
Ranitomeya vanzolinii Myers 1982
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3083.1.1 |
|
persistent identifier |
https://treatment.plazi.org/id/1D338788-950A-1516-C8FC-9B5C3960FE30 |
|
treatment provided by |
Felipe |
|
scientific name |
Ranitomeya vanzolinii Myers 1982 |
| status |
|
Ranitomeya vanzolinii Myers 1982 View in CoL
Account authors: J.P. Caldwell, J.L. Brown, E. Twomey, P.R. Melo-Sampaio, M.B. Souza
Figs. 3 View FIGURE 3 , 4 View FIGURE 4 , 9 View FIGURE 9 , 23 View FIGURE 23 (j, k), 28, 31
Tables 1, 4 – 6
Dendrobates quinquevittatus (non Steindachner, 1864)—Silverstone 1975 (partim): p. 33, Fig. 12 View FIGURE 12 (drawing), pattern D and 14 (drawing), pattern E
Dendrobates vanzolinii Myers 1982: p. 9 View in CoL , Fig. 4 View FIGURE 4 , Table 1 [MZUSP 51597 collected by Paulo E. Vanzolini and Miguel Petrere at Pórto Walter on Rio Juruá, Acre, Brazil];— Vanzolini 1986; Caldwell & Myers 1990: p. 4; Caldwell 1997: p. 211, Fig. 1; Moreira et al. 1997; Caldwell & de Oliveira 1999: p. 565, Table 1 – 2, Figs. 1 – 3; Schulte 1999: p. 110, Fig. DB-032, DB-38; Symula et al. 2003: p. 452, Table 1, Figs. 1, 3 – 6 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 ; Christmann 2004: p. 6, Figs. on p. 36. 158; Pröhl 2005: p. 358, Table 1; Roberts et al. 2006a: p. 379, Table 1, Figs. 1, 4 View FIGURE 4 ; Santos et al. 2009, by implication
Ranitomeya vanzolinii View in CoL — Bauer 1988: p. 1; Grant et al. 2006: p. 171, Fig. 76; Lötters et al. 2007: p. 497, Figs. 498 – 629; von May et al. 2008a: p. 396, Appendix 2; Souza 2009: p. 31.
Background information. A population of R. vanzolinii View in CoL from near Porto Walter, State of Acre, Brazil was the focus of intensive behavioral study ( Caldwell 1997; Caldwell & de Oliveira 1999) and included in several phylogenetic studies ( Symula et al. 2003; Grant et al. 2006). Little information has been published on other populations of this species. Some confusion exists regarding the taxonomic status of R. flavovittata View in CoL , the sister species to R. vanzolinii View in CoL (see R. flavovittata View in CoL account for more details).
Tadpole. A total of seven tadpoles of Ranitomeya vanzolinii were collected from Porto Walter , Acre, Brazil, from 4–17 April 1996. A tadpole in Stage 38 ( OMNH 36057 View Materials ) was chosen for a complete description. All seven tadpoles were used to examine variation. These tadpoles ranged from stages 26 to 40, although not all stages were represented in the sample .
Description of OMNH 36057. Total length 27.8; body length 8.9; tail length 18.9, tail length 67.9% total length. Body depressed, maximum width 6.1, maximum depth 4.4. Snout broadly rounded in dorsal view, rounded in lateral view. Nares small, directed anterolaterally, 0.9 from tip of snout; internarial distance 1.6, interorbital distance 1.5. Distance from naris to anterior edge of eye 1.0, eye width, 1.4. Eyes dorsal, directed dorsolaterally.
Spiracle sinistral, short, 1.0, forming small tube free only on outer margin, opening directed posterodorsally 5.7 from tip of snout. Spiracle located 63.8% of body length from tip of snout, just below horizontal midline of body. Vent tube dextral, 0.5 in length.
