
Rhinopycnodus gabriellae, Taverne & Capasso, 2013
|
publication ID |
https://doi.org/ 10.5852/ejt.2013.67 |
|
publication LSID |
lsid:zoobank.org:pub:2358BCCE-7E9E-445D-B4B6-BBB56453055B |
|
DOI |
https://doi.org/10.5281/zenodo.3846649 |
|
persistent identifier |
https://treatment.plazi.org/id/D5399E6C-45DF-4D7D-BA77-545D6014BE22 |
|
taxon LSID |
lsid:zoobank.org:act:D5399E6C-45DF-4D7D-BA77-545D6014BE22 |
|
treatment provided by |
Carolina |
|
scientific name |
Rhinopycnodus gabriellae |
| status |
gen. et sp. nov. |
Rhinopycnodus gabriellae gen. et sp. nov.
Figs 1-8 View Fig View Fig View Fig View Fig View Fig View Fig View Fig View Fig
urn:lsid:zoobank.org:act:D5399E6C-45DF-4D7D-BA77-545D6014BE22
Diagnosis
Primitive pycnodontid characterized by a long and broad premaxilla bearing one short and very broad tooth. Elongated head with a long preorbital region. Dentary bearing 2 small incisiform teeth. No temporal fossa. Exoccipital-basioccipital region well visible behind the dermopterotic. Mouth gape obliquely oriented. Preopercle larger than the exposed region of the hyomandibula-dermohyomandibula. Maximum body depth: 67.6 % of the SL. Pectoral fin with 9 rays. Ventral fin with 3 rays. Dorsal fin with 49 pterygiophores. Origin of the dorsal fin behind the dorsal apex and at 76.2 % of the SL. Anal fin with 47 rays and 45 pterygiophores. Origin of the anal fin on the ventral apex and at 63.8 % of the SL. Neural and haemal arches almost completely surrounding the notochord. 