
Pleuroxus carolinae ( Methuen, 1910 )
|
publication ID |
https://doi.org/ 10.1080/00222930600958870 |
|
persistent identifier |
https://treatment.plazi.org/id/1A2F87C2-0A01-FF8D-0499-FE188EB0FD65 |
|
treatment provided by |
Carolina |
|
scientific name |
Pleuroxus carolinae ( Methuen, 1910 ) |
| status |
|
Pleuroxus carolinae ( Methuen, 1910) View in CoL
( Figures 24–48 View Figures 24–32 View Figures 33–38 View Figures 39–46 View Figures 47–55 )
Pleuroxus inermis Sars, 1896 View in CoL in Sars 1916, p 342 –343, Plate 41, Figure 2, 2a, b View Figures 1–11 .
Chydorus carolinae Methuen 1910, p 157 , Plate 16, Figure 44a, b View Figures 39–46 ; Methuen 1911, p 255.
Neotype locality
Small rockpools (28 u 409430S, 28 u 509280E) at 2050 m a.s.l. near Fika Patso, Qwaqwa, Freestate, Republic of South Africa. The sample was collected on 8 November 1996 by K. Martens, and marked as RSA /96/111.
The former type locality was ‘‘Lake Chrissie, the lake lies in the Carolina district due east of Pretoria near the borders of Swaziland’’ ( Methuen 1910).
Material examined
Neotype: a parthenogenetic female in 96% alcohol, MGU Ml 56. Author’s type material is apparently lost.
Other material examined. Five pathenogenetic females from neotype locality (old number RSA/96/111); four females from Sani River pool 5 at 2890 m a.s.l. (29 u 349320S, 29 u 179560E), Sani Plateau, Lesotho, collected on 23 March 1995 by K. Martens, NNS 2002-073 (old number LES/95 About LES /026) .
Short diagnosis
Body dark brown, not transparent, in anterior view dorsum as a smoothed angle, but true medial keel absent, postero-ventral angle without denticles. Rostrum does not reach apex of labral keel. Head shield posteriorly widely rounded, anteriorly makes a blunt rostrum, PP51.5–2 IP. Obscure striation expressed only in antero-ventral and postero-ventral portions of valve. All setae of valve ventral margin exactly marginal. Setules on posterior margin of valve situated on inner side of valve, relatively far from the margin, so their tips do not reach it. Postabdomen relatively long, slightly narrowing distally, with rounded and slightly prominent dorso-distal angle, its anal margin as long as preanal margin and clearly shorter than postanal margin, anal teeth thin and rather long, organized in successive series of two to three, rarely solitary, slightly increasing in size distally. Anntenna I not reaching tip of rostrum, with a strong basal peg. On antenna II, all apical setae subequal in size. On limb I smallest ODL seta rudimentary, two largest IDL setae unequal in size. On innerdistal portion of limb II, scrapers 1–3 with size decreasing basally, scraper 8 smaller than others; setules on gnathobase naked, filter plate II with nine, rarely 10 setae. On exopodite III seta 1 shorter than 3, filter plate III with eight setae. On inner portion of limb IV setae 2– 4 with robust setules, filter plate IV with six to seven setae. Exopodite V with two projections distally to distal seta 1.
Redescription
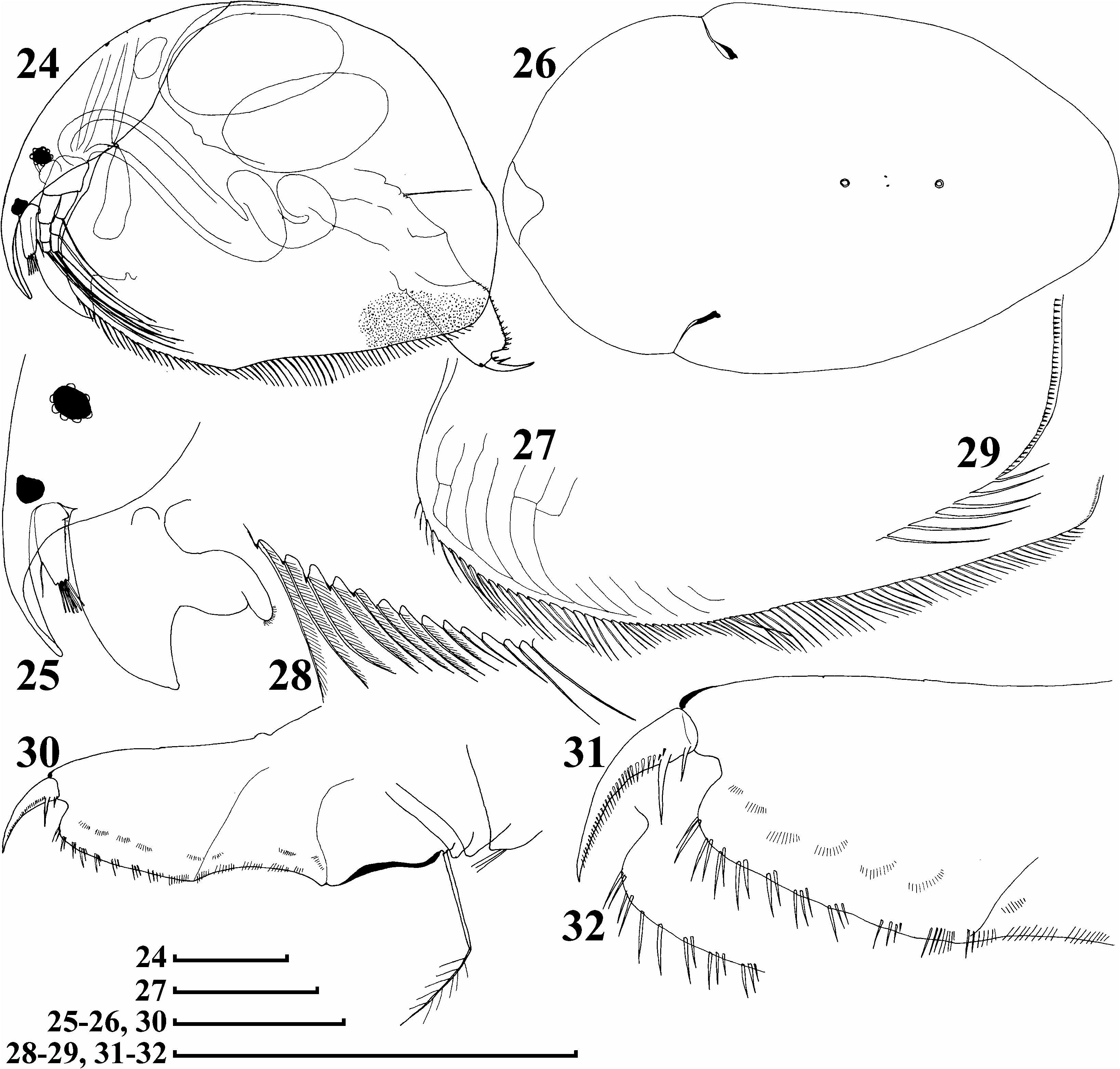
Parthenogenetic female. Body brown, not transparent. In lateral view body widely oval, high (body height/body length50.74– 0.80 in adults), maximum height in middle ( Figures 24 View Figures 24–32 , 39, 40 View Figures 39–46 ). Dorsal margin evenly arched from tip of rostrum to postero-dorsal angle, which is well-defined, posterior margin straight, postero-ventral angle broadly rounded, without teeth, ventral margin with a slight prominence in middle. Obscure striation expressed only in antero-ventral ( Figure 27 View Figures 24–32 ) and postero-ventral portions of valve, while entire valve punctuate (in Figure 24 View Figures 24–32 this punctulation is represented only in postero-dorsal region). In anterior view body subovoid, with dorsum a smoothed angle, but dorsal keel absent.
Head with long rostrum, protruding downward and posteriorly ( Figures 25 View Figures 24–32 , 41–43 View Figures 39–46 ). Eye only slightly smaller than ocellus, distance from tip of rostrum to ocellus greater than that between ocellus and eye. Head shield elongated, with maximum width immediately behind mandibular articulation, its posteriormost extremity widely rounded ( Figure 26 View Figures 24–32 ). Two major head pores, PP51.5–2 IP. Lateral head pores minute, normally located asymmetrically to midline. Labrum with fleshy main body, small distal labral plate (anterior plug in terms of Dumont and Silva-Briano 2000), and large medial labral keel, with well-defined apex projected significantly behind rostrum.
Valves large, ventral margin armed with numerous setae of different size in different regions, all plumose and located exactly marginally ( Figures 27–29 View Figures 24–32 ). A row of small setules situated on inner side of posterior valve margin, relatively far from the margin, so their tips do not reach it ( Figure 29 View Figures 24–32 ).
