
Ranitomeya amazonica Schulte 1999
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3083.1.1 |
|
persistent identifier |
https://treatment.plazi.org/id/1D338788-950D-150E-C8FC-9B033F23FB6B |
|
treatment provided by |
Felipe |
|
scientific name |
Ranitomeya amazonica Schulte 1999 |
| status |
|
Ranitomeya amazonica Schulte 1999 View in CoL
Account authors: E. Twomey, J.L. Brown, M. Pepper, M. Sanchez-Rodriguez
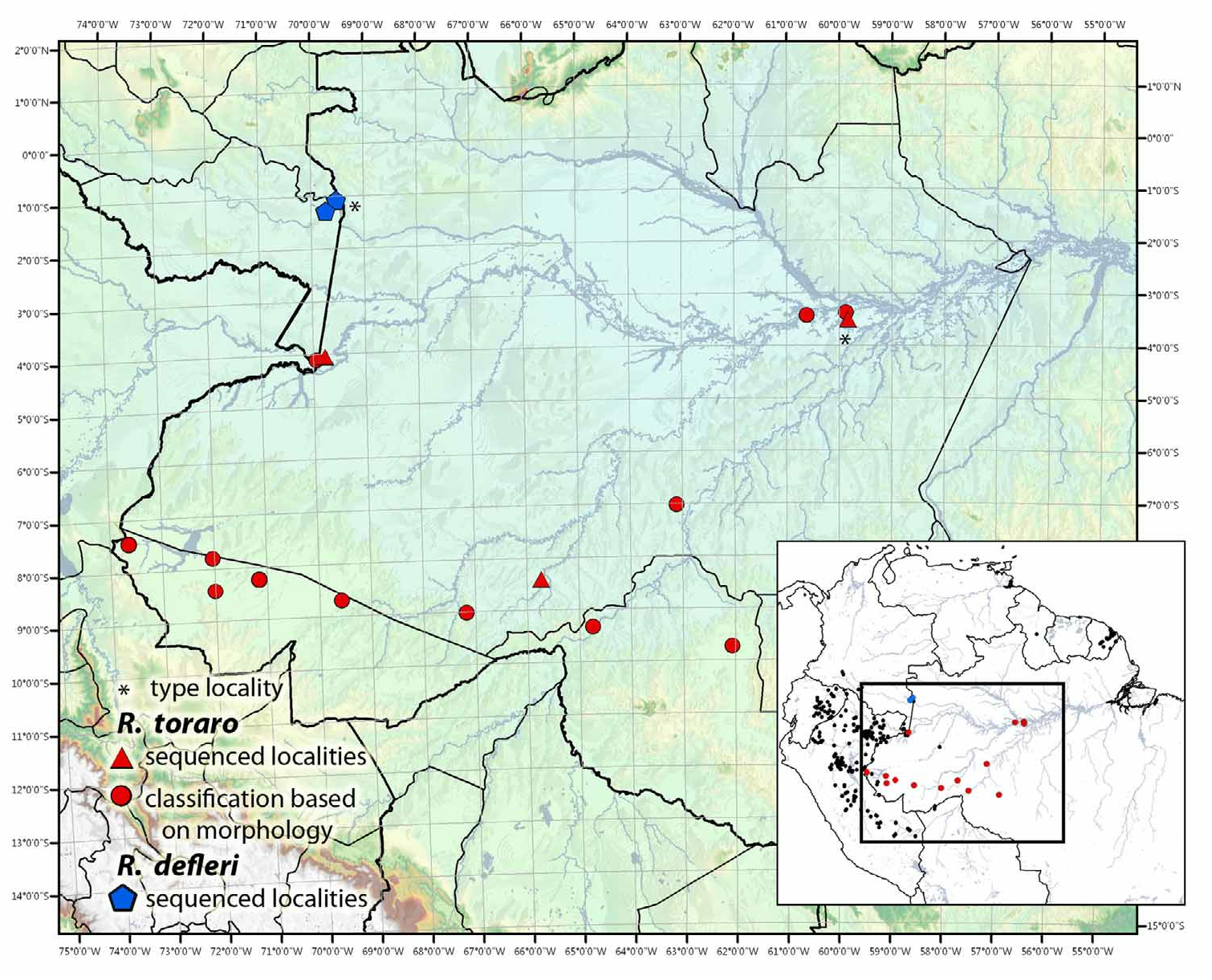
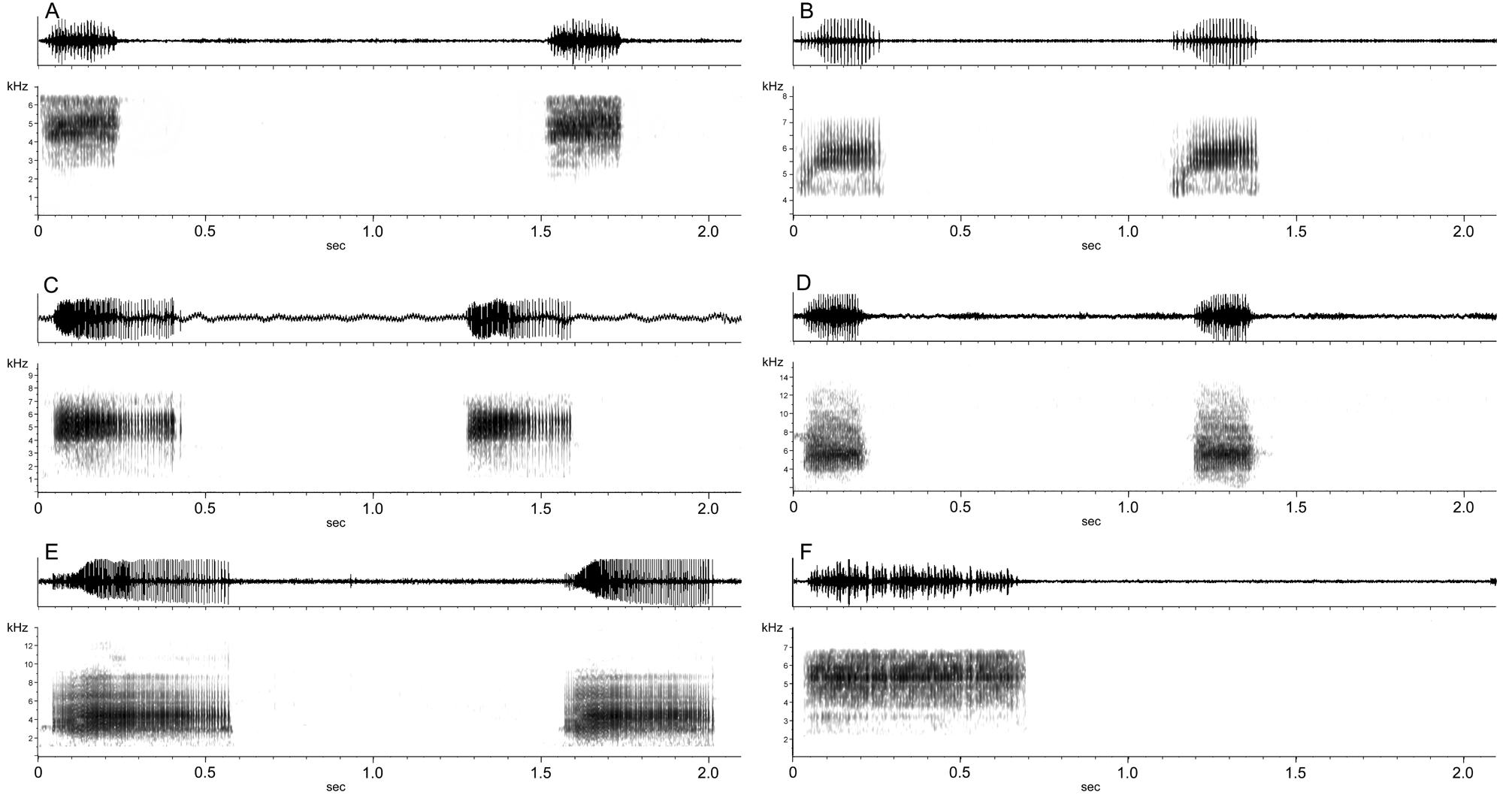
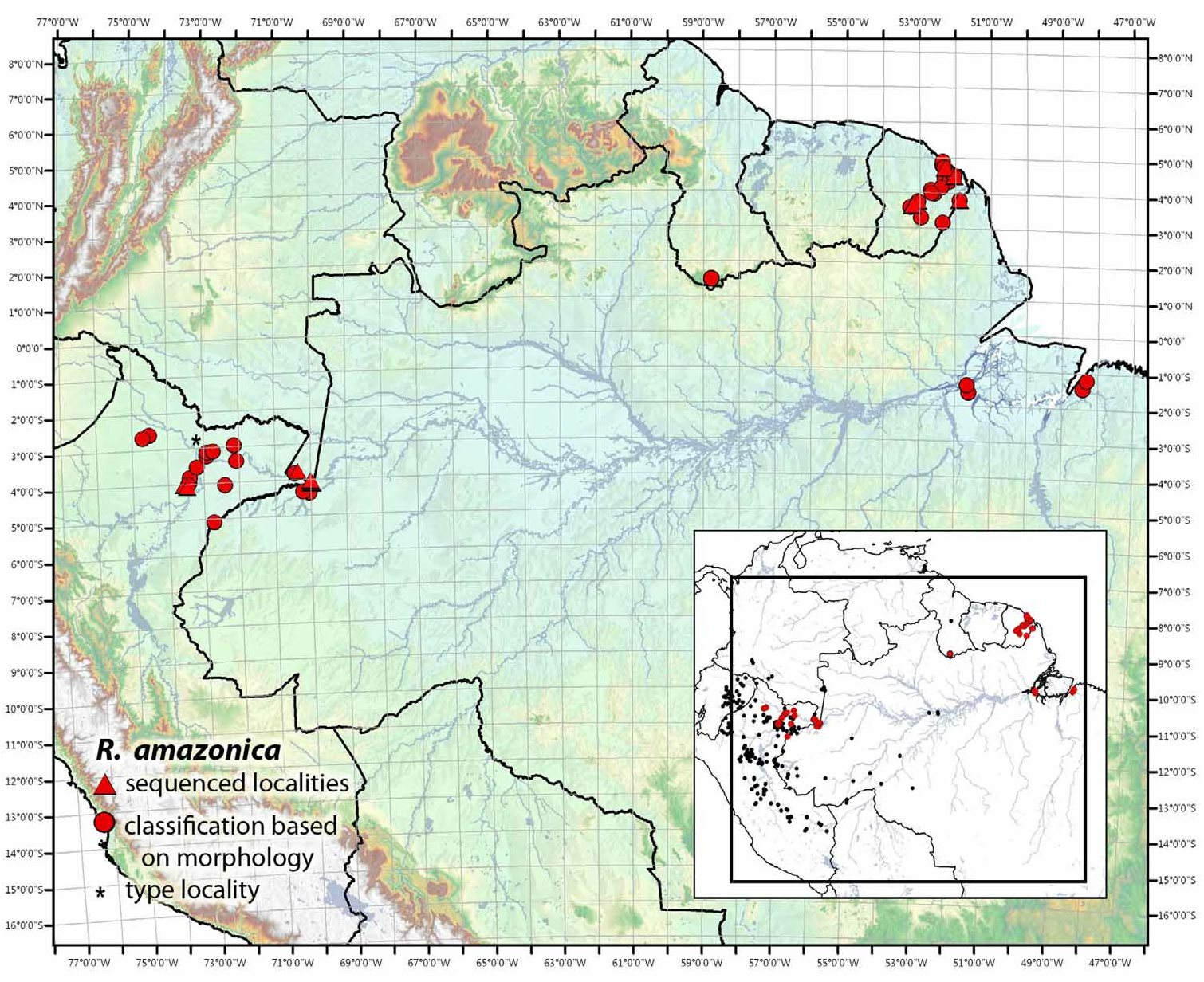
Figs. 3 View FIGURE 3 , 4 View FIGURE 4 , 9 View FIGURE 9 , 14 View FIGURE 14 , 34 View FIGURE 34 (l – q), 35 (a – q), 36
Tables 1, 4 – 6, 8
Dendrobates quinquevittatus View in CoL (non Steindachner 1864)—Silverstone 1975: p. 35 (partim) [ French Guiana specimens from Crique Ipoucin, Approuague drainage (LACM 42309); Camopi (MHNP 50-08); Saut Maripa, Fluve Oyapock (MHNP 50- 09; LG 73); Crique Gabrielle, Mahury drainage (LG 701); Mont Galbao (LG1422)]; Lescure & Bechter 1982: p. 26; Ávila-Pires & Hoogmoed 1997