Caudal musculature depth at body–tail junction 3.0; depth of musculature nearly uniform until it begins tapering at posterior one-third of tail. Upper fin originates 1.1 behind tail–body junction, upper fin at midtail 1.0, slightly deeper than lower fin at midtail, 0.8. Tail tip rounded, notochord ends 0.8 from end tail.
Oral disc emarginate, located ventrally, not visible from above. Transverse width oral disc 2.4, 38.9% of body width. Lower labium free from body wall. Short, conical papillae present on entire margin of posterior labium and laterally on anterior labium; most of anterior labium free of papillae. Papillae mostly in single row, double in few places. Lower jaw sheath broadly V-shaped, deeper than upper jaw sheath. Transverse width upper jaw sheath 1.1, 44.5% transverse width oral disc. Both upper and lower jaw sheaths with finely serrated cutting edges, serrations becoming smaller on proximal lateral processes of upper sheath, absent on ends of lateral processes.
Labial tooth row formula 2(2)/3(1). Labial tooth row A-1 complete, 1.5; A-2 with medial gap, 0.9, right half A- 2 length, 0.3, 20.0% of A-1. P-1 with medial gap, 0.1; right half P-1 length, 0.5, 44.6% of P-2; P-2 and P-3 equal in length, 1.1.
Variation. The two smallest tadpoles, stages 26 and 27, averaged 15.1 ± 1.2 total length. Four tadpoles in stages 34 to 38 averaged 24.8 ± 1.2 total length (range, 22.1–27.8). The single tadpole in early metamorphosis at stage 40 was 24.7; in this specimen tooth rows were becoming irregular, the spiracle opening is beginning to close, and the vent tube is partially absorbed.
Tail length as a percentage of total length averaged 62.8 ± 0.01 (range, 60.7–67.9). In the two smallest tadpoles, the upper fin originated at the tail–body junction, whereas in the five largest tadpoles, the fin was low at the tail–body junction, but began to increase in height at an average of 1.4 ± 0.2 (range, 1.0–1.8) behind the tail–body junction. Height of the upper and lower fins was nearly equal in the two smaller tadpoles (upper, 0.40 and 0.45; lower, 0.70 in both individuals). Height of the upper fin was slightly greater in the five larger tadpoles (upper, 1.12 ± 0.10, range 0.90–1.46; lower, 1.01 ± 0.10, range, 0.80–1.38). Tail musculature depth at the tail–body junction was 2.5 ± 0.2 (range, 2.1–2.9) in the five largest tadpoles.
Transverse width as a percentage of body width in the seven tadpoles averaged 39.4 ± 0.01 (range, 35.7–45.6). Transverse width of the upper jaw sheath as a percentage of the transverse width of the oral disc averaged 49.7 ± 0.01 (range, 44.5–55.6). A-2 gap is larger than the P-1 gap. Average percentage of the width of the A-2 gap compared to the total width of A-2 is 49.4 ± 0.04 (range, 37.1–60.5). Average percentage of the width of the P-1 gap compared to the total width of P-1 is 17.8 ± 0.04 (range, 8.3–33.6). P-2 and P-3 are nearly identical in length in 5 tadpoles in which both were normal and well-developed. In the smallest tadpole, P-3 was shorter than P-2 but may not have been fully developed. P-3 was deformed in one other tadpole; the ends of P-3 curved upward and fused with P-2.
Color in life (based on tadpole observed in a small treehole, later transferred to open-air laboratory). Body dark gray, tail musculature lighter gray, fins opaque light gray. Eye black. First observed 3 March, tadpole nearing metamorphosis by 21 March and had developed small light yellow spots on dorsum.
Color in preservative. Dorsum of body grayish brown. Venter opaque, intestinal coils dark grayish brown, some organs seen through venter light yellow. Tail musculature light yellow-brown. Back legs grayish brown. Fins translucent. Eyes black.