32 vertebral elements (neural spines) before the epichordal series. Neural spines 1-8 autogenous. 11 pairs of ribs. Postcoelomic bone reaching the axial skeleton and the ventral margin. 15 haemal spines before the hypochordal series. 7 epichordal and 11 hypochordal elements in the caudal skeleton. Caudal fin with 30 principal rays. Body scales only in the abdominal region. Complete scales ventrally and scale bars dorsally. 19 dorsal ridge scales. First dorsal ridge scale small, triangular in shape and not sutured to the dermosupraoccipital. Some spiny scales in the dorsal ridge and in the ventral keel. 3 pelvic scales. 2 postcloacal scales.
Etymology
The species name of the new Lebanese fossil fish is dedicated to Mrs. Gabriella di Tota, the co-author’s wife.
Formation and locality
Marine Upper Cenomanian, Haqel, Lebanon.
Holotype and unique specimen
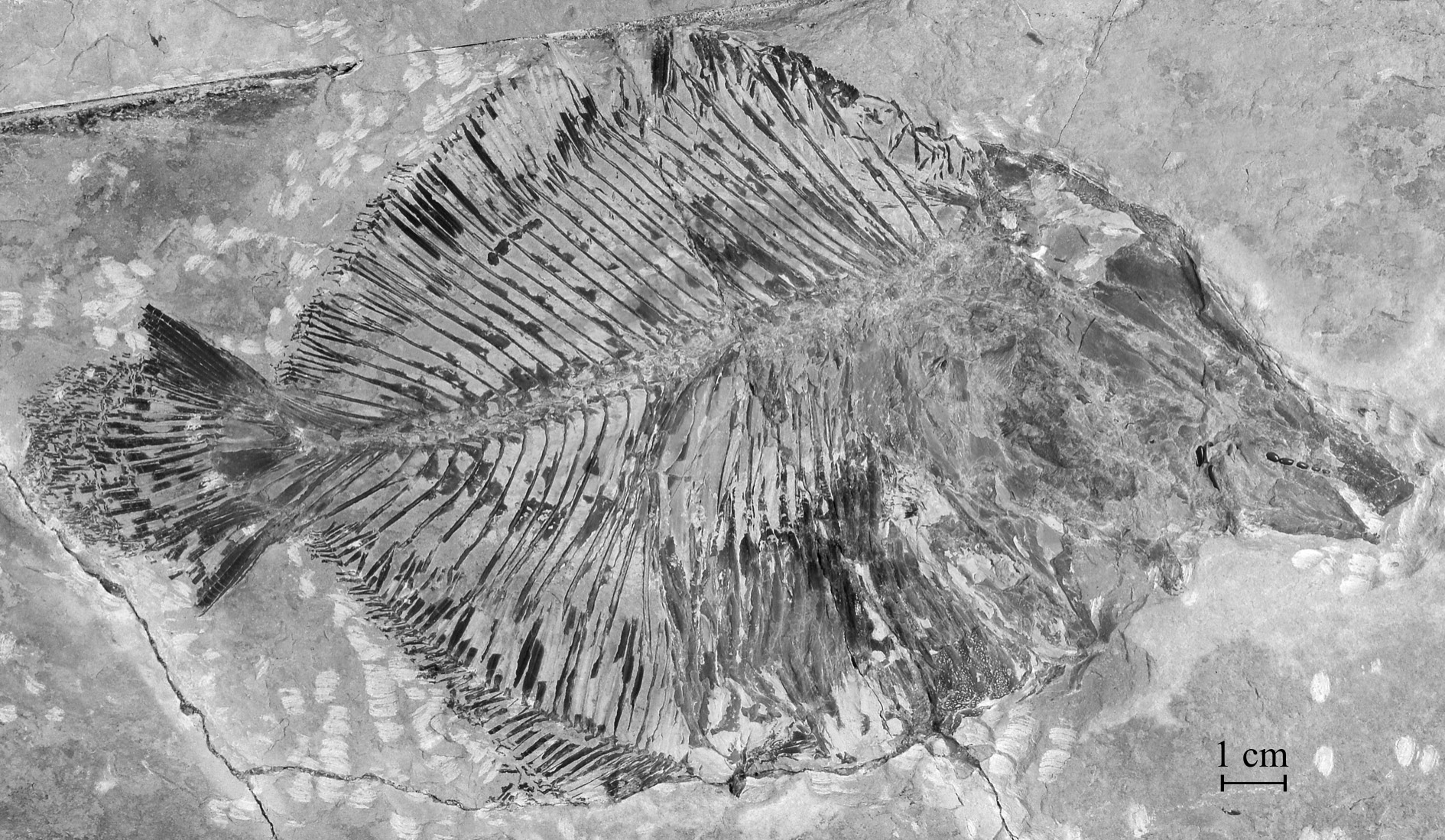
Sample CLC S-725 , a complete specimen seen by its right side ( Figs 1 View Fig , 2 View Fig ) from Haqel, Lebanon. Total length: 223 mm.
Holotype morphometric data
The morphometric data are given in % of the standard length (183 mm) of the holotype.