Dendrobates ventrimaculatus View in CoL (non Shreve 1935)— Daly et al. 1987: p. 1025 (partim) [specimens from Mishana, Río Nanay drainage, Loreto, Peru]; Kok 2000: p. 19; Estupinan et al. 2001: Lescure & Marty 2001; Fig. 3 View FIGURE 3 ; Brown et al. 2006 (partim): p. 55, Table 2, Fig. 1; Christmann 2004: p. 6, Figs. on p. 23, 24, 42, 110, 112, 113; Roberts et al. 2006a (partim): p. 381, Table 1, Fig. 3 View FIGURE 3 ; Noonan & Wray 2006 (partim): p. 1009, Tables 1–2, Figs. 1, 4–5 View FIGURE 4 View FIGURE 5 ; Poelman & Dicke 2007: p. 217; 2008: p. 271
Dendrobates amazonicus Schulte 1999: p. 32 View in CoL , Figs. DB-047, PB-045, DB-073, [CRS BD 3 P (holotype) collected by Rainer Schulte at “Bosque UNAP, Iquitos ( PERU), ca. 130 m NN.” = Allpahuayo-Mishana National Reserve near Iquitos, Loreto, Peru, ca. 130 m. a.s.l];— Lötters & Vences 2000: p. 250; Symula et al. 2003: p. 453, Table 1; Christmann 2004: p. 17, Figs. on p. 17, 18, 20, 22, 42, 89, 90, 93, 94; Brown et al. 2006: p. 55, Table 2, Fig. 1; Santos et al. 2009, by implication
Ranitomeya amazonica View in CoL — Grant et al. 2006: p. 171, Fig. 76; Lötters et al. 2007: p. 463, Figs. 579–581