Postabdomen elongated, wide, its ventral margin almost straight ( Figures 30 View Figures 24–32 , 45 View Figures 39–46 ). Preanal margin slightly concave ( Figure 44 View Figures 39–46 ), as long as anal margin, preanal and postanal angle well-defined, postanal margin clearly longer than anal margin, dorso-distal angle widely rounded and slightly prominent distally, inflated basis of claws bordered from postanal margin by a distinct depression ( Figures 31, 32 View Figures 24–32 , 46 View Figures 39–46 ). Each side of postanal portion provided with a row of thin and rather long postanal teeth, organized in successive series of two to three, rarely solitary, slightly increasing in size distally. Series of postanal denticles evenly grading into five to six series of marginal setules on anal margin. Laterally to marginal denticles, a row of fascicles consisting of short, fine setules. Postabdominal seta relatively short, with basal segment approximately as long as preanal margin and distal segment as long as basal one, supplied with delicate setules. Postabdominal claw short (significantly shorter than preanal or anal margin), massive, evenly curved, with setules along ventral margin, and two basal spines, proximal one being half size of distal ( Figures 46 View Figures 39–46 , 47 View Figures 47–55 ), sometimes basalmost spine duplicated ( Figure 48 View Figures 47–55 ).
Antenna I not reaching tip of rostrum, slightly narrowing distally, with a well-defined basal peg ( Figure 33 View Figures 33–38 ). Antennular sensory seta slender, longer than half the antennule, arising at one-third of antennular length from distal directly from antenna I, without any prominences. Nine short aesthetascs of slightly differing size. Antenna II relatively short, coxal part with two sensory setae, basal segment robust, with a rudimentary distal spine ( Figure 34 View Figures 33–38 , arrow). Antennal branches relatively elongated, exopod and endopod subequal in length, all segments cylindrical, antennal formula, setae 0-0-3/1-1-3, spines 1-0-1/0-0-1. Exopod and endopod apical swimming setae subequal in size. No chitinous insertions within distal segments. Basal lateral seta thinner and shorter than distal lateral seta. Spine on basal segment of exopod rudimentary.
Limbs very similar to those in P. wittsteini , but limb I with ODL smaller seta rudimentary, IDL second and third setae unequal in size ( Figure 35 View Figures 33–38 ), filter plate II ( Figure 36 View Figures 33–38 ) with nine, and rarely 10, setae; scrapers 1–3 on limb II with size decreasing basally, scraper 8 smaller than others; seta 1 on exopodite III shorter than seta 3 ( Figure 37 View Figures 33–38 ); filter plate IV with six to seven setae; seta 1 on exopodite V especially long, longer than seta 2 on inner limb portion and armed with rare setules ( Figure 38 View Figures 33–38 ).
Ephippial female, male
Unknown.
Size
Parthenogenetic females 0.42–0.47 mm in available material.
Distribution
Known from four high-altitude localities, our two localities plus those of Methuen (1910) and Sars (1916), on the Sani Plateau and the Cape Flats.
Ecology
Our specimens were collected from pools, at pH 7.4–7.9, conductivity 142 mS cm 21, 14.8– 19.6 u C, altitude 2050–2890 m a.s.l. The species occurs also in Lake Chrissie, a large and shallow high-altitude lake.
Comments
The taxon was reported by Sars (1916) as P. inermis from a pond in the Cape Flats; he said that his ‘‘ P. inermis ’’ was ‘‘without any obvious denticles’’ at postero-ventral angle, had reticulation only at anterior and posterior margins, and strong postanal spines. Then, Methuen (1910) established Chydorus carolinae as a new species. His too small and inadequate description was accompanied with quite realistic illustrations ( Methuen 1910, Plate 16: Figure 44a, b View Figures 39–46 ). Keeping in mind that the type material was apparently lost, and that aduncus -like species of Pleuroxus from the southern hemisphere posed a difficult taxonomic problem, we selected the neotype of this species ‘‘to define the nominal taxon objectively’’ ( International Commission on Zoological Nomenclature 2000, Article 75).
There are also other aduncus -like forms in Africa ( Rühe 1914; Jenkin 1934; Chiambeng and Dumont 2004), but their status must be checked accurately.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Genus |
Pleuroxus carolinae ( Methuen, 1910 )
| Smirnov, Nikolai N., Kotov, Alexey A. & Coronel, Jorge S. 2006 |
Pleuroxus inermis
| Sars GO 1916: 342 |
Chydorus carolinae
| Methuen PA 1911: 255 |
| Methuen PA 1910: 157 |