Length of the head (opercle included) ……………………………………………………………45.2 % Depth of the head (in the occipital region) ………………………………………………………36.7 % Maximum depth of the body (at the anal fin origin level) ………………………………………67.6 % Prepelvic length …………………………………………………………………………………52.4 % Predorsal length …………………………………………………………………………………76.2 % Basal length of the dorsal fin ……………………………………………………………………39.0 % Preanal length ……………………………………………………………………………………63.8 % Basal length of the anal fin ………………………………………………………………………40.0 %
Osteology
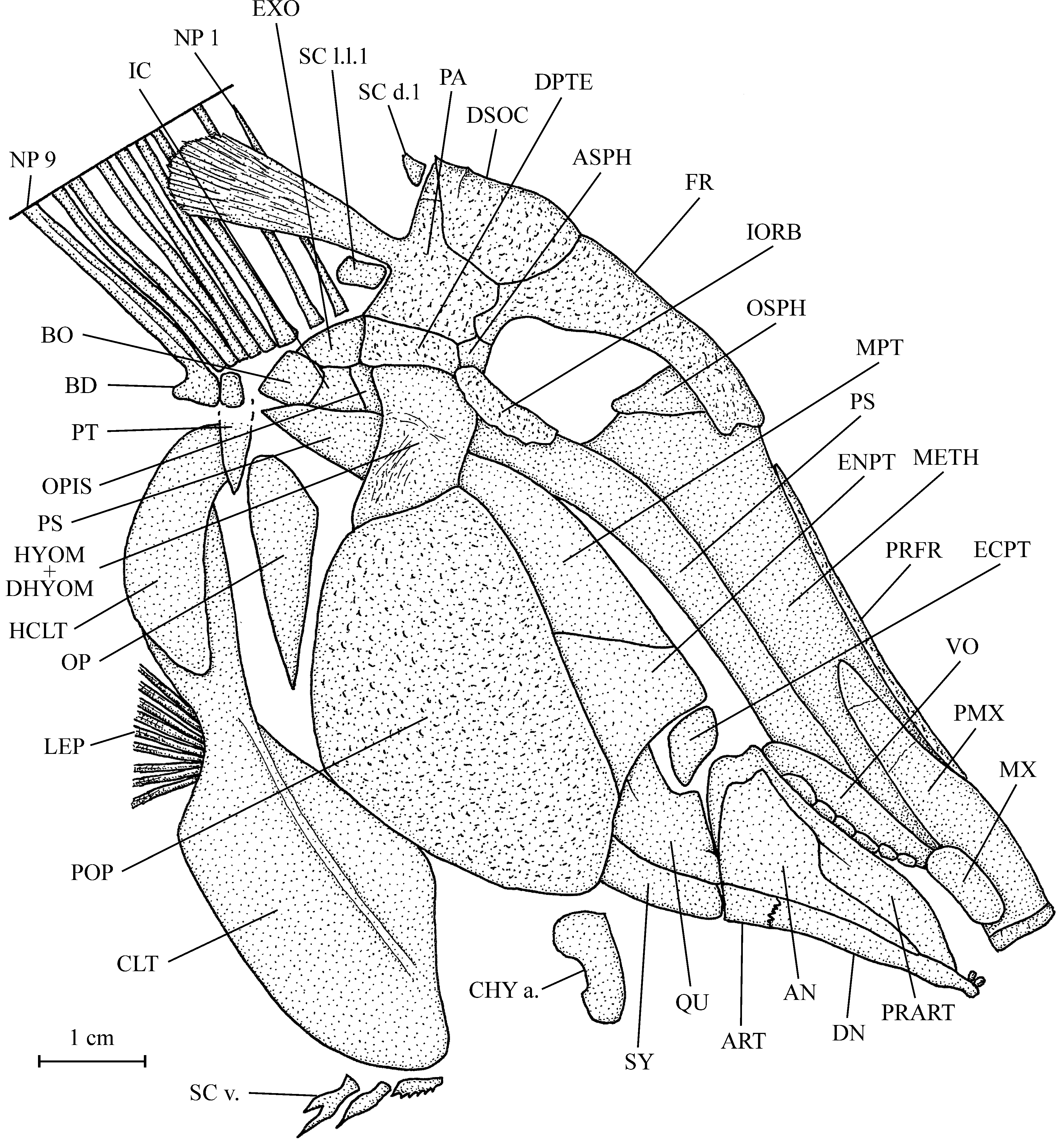
1. The skull ( Figs 3 View Fig , 4 View Fig )
The head is high, with the preorbital region much longer than the orbital-postorbital region. The dermal bones of the skull are slightly ornamented with small granulations. The mouth gape is inclined ventrally.
The mesethmoid is broad, very long and its upper margin is covered by a pair of long and very narrow prefrontals. The vomer is seen in profile and only six rounded molariform teeth ranged in one rank are visible. The frontal is rather short and not very broad. The posterior margin of the bone is a little enlarged and meets the dermosupraocciptal, the parietal and the small autosphenotic, but not the dermopterotic. Posteriorly, the dermosupraoccipital ends in a short pointed tip. The parietal bears a large posterior brush-like process. There is no temporal fenestra. The dermopterotic is not deepened and is located at the level of the upper border of the orbit. The opisthotic, intercalar, basioccipital and exoccipital are visible behind the dermopterotic and the hyomandibula. A small orbitosphenoid is pressed against the posterior border of the mesethmoid. The parasphenoid is very long and its trabecular region is obliquely oriented. Posteriorly, the parasphenoid reaches the level of the basioccipital. The sensory canals on the braincase are not visible.
The quadratic arch contains a well developed quadrate, a large metapterygoid, a large entopterygoid and a small ectopterygoid. The quadrate and the symplectic are both articulated on the lower jaw.
The premaxilla is long and very broad. It bears only one short but very broad tooth. The maxilla is small, deeper than long, reniform, toothless and pressed against the premaxilla by its upper margin. When seen in profile, the upper jaw is hog snout-like because of the broadening of the premaxilla and of its tooth. The lower jaw is small and triangular in shape. The dentary is reduced to its ventral branch and it bears two small incisiform teeth. The angular covers a great part of the external face of the mandible. The small articular and the dentary are articulated together. The coronoid process of the prearticular is well marked. Three deep molariform teeth fallen from the prearticular are visible just behind the jaw.
The last infraorbital is long, well ossified and forms the posterior and ventral margins of the orbit. No other infraorbital is preserved. The dermosphenotic is lost.
The preopercle is the largest bone of the skull, all together deep and broad. The hyomandibula and dermohyomandibula are fused and their exposed part is important but, however, much smaller than the preopercle. The opercle is well developed, with a pointed ventral tip and a broader upper part. No branchiostegal ray is preserved.