Ranitomeya ventrimaculata View in CoL (non Shreve, 1935)— Grant et al. 2006 (partim): p. 171, Fig. 76; Lötters et al. 2007 (partim): p. 504, Fig. 635; Perez-Peña et al. 2010 (partim): p. 18, Fig. 13 View FIGURE 13
Background information. Specimens of this species have been known at least as early as 1975 from material collected from several localities in French Guiana, when Silverstone (1975) referred to them as Dendrobates quinquevittatus View in CoL . In the same paper, he mentioned a number of specimens from the Iquitos region, although from his drawings and descriptions all these apparently belong to what is currently referred to as Ranitomeya reticulata View in CoL . Later, Myers (1982), after having worked with John W. Daly in Allpahuayo–Mishana near Iquitos, suggested that two species of small Dendrobates View in CoL were present in the area (both of which had been referred to as quinquevittatus View in CoL in Silverstone 1975). One of these was Dendrobates reticulatus Boulenger 1884 View in CoL “1883”; however, for the second species, he continued to apply the name Dendrobates quinquevittatus View in CoL . Caldwell & Myers (1990) began using the name D. ventrimaculatus View in CoL to refer populations of Dendrobates quinquevittatus sensu Silverstone View in CoL after the redescription of D. quinquevittatus View in CoL sensu stricto (now Adelphobates quinquevittatus View in CoL ), including the Allpahuayo–Mishana frogs. Schulte (1999) described the Allpahuayo-Mishana as a new species, D. amazonicus View in CoL , primarily on the basis of its advertisement call, which was said to be shorter and more highly pulsed than that of D. ventrimaculatus View in CoL . Schulte also included populations from French Guiana in the definition of amazonicus View in CoL on the basis of call similarities. This description was criticized by Lötters & Vences (2000), who focused on two issues: (i) the calls of both ventrimaculatus View in CoL and amazonicus View in CoL were not described on the basis of quantitative data; and (ii) the name amazonicus View in CoL was considered a possible junior synonym of D. igneus Melin 1941 View in CoL , described from the same general area. We have dealt with the name D. igneus View in CoL and concluded that it is a junior synonym of R. reticulata View in CoL (see account).
Studies using genetic data (e.g., Symula et al. 2001; Roberts et al. 2006a; Brown et al. 2006; Noonan & Wray 2006; Twomey & Brown 2008) supported Schulte’s (1999) conclusion that amazonicus was a taxon separate from what was called ventrimaculatus at that time (now considered R. variabilis ). Furthermore, putative populations of amazonicus from near Iquitos appeared to be closely related to individuals from French Guiana, further confirming Schulte’s (1999) arrangement. Still, the taxonomic status of amazonicus remained dubious because it rendered ventrimaculatus paraphyletic, yet at the same time there was speculation that ventrimaculatus may actually include several species, so no taxonomic actions dealing with amazonicus were made.
One of the main purposes of this monograph is to clarify the taxonomic status of R. ventrimaculata sensu Caldwell & Myers (1990) , while simultaneously assessing the validity of R. amazonica . Therefore, since 2004, we densely sampled individuals of both suggested taxa and obtained call recordings from various localities. The results of our phylogenetic and acoustic analyses largely support the conclusions of Schulte (1999). Individuals from near Iquitos and Río Napo in Peru, Leticia in Colombia and throughout French Guiana all form a monophyletic clade with relatively low genetic diversity. This clade is sister to a widespread, diverse clade containing individuals throughout much of the eastern Andean versant and parts of the upper Amazonian basin. The latter contains individuals previously referred to as R. ventrimaculata or R. variabilis (i.e., R. variabilis sensu this paper). Surprisingly, R. amazonica and R. variabilis sensu this paper occur in remarkably close contact south of Iquitos and may even be sympatric in some areas along the lower Río Tigre, Loreto, Peru (M. Sanchez-Rodriguez, pers. comm.).
Additionally, call differences do exist between these two species. Ranitomeya amazonica has (on average) significantly fewer pulses per note and a lower pulse rate than R. variabilis ( Table 8). However, interspecific overlap occurs in these parameters. As an example, R. variabilis from Cerro Yupati, Colombia and Shucushuyacu, Peru, had notes containing 19.5 and 20.6 pulses (two topotypic R. amazonica had notes containing 20.4 and 21.8 pulses). Also, when measuring pulse rate, one R. variabilis individual from Shucushuyacu had a pulse rate of 106 pulses/ note, while one R. amazonica individual from km. 41 on the Iquitos–Nauta road ( Peru) had a pulse rate of 108 pulses/note. We should note that Schulte (1999) described the call of R. amazonica as being shorter and more pulsed than that of R. variabilis sensu this paper. We did not find significant differences in note length, and the pulse-rate data indicate that R. variabilis (rather than R. amazonica ) has more highly-pulsed calls (Table 5).
Dorsal coloration and pattern do not appear to be good characters to distinguish R. variabilis and R. amazonica . Schulte (1999) recognized this by including individuals from French Guiana (which have yellow stripes) under his definition of R. amazonica , even though the populations near Iquitos (around km. 23 on the Iquitos–Nauta road) all have vibrant red or reddish-orange stripes. One of us (E. Twomey, unpub. data) has also found yellow individuals near Iquitos that fall within the amazonica clade, sampled at km. 41 on the Iquitos–Nauta road. Interestingly, individuals sampled from km. 81 (next to Nauta) all fall within the variabilis clade; these individuals are also yellow.
In addition to high variation in dorsal coloration in R. amazonica , their patterns have high levels of variation as well. In 2007, a population of R. amazonica was discovered by M. Pepper and M. Sanchez-Rodriguez that was strikingly similar in appearance to R. reticulata ( Fig. 35d – k View FIGURE 35 ). At the time of discovery, this population was thought to potentially represent a new species; however, after extensive genetic testing (see terminals labeled “Arena Blanca” in the phylogeny), we concluded these individuals were another morph of R. amazonica that may be involved in a mimicry system with the sympatric R. reticulata (see Mimicry section).
