
Pazliopsis reyi, Friis & Crane & Pedersen, 2018
|
publication ID |
https://doi.org/10.2478/if-2018-0010 |
|
persistent identifier |
https://treatment.plazi.org/id/1A4787F5-FFB9-E115-FC2A-65DEFD37B979 |
|
treatment provided by |
Diego |
|
scientific name |
Pazliopsis reyi |
| status |
gen. et sp. nov. |
Pazliopsis reyi gen. et sp. nov.
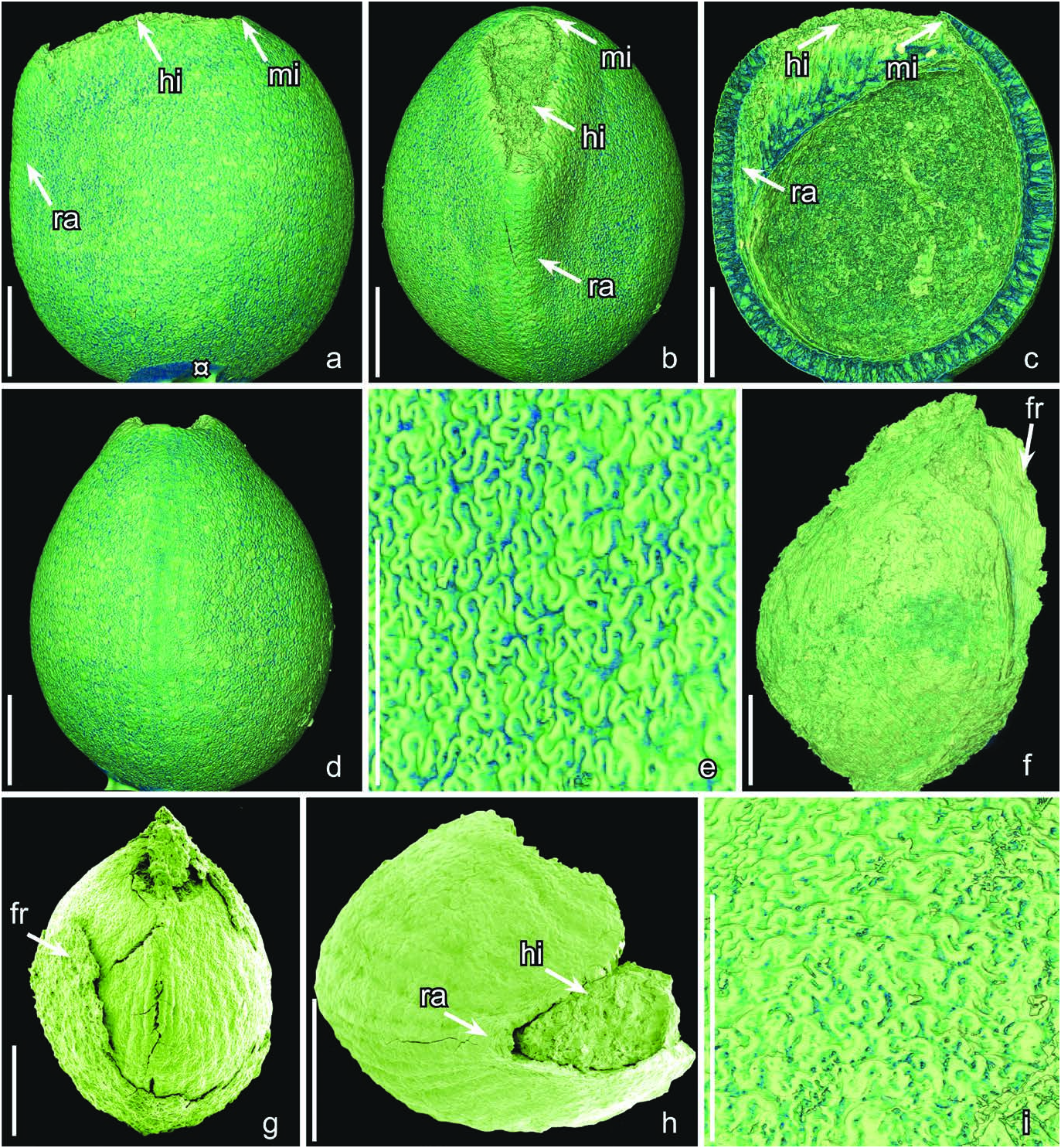
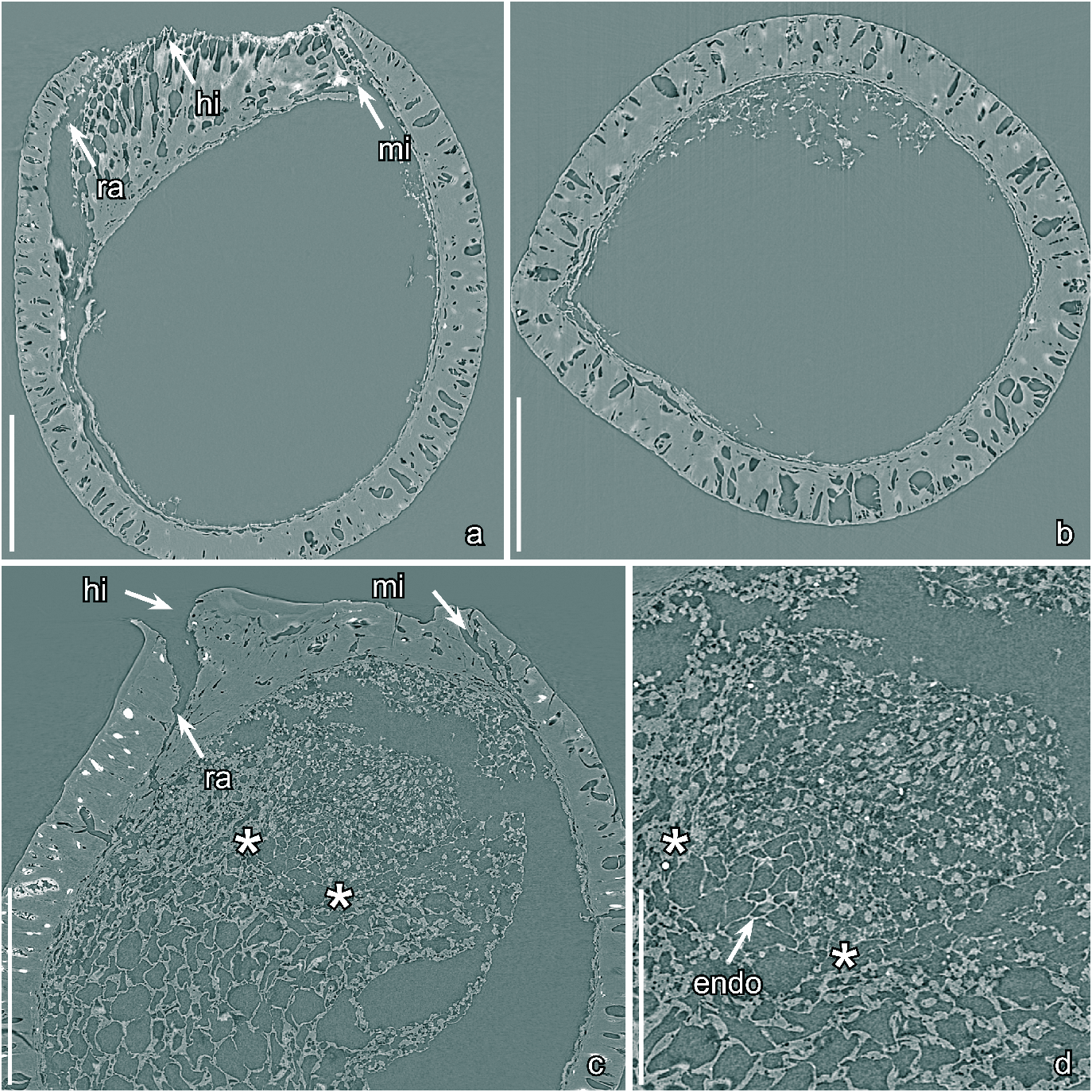
Text-figs 3f–i View Text-fig , 4c, d View Text-fig
H o l o t y p e. Designated here, S171534 (Torres Vedras sample 043; illustrated here on Text-fig. 3g View Text-fig ).
P l a n t F o s s i l N a m e s R e g i s t r y N u m b e r.
PFN000093 (for new species).
