AM J6169 (21); NMV F95593 (6); F97690 (6); F96699 (1); Maldive Is, Z6854 (1); Samoa, F96698 (2); Torres Strait, Darnley I., Z6849 (2); Western Australia, Kimberley, Z6843 (1).
A. batheri . Japan, Toyama Bay, NMV F97441 (1); AM J11564 (2); Oman, UF 70 (7).
A. burtoni . Gulf of Suez, AM J17892 (2); TM H1815 (2); Red Sea, Egyptian coast, H1814 (1).
A. cephea iranica . Syntype. Persian Gulf , S of Bushire, coral reef, AM J17891 (1).
A. cepheus . Queensland, NMV F95594 (2), AM J23331 (1); Western Australia, Abrolhos Is, WAM Z6778 (7); J8321 (5); New Guinea, Trobriand Group, J22934 (1); Hong Kong, BMNH 1981.2.6.25 (1; previously identified as Asterina limboonkengi ).
A. corallicola . Paratypes. Caroline Is, WAM Z1704 About WAM (3).
A. coronata . Northern Territory, Darwin, NMV F95796 (4); AM J6613 (1); J8206 (2); Caroline Is, J13660 (1); Bombay, BMNH 1960.10.4.11-16 (2; previously identified as Asterina lorioli ); Taiwan, J19956 (1); Japan, J11563 (2).
A. heteractis . Queensland, Townsville, AM J9541 (2); Heron I., J19449 (1).
A. limboonkengi . Oman, UF 1645 (2); UF 246 (1); UF 68 (6).
A. minor . Japan, NMV F96697 (2).
Paranepanthia rosea . Paratypes. Western Australia, Rottnest I., AM J6171 (3). Other material. Western Australia, Jurien Bay , AM J7437 (4).
A. scobinata . Holotype. Tasmania, AM J1241 . Other material. Tasmania, Eaglehawk Neck, J9060 (3); Victoria, Phillip I., NMV F72998 About NMV (1); Port Fairy , F72985 (2); Killarney, F72997 (10) .
Description with species variations. Rays predominantly 5, or 6 (fissiparous corallicola , heteractis ), or 6–7 (fissiparous burtoni ), or 7–8 (fissiparous anomala ); interradial margin deeply incurved, form stellate; rays discrete, broad at base, tapering, rounded distally; flat actinally, high convex abactinally; size medium ( iranica up to R = 35 mm) to small (pedomorphic species anomala , burtoni , corallicola , heteractis , minor with R up to 10 mm); pedicellariae sometimes present, scattered ( iranica ) or few on proximal lateral rays above papulae ( batheri , coronata ); gonopores abactinal or actinal ( corallicola , scobinata ); fissiparity in 4 species.
Abactinal surface even, or very uneven with some paxilliform plates ( coronata , rosea ); plates in longitudinal series, not series perpendicular to margin; papulate areas extensive, to near distal end of rays, to near interradial margin; papular spaces small with few secondary plates and predominantly 1 large papula (rarely up to 3); 2–6 distinct longitudinal series of large papulae along each side of rays, series variably evident carinally; disc variably bordered by continuous series of 5 radial and 5 interradial plates (not in fissiparous species); abactinal plates predominantly singly notched for papulae, crescentiform on rays and interradially to near margin, round distally, with transverse or round spinelet-bearing proximal elevations; carinal series of plates absent, or proximal series of singly or doubly notched plates not widely separated by secondary plates, or doubly papulate to end of ray ( heteractis ); cleared abactinal plates with glassy convexities; abactinal spinelets glassy, conical or sacciform or splay-pointed sacciform; abactinal spinelets in transverse bands or tufts of 8–40 spinelets per plate, tufts sometimes multiple per plate ( batheri , cepheus ), spinelets sometimes 2 distinct forms ( coronata ), spinelets sometimes paxilliform ( coronata , rosea ); marginal plates with abactinal spinelets; superomarginal plates not larger than inferomarginals; acute margin formed by projecting inferomarginal plates (rounded margin in pedomorphic corallicola ).
Actinal plates in longitudinal series, sometimes also oblique ( coronata ), or oblique (pedomorphic heteractis ).
Actinal spines per plate: oral 5–9; suboral 3–12; furrow 3–9 proximally; subambulacral 2 up to tuft; adradial actinal plates fully spinous; actinal interradial spines similarly variable forms as abactinally, predominantly in tufts, up to 20 per plate.
Superambulacral plates present for all of ray, or distal half (pedomorphic minor ), or absent (pedomorphic corallicola ); superactinal plates present.