Biogeographically, R. amazonica represents somewhat of an enigma, although it appears that Schulte (1999) chose the specific epithet wisely, as this species appears to be affiliated with the Amazon River. Noonan and Wray (2006) pointed out that, while R. variabilis sensu this paper appears to be phylogeographically structured along a latitudinal axis, the phylogeographic structuring in R. amazonica appears to be along the Amazon River. Inclusion of additional samples from Río Napo ( Peru), Puerto Nariño ( Colombia) and Leticia ( Colombia) support this hypothesis. It is worth noting that there are large collection gaps between Leticia, Colombia and French Guiana, either explainable with the generally inadequate sampling in this region or natural absence (see Lötters et al. 2010 and discussion).
Definition and diagnosis. Assigned to the genus Ranitomeya due to the combination of the following characters: Small adult SVL (16 – 19 mm), dorsal coloration conspicuous, dorsolateral stripes highly variable and sometimes absent, but when present extend to top of thigh, ventrolateral stripes usually present (usually absent in ‘Arena Blanca’ populations), brightly colored throat, distinctive pale reticulation on limbs and venter, dorsal skin smooth, finger I is greatly reduced and shorter than finger II, finger discs II – IV greatly expanded, disc of finger 2.6 times wider than finger width, thenar tubercle conspicuous, toe discs III – V moderately expanded, toe webbing absent, larval vent tube dextral. Adults use arboreal phytotelmata for reproduction and deposit eggs above phytotelm, maxillary and premaxillary teeth absent. Two morphs are currently known of this species: (i) the Nominotypical morph ( Figs. 34 l – q View FIGURE 34 , 35 a – c, n – q View FIGURE 35 ), and (ii) the ‘Arena Blanca’ morph ( Fig. 35 d – m View FIGURE 35 ):
(i) The Nominotypical morph has a well-defined black ‘Y’ on the dorsum, with the two ends terminating just medial to the eyelids and the single end terminating on the posterior dorsum. In most individuals, a small black dot is present on the snout (as in Fig. 34m View FIGURE 34 ), but in some individuals this spot is absent (e.g., Fig. 35c View FIGURE 35 ). The dorsal stripe color is variable. Most populations around Iquitos have dorsal stripe coloration ranging from red to orange, but at least one individual in the area has been found with yellow dorsal stripe coloration. Most individuals have somewhat irregular dorsolateral stripes, and an incomplete middorsal stripe that terminates between the shoulders (i.e., between the ‘fork’ of the black ‘Y’). Ventrolateral and labial stripes are present and typically yellow. Limbs and venter are black with prominent pale reticulation that ranges from bright sky-blue to dark bronze-blue or gray. The throat is brightly colored, usually bright yellow or orange and has irregular black markings around the lateral edges. This morph occurs throughout much of the Amazon basin (see Distribution below). Red and orange individuals have been found from near Iquitos, Loreto, Peru near Río Napo and 70 km N Leticia, Amazonas, Colombia (T. Defler, pers. comm.). Yellow individuals have been found south of Iquitos, in Puerto Nariño, Leticia, throughout much of French Guiana and in extreme eastern Brazil near the Amazon river delta.
(ii) The ‘Arena Blanca’ (white sands) morph is a highly variable morph that may be involved in Müllerian mimicry with R. reticulata . This morph generally has a bright orange to red dorsum with irregular black markings. These markings can be manifested as partial stripes (i.e., Fig. 35e View FIGURE 35 ) or small spots (e.g., Fig. 35f View FIGURE 35 ). In extreme cases, these black markings can be attenuated, rendering the dorsal pattern nearly identical to that of R. reticulata (e.g., Fig, 35k). This morph is known only from a handful of sites in the Department of Loreto, Peru, exact locations undisclosed (available upon request).
Tadpole. The description is based on a single tadpole from lower Río Napo, Loreto, Peru. Mouthparts were verified in an individual from an unknown locality in French Guiana.
Tadpole in stage 29, no external gills, feeding on detritus. Body ovoid in dorsal view, wider near vent. Total length 16.6; body length 9.2; tail length 7.4, 45% of total length. Body width 7.6; body depth 5.3, 70% of body width. Eye well developed; naris small; distance from naris to anterior edge of eye 1.0. Eye positioned dorsally on head, directed dorsolaterally. Spiracle well developed, sinistral; vent tube dextral.
Tip of tail bluntly rounded. Tail muscle height at base of tail 2.5; tail muscle width at base of tail 1.9; maximum tail height 3.3. Dorsal and ventral fins approximately of same height.
Oral disc ventral, emarginated, transverse width 1.7, 22% of body width. Single row of small papillae present laterally and ventrally; dorsal gap where papillae absent. LTRF 2(2)/3(1) with A-1 developed on upper labium, A-2 with wide medial gap (one-third total width of tooth row); P-1 on lower labium with narrow medial gap; P-2 width equal to P-1; P-3 80% width of P-1.
Color in life. Head and body gray, mouthparts visible from above. Abdomen mostly transparent; intestinal coils black; heart visible. Tail musculature uniform gray, dorsal and ventral fins opaque gray.
Natural history. The natural history of R. amazonica (mainly French Guiana populations) has been described in detail elsewhere (e.g., Lescure & Bechter 1982; Schulte 1999; Lötters et al. 2007; Poelman & Dicke 2007, 2008); however, we will summarize some pertinent information here. Like other members of the variabilis group, R. amazonica breeds most frequently in bromeliads. Clutches consist of 2 – 6 eggs are placed in the axils of bromeliads partially submerged in water ( Lötters et al. 2007; Poelman & Dicke 2007). Males carry tadpoles to other bromeliad axils and cannibalistic tadpoles are deposited individually. These tadpoles are abandoned (i.e., no trophic egg-feeding) and feed on mosquito larvae, algae, detritus or other tadpoles that may be unwittingly deposited in the same phytotelm.
In French Guiana, this species was observed changing its food provisioning strategy. Early in the breeding season, tadpoles subsist predominantly on food within the phytotelm, whereas later in the breeding season (apparently due to increased desiccation risk), parents deliberately deposit embryos as a food source in pools containing their offspring to ensure quick metamorphosis ( Poelman & Dicke 2007, 2008).
Vocalizations. The call of R. amazonica consists of a series of short buzz notes, note range 0.16 – 0.36 sec in duration, repeated at a rate of 24 – 70 notes per minute.
Distribution. This species occurs in Amazonian rainforests of Brazil (States: Amapá, Amazonas, Pará), Colombia (Department: Amazonas), Guyana (Region: Upper Takutu-Upper Essequibo), French Guiana (Arrondissements: Cayenne, Saint-Laurent-du-Maroni) and Peru (Department: Loreto), Fig. 36 View FIGURE 36 .
Conservation Status. Following the IUCN Red List categories and criteria ( IUCN 2010), we suggest listing this species as Least Concern (LC).





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Ranitomeya amazonica Schulte 1999
| Brown, Jason L., Twomey, Evan, Amézquita, Adolfo, Souza, Moisés Barbosa De, Caldwell, Jana- Lee P., Lötters, Stefan, May, Rudolf Von, Melo-Sampaio, Paulo Roberto, Mejía-Vargas, Daniel, Perez-Peña, Pedro, Pepper, Mark, Poelman, Erik H., Sanchez-Rodriguez, Manuel & Summers, Kyle 2011 |
Ranitomeya amazonica
| Lotters, S. & Jungfer, K. - H. & Schmidt, W. & Henkel, F. W. 2007: 463 |
| Grant, T. & Frost, D. R. & Caldwell, J. P. & Gagliardo, R. & Haddad, C. F. B. & Kok, P. J. R. & Means, D. B. & Noonan, B. P. & Schargel, W. E. & Wheeler, W. 2006: 171 |
Dendrobates amazonicus
| Brown, J. L. & Schulte, R. & Summers, K. 2006: 55 |
| Christmann, S. P. 2004: 17 |
| Symula, R. & Schulte, R. & Summers, K. 2003: 453 |
| Lotters, S. & Vences, M. 2000: 250 |
| Schulte, R. 1999: 32 |