Natural history. Ranitomeya vanzolinii exhibits biparental care and likely, as observed in R. imitator , a monogamous mating system ( Caldwell & de Oliveira 1999). After one to two embryos hatch, the male carries the tadpoles on its back and deposits each individually in a phytotelm. Paired males and females remain together in small territories. Males call sporadically each day, and, on average, the female mate responds to his calls every 4.8 days. The male guides the female to their tadpole, they undergo some courtship behavior, and the female deposits two (usually) unfertilized trophic eggs, which the tadpole immediately consumes. The tadpoles are continually fed until metamorphosis. For a more detailed discussion of these behaviors and their evolution see Caldwell (1997) and Caldwell & de Oliveira (1999).
In the Reserva Extrativista do Alto Juruá (REAJ) and Parque Nacional da Serra do Divisor (PNSD) of Brazil, this species is frequently observed in primary forests during the rainy season, mainly in areas abundant in bamboo ( Guadua sp. ), bromeliads, and other phytotelmata (i.e. tree holes; Souza 2009). Adults frequently climb and jump on leaves, stems, and trunks of herbaceous vegetation up to 4 meters in height. In REAJ and PNSD this species uses bamboo internodes, the leaf axils of bromeliads, and other phytotelmata for tadpole deposition ( Souza 2009). In forests near Porto Walter, this species uses primarily cavities in small saplings and woody vines for tadpole deposition ( Caldwell & de Oliveira 1999).
Vocalizations. Like other members of this species group, the call of R. vanzolinii is a loud trill ( Fig. 28 View FIGURE 28 ). Vocalization of Ranitomeya vanzolinii was studied at Porto Walter, Acre, Brazil, using two methods: recordings of males analyzed with Raven 1.3 (Cornell Laboratory of Ornithology, Ithaca, New York), and timing of vocalizations recorded during focal observations of males engaged in distinct behaviors. Focal observations were also made of vocalizing frogs defending territories.
Vocalizations of 9 individuals of R. vanzolinii were recorded with a Sony WMD-6 tape recorder. Temperatures at the time of recording ranged from 24.0–27.2 C. The number of calls analyzed from these recordings per individual varied from 2 to 10 (mean 6.8 calls/individual); calls were averaged within each individual, then across individuals to obtain means for each call variable. The call of R. vanzolinii is a short trill audible from> 5m. Mean number of calls per minute and call duration varied depending on the individual’s behavior (see below; Caldwell & de Oliveira, 1999). Mean number of pulses per call was 16.8 ± 1.7 (range, 8.6–23.8), and number of pulses per second was 31.3 ± 0.8 (range, 27.9–33.6). Dominant frequency ranged from 5162–5412 Hz (n = 7). Some calls revealed four harmonics; the lowest ranged from 2444–2745 Hz (n = 5).
During focal observations of males engaging in three behaviors, time of vocalization (to the second) was recorded using a stopwatch. The number of calls per minute as a measure of call rate was obtained during the following behaviors: 1) calling when no female was present; 2) calling when guiding a female to a tadpole rearing chamber; and 3) calling when inside a tadpole rearing chamber with a female. At times when calling from inside a tadpole rearing chamber (i.e., tiny vinehole or treehole), the calls given were much shorter and had only a few pulses, giving the call a drawn-out “creeking” sound. When calling alone, males vocalized at a rate of 3.41 ± 0.25 calls per minute (range, 0–11; n = 6). When calling while guiding a female, males called at a rate of 8.05 ± 0.32 (range, 0–17; n = 8), and when calling while inside a rearing chamber with a female, males vocalized at a rate of 3.13 ± 0.23 (range, 0–16; n = 7). Thus, males called more intensely when guiding a female to a rearing chamber than when calling alone (Mann-Whitney U-test, p <0.001), or when they were undergoing courtship behavior with a female inside a cavity (Mann-Whitney U-test, p <0.001). Call rate did not differ when males were alone compared to being in a rearing chamber with a female (Mann-Whitney U-test, p = 0.11). Call duration also varied depending on the male’s behavior. When alone, call duration averaged 0.721 ± 0.08 sec (range, 0.642 –0.799; n = 2), when guiding a female 0.378 ± 0.12 (range, 0.256 –0.500; n = 2), and when in a tadpole rearing chamber with a female 0.566 ± 0.09 (range, 0.291 –0.844; n = 5). Thus, when guiding a female, shorter calls were given at a faster rate.