Distribution. Eastern Mediterranean, northern Indian and western Pacific Oceans, Gulf of Suez, Red Sea, Persian Gulf, Arabian Sea, southern Australia, Japan; 0– 15 m.
Etymology. From the Latin aquilonalis (northern) and astrum (star), referring to the predominantly northern Indo-Pacific occurrence of the species of this genus (feminine).
Remarks. The new genus Aquilonastra is represented on the molecular clade of Waters et al. (2004, clade IV, part) by six species: A. anomala , A. batheri , A. burtoni , A. coronata , A. minor , A. scobinata . These share a consistent morphology: discrete high rays; series of single large papulae along rays; fine glassy spinelets; presence of superambulacral and superactinal plates. Other species share this morphology and are included in the genus: cepheus , corallicola , heteractis , limboonkengi and rosea . The new genus shares some characters with Cryptasterina Dartnall et al., 2003 : extensive papulate areas; small papular spaces with few secondary plates and single papulae per space; up to four longitudinal series of papulae along each side of the rays; doubly papulate carinal plates variably present, as short or long series or absent; margin formed by projecting inferomarginal plates; superactinal plates support the interradial margin. This similarity is supported by a sister taxon relationship on the molecular tree (clade IV, part). However, there are significant morphological differences exhibited by Cryptasterina : form pentagonal or subpentagonal; abactinal spinelets granuliform; at most very low raised rounded spinelet-bearing elevations along the proximal projecting edge of abactinal plates; superomarginal plates larger than inferomarginals; actinal spines fewer (oral 4–5, suboral 1–2, furrow 2–3, subambulacral 1–2 proximally, actinal interradial 0–1 proximally 1–2 distally), conical to digitiform; adradial actinal series of plates with none to few spines; superambulacral plates not in complete series, few present.
In an electrophoretic study of five Asterina species from Japan, Matsuoka (1981) found them to be a heterogeneous phylogenetic group with A. batheri and A. coronata japonica closely related, A. pseudoexigua pacifica fairly closely related, and A. pectinifera and A. minor each probably belonging to separate lineages. This result is generally congruent with the molecular phylogeny of Waters et al. (2004), except that in their clades A. minor was not separate but in the same lineage as A. batheri and A. coronata .
H.L. Clark (1938) described juvenile specimens of Paranepanthia rosea (up to R = 8 mm). Material examined in this work from Jurien Bay was significantly larger (up to R = 17 mm) and characterised by: absence of two distinct ‘fields’ of abactinal plates; splay-pointed sacciform spinelets and spines; predominantly large single papulae in papular spaces; numerous longitudinal series of papulae along the sides of rays; actinal plates not in oblique series; presence of series of superambulacral plates; and single superactinal plate supports. P. rosea is removed from Paranepanthia . The inferomarginal plates define the margin in P. rosea , not superomarginal plates as stated by H.L. Clark (1938).
Mortensen (1940) distinguished a variety iranica for the species A. cepheus on the basis of having 5–6 suboral spines per plate and A. cepheus two (up to four seen in this work). Further to this observation, the variety is distinguished from A. cepheus (characters in brackets) by having: up to eight stout sacciform spinelets in single series across proximal abactinal plates (up to 20 fine spinelets in bands across plates); up to ten stout inferomarginal spinelets per plate (more than, 20); typically two actinal spines per plate (4–8). Asterina cepheus iranica is raised to species status.
Marsh (1977) compared the three similar small fissiparous Indo-Pacific asterinids A. anomala , A corallicola and A. heteractis , and her confirmation of valid species status for all three is supported here. A. heteractis has some characters which are exceptional to those shared by other Aquilonastra species: regular doubly papulate carinal series for length of ray; actinal plates in oblique series; fewer actinal spines per plate, with up to four oral, sometimes no suboral, up to four furrow, up to three actinal. These differences are not sufficient to justify assignment to another genus.
Smith (1927), Liao (1980) and Liao and Clark (1995) observed that A. limboonkengi was close to A. cepheus (Smith as close to A. burtoni from the Philippines, A. burtoni presumably A. cepheus ). Liao and Clark (1995) described the differences between A. cepheus and A. limboonkengi . Our observations confirm that the abactinal spinelets in A. cepheus are long and fine, and those in A. limboonkengi short and stout. Rowe (1995) rejected the subspecies A. coronata fascicularis Fisher, 1918 . The possible species status of the other subspecies of A. coronata (cristata Fisher, 1916 and euerces Fisher, 1917) has not been examined.
Pairs of abactinal spinelets on A. anomala , A. cepheus , A. corallicola and A. mino r are sometimes angled and contiguous, suggesting that they may act as pedicellariae. But the spinelets are not differentiated in form.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |