
Gekko japonicus
|
publication ID |
https://doi.org/10.5281/zenodo.278393 |
|
DOI |
https://doi.org/10.5281/zenodo.6188386 |
|
persistent identifier |
https://treatment.plazi.org/id/19358A1A-8B07-FFB8-8980-FB73FEC5F7A7 |
|
treatment provided by |
Plazi |
|
scientific name |
Gekko japonicus |
| status |
|
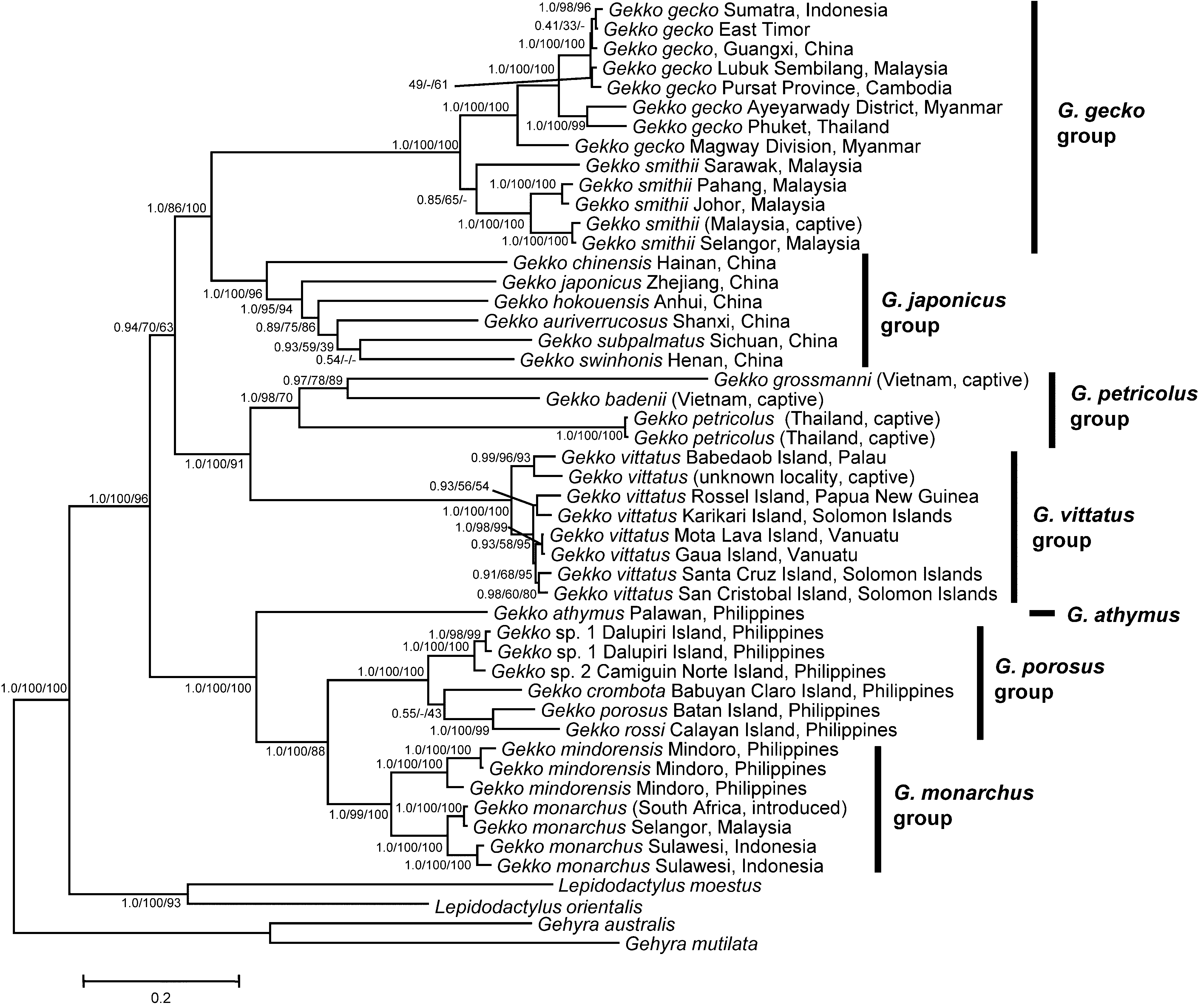
Gekko japonicus group
( Figures 4 View FIGURE 4. A , 5A View FIGURE 5. A )
Species. G. auriverrucosus Zhou & Liu 1982 ; G. canhi Rösler, Nguyen, Doan, Ho & Ziegler 2010 ; G. chinensis Gray, 1842 ; G. hokouensis Pope, 1928 ( Figure 4 View FIGURE 4. A B); G. japonicus ( Schlegel, 1836) ( Figure 4A View FIGURE 4. A ); G. l i b o e n s i s Zaho & Li, 1982; G. melli Vogt, 1922 ; G. palmatus Boulenger, 1907 ( Figure 4 View FIGURE 4. A C); G. scabridus Liu & Zhou, 1982 ; G. s c i e n - tiadventura Rösler, Ziegler, Vu, Herrmann & Böhme, 2004 ( Figure 4 View FIGURE 4. A D); G. shibatai Toda, Sengoku, Hikida & Ota, 2008 ; G. similignum Smith, 1923 ; G. subpalmatus Günther, 1864 ; G. swinhonis Günther, 1864 ; G. taibaiensis Song, 1985 ; G. t a w a e n s i s Okada, 1956; G. vertebralis Toda, Sengoku, Hikida & Ota, 2008 ( Figure 5A View FIGURE 5. A ); G. wenxianensis Zhou & Wang, 2008 ; G. yakuensis Matsui & Okada, 1968 .
Characters. 58.9–99.2 mm SVL; nares in contact with rostral (except for G. auriverrucosus ); nasals 3 (rarely 2 in G. c h i n e n s i s); postmentals relatively small (e.g., G. similignum ), largest in G. c a n h i, G. c h i n e n s i s, G. palmatus , G. scientiadventura ; 0–21 dorsal tubercle rows; 0–32 precloacal pores; postcloacal tubercles 1–4; webbing between fingers and toes weakly developed to extensive ( G. chinensis , G. m e l l i, G. palmatus , G. scientiadventura , G. similignum , G. subpalmatus ); tubercles present on fore and hind limbs, hind limbs only, or lacking all together; lateral fold without tubercles; subcaudals enlarged, in a longitudinal row (in G. yakuensis medially subdivided); head pattern present or not, without figure-shape (UU- to W-shaped in G. m e l l i and W-shaped in G. scientiadventura ); vertebral region with relatively large, light flecks, blotches or bands.
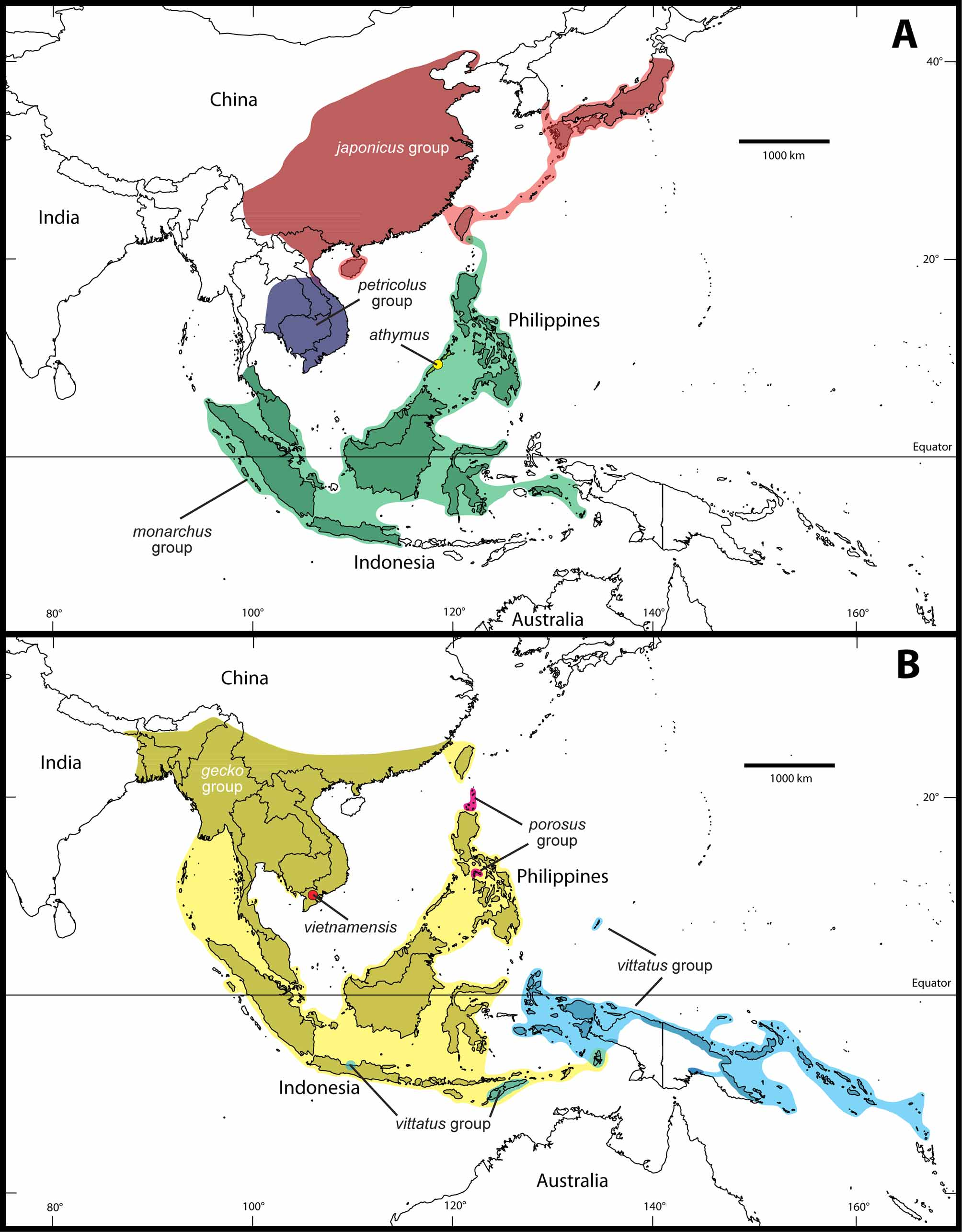
Distribution. China, Japan, Korea, Taiwan, and Vietnam including offshore islands; possibly Laos ( Figure 3 View FIGURE 3 A).
Remarks. The Gekko japonicus group is very complex. Different morphological features of the species of this group (e. g., nares in contact with rostral or not, broad webbing between fingers and toes or not, dorsal tubercles absent or present, tubercles on limbs absent or present, see Table 2) demonstrate that this genetically well-defined clade is highly morphologically variable (see Figure 1 View FIGURE 1 ).
Numerous authors have listed Gekko chinensis as a representative of Vietnam’s herpetofauna (e.g., Bourret 1937b, 1940a, 1943; Zhao & Adler 1993; Welch 1994; Bobrov 1995a; Rösler et al. 2005). According to Nguyen et al. (2005), G. chinensis is widely distributed in the north of the country, whereas G. palmatus is only recorded from the two northern provinces of Lang Son and Vinh Phuc (see also Nguyen & Ho 1996). However, according to Ziegler et al. (2006) and based on new locality records (see Appendix 1) the distribution of G. palmatus in Vietnam in fact extends from several northern provinces southward to central Vietnam in Quang Binh Province. Taking into account the records given for G. c h i n e n s i s by Nguyen & Ho (1996), the latter species must occur in sympatry with G. palmatus at least in the northern Vietnamese provinces of Bac Giang, Bac Kan, Cao Bang, Lang Son, Quang Ninh, and Vinh Phuc. However, all Gekko studied by us from these provinces proved to be G. palmatus and none had the diagnostic characters of G. c h i n e n s i s. This also makes the records of G. chinensis from Hai Duong and Ninh Binh provinces (see Nguyen & Ho 1996) questionable and they are probably based on misidentifications of G. palmatus . Bobrov & Semenov (2008) listed all previous identifications of G. chinensis from Vietnam as G. palmatus . Ota et al. (1995) believed G. c h i n e n s i s and G. palmatus to represent allopatric taxa (see also Matsui & Ota 1995). Concordant with the latter authors we herein suggest that the known occurrence of G. chinensis is limited to China (southern provinces of Fujian, Guangdong, Guangxi, Guizhou, Hong Kong, Sichuan, and Yunnan) whereas that of G. palmatus is only in northern and central Vietnam (Bac Giang, Bac Kan, Cao Bang, Hai Duong, Hai Phong, Lang Son, Ninh Binh, Quang Binh, Quang Ninh, and Vinh Phuc provinces). Further collections or study of museum specimens are needed to clarify the identification of populations along the China-Vietnam border and whether G. chinensis and G. palmatus occur in sympatry.
The distinguishing character of broad toe webbing in G. palmatus versus a narrower webbing in G. c h i n e n s i s, which can regularly be found in the older literature (e.g., Boulenger 1907; Smith 1935; Bourret 1937b), may have conspicuously added to the above listed confusion and misidentifications, because it represents an ambiguous diagnostic character (Ota et al. 1995). Smith (1935: 114) even concluded that G. palmatus is “possibly a racial form of G. c h i n e n s i s.” However, G. palmatus and G. chinensis can be easily distinguished by the size of the internasals compared to the nasorostrals (internasals same size or larger than nasorostrals in G. chinensis versus always smaller in G. palmatus ) and statistically significant differences in the number of midbody scales and the number of lamellae on the first, third and fifth fingers as well as the first toe (Ota et al. 1995). A key character that allows for proper identification of both aforementioned species is the presence of limb tubercles. These are present in G. c h i n e n s i s, but are completely lacking in G. palmatus (our data). Ota et al. (1995) indicated shank tubercles for G. chinensis and the specimen depicted in Zhao & Adler (1993: pl. 21, fig. G.) has tubercles both on the fore and hind limbs; but the specimen of G. palmatus depicted by Nguyen et al. (2009: fig. 314) completely lacks such tubercles.
The taxonomic history of G. similignum is controversial. The type locality of this taxon is Hainan Island ( Smith 1923). Stejneger (1932) synonymized G. similignum with G. chinensis (see also Zhao et al. 1999). However, Ota et al. (1995) and Kluge (2001) listed it as valid at the specific rank. Gekko chinensis and G. smilignum differ in maximum SVL, in the number of scales around midbody and in the number of lamellae on the first and fourth toes (see Table 1).
Gekko liboensis was described by Zhou & Li (1982) based on an adult female specimen collected in 1979. Three further specimens (two females and a juvenile) were collected in 1984 ( Zhao et al. 1999). Zhao & Adler (1993) synonymized G. liboensis with G. hokouensis (see also Günther 1994; Kluge 2001; Henkel & Schmidt 2003). In contrast, Kluge (1993), Bauer (1994), Welch (1994), Matsui & Ota (1995), Zhao et al. (1999), Rösler (2000), Rösler et al. (2005, 2006) treated G. liboensis as a valid species. Gekko liboensis differs from G. hokouensis by a larger SVL (85 versus 70 mm), absence of internasals, more interorbitals (40 versus 30–33), flat dorsal tubercles, fewer dorsal tubercle rows (10 versus 12–18) and by the number of lamellae below the fourth toe (9 versus 15–18). However, as adult males are still lacking for G. l i b o e n s i s, important diagnostic characters and possibly additional distinguishing features such as the number of precloacal pores are still missing. The nares of G. auriverrucosus are not in contact with the first supralabial (Zhou & Liu 1982), which seems to represent a derived character of the Gekko gecko group (except for G. verreauxi ).
Zhao & Adler (1993) synonymized G. scabridus with G. chinensis (see also Kluge 2001), but Matsui & Ota (1995) and Zhao et al. (1999) again evaluated the former taxon as valid species. Liu & Zhou (1982) pointed to the phenotypic similarity between G. scabridus and G. japonicus . According to Liu & Zhou (1982), G. scabridus differs from G. japonicus by having tubercles on its back and hind limbs and by a higher count of precloacal pores. From Table 1 it is obvious that there exist additional distinguishing features between the two species. Compared with G.
japonicus , G. scabridus has fewer interorbitals (30 versus 32–35), fewer lamellae below the first toe (6–9 versus 10–12), and fourth toe (7–9 versus 14–16), more dorsal tubercle rows (17–21 versus 9–14) and a higher number of precloacal pores (10–15 versus 6–9). In comparsion to G. chinensis it has only slightly developed webbing between the fingers and toes, fewer precloacal pores, and more postcloacal tubercles. We, therefore, accept the specific status of G. scabridus and regard it as a valid member of the Gekko japonicus group.
Vogt (1922) originally described G. m e l l i based on a large series of which now only two specimens still exist ( Bauer & Günther 1991). Stejneger (1932) listed G. m e l l i as a subspecies of G. subpalmatus and Pope (1935) finally synonymized the former species with G. subpalmatus . Rösler et al. (2005) revalidated G. melli as a distinct species (see also Rösler & Tiedemann 2007).
With respect to G. subpalmatus it is interesting to note that the back of the holotype is slightly banded and that the vertebral region shows four indistinct light blotches (Rösler et al. 2005: figs. 11–12). However, the ground coloration of the live specimen depicted in Zhao & Adler (1993: their Pl. 32, fig. B) is blackish-olive (Black 3U). This specimen, in addition, has a repeatedly interrupted orange (722U) vertebral stripe and tail blotches of the same color; a banded pattern as observable from the holotype is not discernible (see also fig. in Huang & Zong 1998: pl. 29). Whether this reflects intraspecific variation or a persistent confusion over the identity of this species remains to be investigated.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.