Territories were maintained using vocal bouts in which two resident males in adjacent territories established posts about 2 m apart and alternated calls ( Caldwell and de Oliveira, 1999). In one such encounter in which the timing of vocalizations was recorded for 5 minutes, the bout (defined as number of sequential call given by a male before the other male called was 1.23 ± 0.63 calls (range, 1–5 calls). Calls were alternated between the two frogs 74 times during the 5-minute observation period.
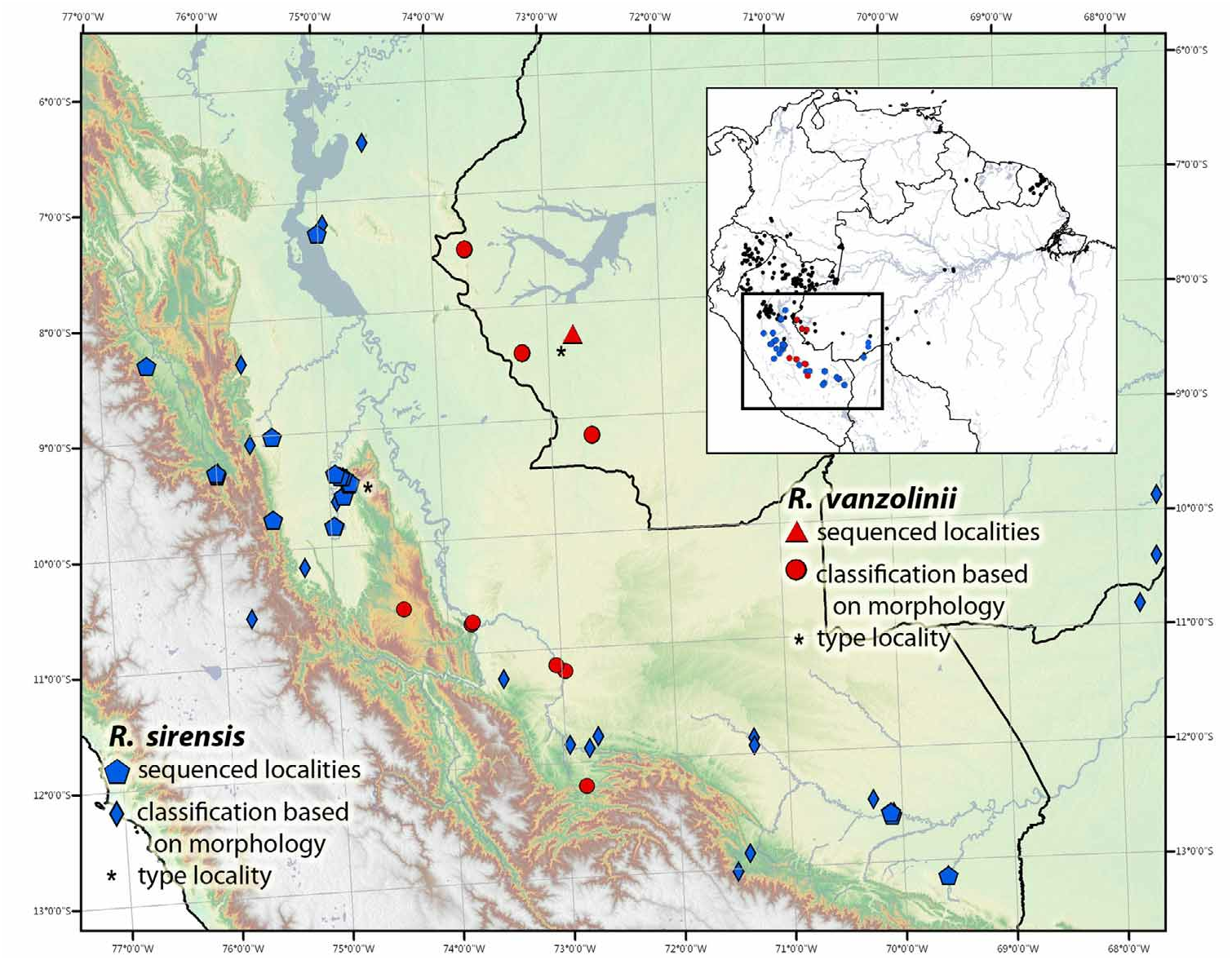
Distribution. Ranitomeya vanzolinii is a widespread species occurring from premontane cloud forest (approximately 1300 m elevation) in southern Peru, eastward to lowland rainforests in Brazil. This species is frequently observed on the east versant of Cordillera El Sira (where it has been subjected to frequent collection from smugglers for the pet trade) and in western Brazil in the vicinity of Porto Walter, Acre state. The distribution of this species appears to overlap with other members of the vanzolinii group, i.e., R. sirensis in the south of Peru and possibly R. cyanovittata in far eastern central Peru. However to date, there are no localities where two species within the vanzolinii group have been registered. This species occurs in Amazonian rainforests of Brazil (States: Acre and possibly Amazonas) and Peru (Departments: Cusco, Pasco, Ucayali), Fig. 31 View FIGURE 31 .








No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Ranitomeya vanzolinii Myers 1982
| Brown, Jason L., Twomey, Evan, Amézquita, Adolfo, Souza, Moisés Barbosa De, Caldwell, Jana- Lee P., Lötters, Stefan, May, Rudolf Von, Melo-Sampaio, Paulo Roberto, Mejía-Vargas, Daniel, Perez-Peña, Pedro, Pepper, Mark, Poelman, Erik H., Sanchez-Rodriguez, Manuel & Summers, Kyle 2011 |
Ranitomeya vanzolinii
| Souza, M. B. 2009: 31 |
| von May, R. & Catenazzi, A. & Angulo, A. & Brown, J. L. & Carrillo, J. & Chavez, G. & Cordova, J. H. & Curo, A. & Delgado, A. & Enciso, M. A. & Gutierrez, R. & Lehr, E. & Martinez, J. L. & Medina-Muller, M. & Miranda, A. & Neira, D. R. & Ochoa, J. A. & Quiroz, A. J. & Rodriguez, D. A. & Rodriguez, L. O. & Salas, A. W. & Seimon, T. & Seimon, A. & Siu-Ting, K. & Suarez, J. & Torres, J. & Twomey, E. 2008: 396 |
| Lotters, S. & Jungfer, K. - H. & Schmidt, W. & Henkel, F. W. 2007: 497 |
| Grant, T. & Frost, D. R. & Caldwell, J. P. & Gagliardo, R. & Haddad, C. F. B. & Kok, P. J. R. & Means, D. B. & Noonan, B. P. & Schargel, W. E. & Wheeler, W. 2006: 171 |
| Bauer, L. 1988: 1 |
Dendrobates vanzolinii
| Roberts, J. L. & Brown, J. L. & von May, R. & Arizabal, W. & Presar, A. & Symula, R. & Schulte, R. & Summers, K. 2006: 379 |
| Prohl, H. 2005: 358 |
| Christmann, S. P. 2004: 6 |
| Symula, R. & Schulte, R. & Summers, K. 2003: 452 |
| Caldwell, J. & de Oliveira, V. 1999: 565 |
| Schulte, R. 1999: 110 |
| Caldwell, J. P. 1997: 211 |
| Caldwell, J. P. & Myers, C. W. 1990: 4 |
| Myers, C. 1982: 9 |