P a r a t y p e s. Designated here, S174735 (Torres Vedras sample 043), S174636, S174637, S175024 (Torres Vedras sample 044), S175120 (Torres Vedras sample 144), S174632 (Torres Vedras sample 298).
R e p o s i t o r y. Palaeobotanical Collections , Department of Palaeobiology, the Swedish Museum of Natural History, Stockholm, Sweden .
E t y m o l o g y. In recognition of Jacques Rey (*1940, †2018) for his important contributions to understanding the Cretaceous geology of Portugal.
T y p e l o c a l i t y. Torres Vedras, Portugal (NE of
Forte de Forca; 39°06′13″N, 9°14′47″W).
T y p e s t r a t u m a n d a g e. Lower part of the Almargem Formation; Early Cretaceous (late Barremian – early Aptian).
D i a g n o s i s. As for the genus.
D i m e n s i o n s. Length of fruit: 1.1 mm; length of seeds: 0.7–0.9 mm; width of seeds: 0.66–0.84 mm.
D e s c r i p t i o n a n d r e m a r k s. The species is based on two fruits and 30 isolated seeds. Four specimens (S171534, S174614, S174632, S174633) were studied using SRXTM. The fruits or fruitlets are small and each contains a single seed ( Text-figs 3f, g View Text-fig , 4c, d View Text-fig ). They are isolated from the floral receptacle and it is unknown whether they were from monocarpellate gynoecia or multicarpellate gynoecia that were apocarpous. The fruit wall is thin, collapsed and partly abraded exposing the seed. It consists of an outer zone of small thin-walled cells and an inner zone of obliquely arranged, longitudinally elongate, fibers ( Text-fig. 3f View Text-fig ). The fibrous inner zone suggests that the fruit was dry.
The seeds are small, anatropous, bitegmic and exotestal with bilateral symmetry, ovate to almost circular in lateral view with a truncate profile at the hilar-micropylar end of the seed ( Text-fig. 3f, g View Text-fig ). The hilar scar is large and ovate in outline and lacks a hilar rim ( Text-fig. 3h View Text-fig ). Where the raphal bundle enters the seed the micropyle and hilum are separated by expanded sclerenchyma tissue beneath the hilar scar ( Text-fig. 4c View Text-fig ), and marked on the seed surface by a transverse slit through the outer integument (testa) adjacent to the hilar scar.
The seed coat is mainly comprised of the exotesta, which consists of a single layer of tall, columnar sclerenchyma cells. These cells are about 100 µm tall over most of the seed but gradually become shorter towards the hilarmicropylar region ( Text-fig. 4c View Text-fig ). The anticlinal walls of the exotestal cells are thickened, and of almost even thickness from the outside to the inside resulting an almost straight lumen ( Text-fig. 4c View Text-fig ). The anticlinal walls of the exotestal cells are strongly undulate towards the outside and inside forming a jigsaw puzzle-like pattern on the seed surface with rounded, deep, undivided lobes ( Text-fig. 3i View Text-fig ). The cell walls are also undulate over the raphe and in the micropylar region. The inner layers of the testa (mesotesta/endotesta) and the tegmen, which is poorly preserved in the micropylar region, are composed of thin walled cells that are typically collapsed.
One specimen has well-preserved nutritive tissue and an embryo ( Text-fig. 4c, d View Text-fig ), and another specimen has a remains of the embryo preserved. In both cases the embryo is tiny, with two rudimentary cotyledons ( Text-fig. 4c, d View Text-fig ). The embryo to seed (E:S) ratio is about 0.02. As in Gastonispermum portugallicum , the cells of the embryo are much smaller than those of surrounding nutritive tissue ( Text-fig. 4c, d View Text-fig ), and each contains a central structure that we interpret as the probable remains of a nucleus ( Text-fig. 4d View Text-fig ). The nutritive tissue immediately surrounding the embryo consists of equiaxial cells with thin-walled, straight to slightly wavy cells walls ( Text-fig. 4d View Text-fig ). This tissue is clearly endosperm. However, the transition between these inner thin-walled endosperm cells and the bulk of the surrounding nutritive tissue is not well preserved so it is uncertain whether all the nutritive tissue is endosperm or whether the outer part of the nutritive tissue could be perisperm.
| R |
Departamento de Geologia, Universidad de Chile |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.