The anterior ceratohyal is the only preserved part of the hyoid bar. A few hook-like branchial teeth and some branchial filaments are visible under some broken regions of the opercle and preopercle.
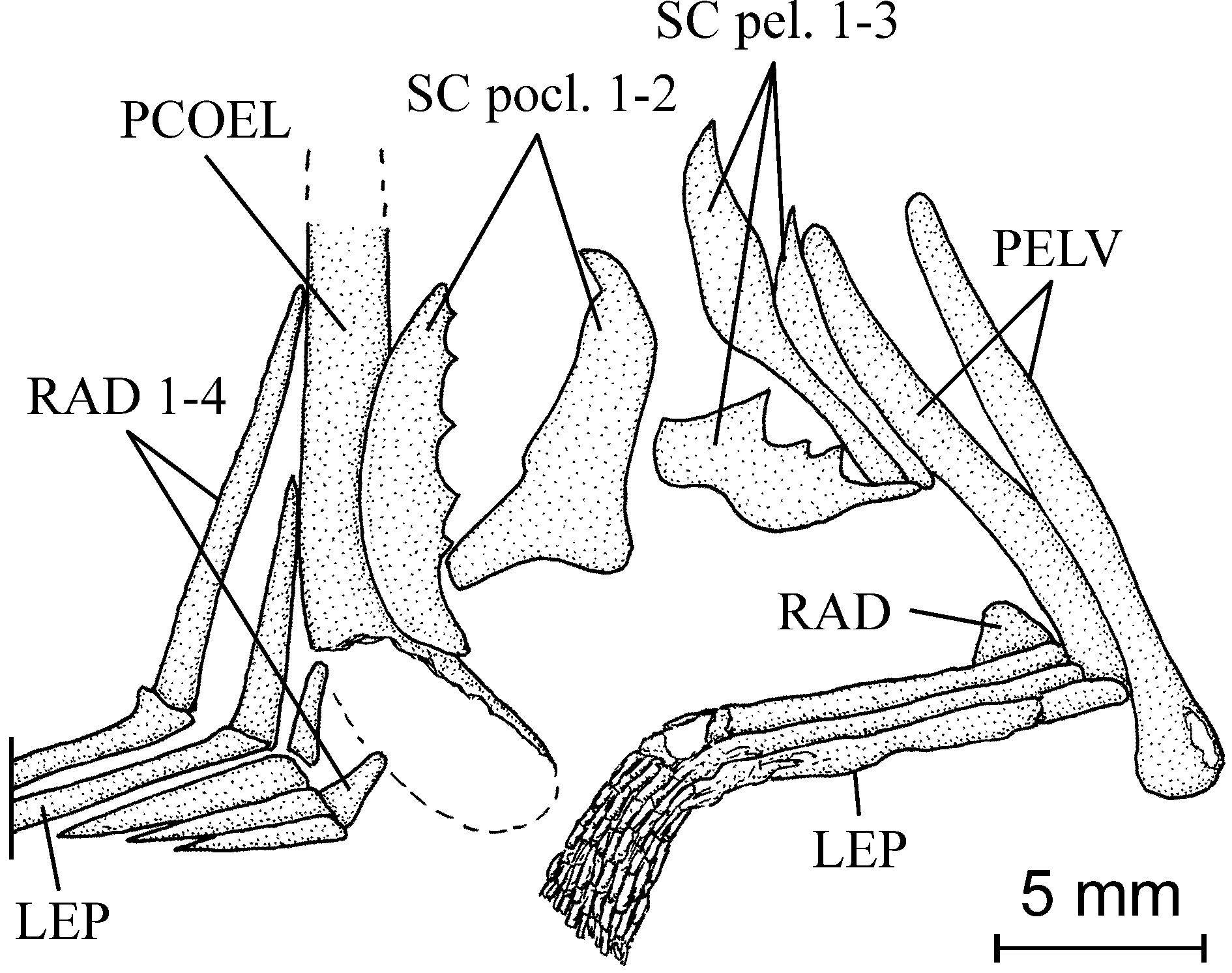
2. The girdles ( Figs 3-5 View Fig View Fig View Fig )
Only the ventral part of the posttemporal is preserved. The hypercleithrum (= supracleithrum) is deep, rather broad and not splint-like as usual in Pycnodontiformes . The cleithrum is a large bone with a broad palaform ventral branch and a sinus in its posterior margin to house the pectoral fin, which is short and contains a least 9 rays.
The two pelvic bones are vertically oriented. The ventral fins are rather long. Each of them contains 3 rays. The origin of the ventral fins is located a little before the midpoint of the ventral contour.
3. The axial skeleton ( Fig. 2 View Fig )
Starting from the caudal region, the vertebral axis progressively elevates to reach the level of the orbit anteriorly. The vertebrae are constituted by only the dorsal and ventral arcocentra. No chordacentrum or autocentrum is visible. The neural and haemal arches surround the notochord almost completely. There are 32 neural spines before the epichordal series and thus 32 vertebral segments before the elements involved in the caudal fin support. The first 17 vertebral segments are abdominal and the following 15 caudal. The anteriormost 8 neural spines are autogenous. The first seven lean on the occipital region of the braincase and the eighth spine is located just above the first ossified but small basidorsal. The basiventrals are strongly reduced in the abdominal region but well developed in the caudal region. There are 11 pairs of long alate ribs and 15 haemal spines before the hypochordal series. The two last pairs of ribs are noticeably shorter than the nine preceding ribs. The first three haemal spines are pressed together and against the postcoelomic bone. The first haemal spine is incomplete and does not reach the axial skeleton. The neural and haemal spines bear an anterior sagittal flange, except for the first five neural and the first three haemal spines. A well developed postzygaphophysis links each neural and haemal arch with the following one. The postcoelomic bone is a long and robust bone dorsally reaching the axial skeleton and ventrally the lower margin of the abdomen.
4. The dorsal and anal fins ( Fig. 2 View Fig )
The dorsal fin begins a little behind the dorsal apex. The fin is supported by 49 long and strong pterygiophores. The first seventeen of them have lost the corresponding rays. The last thirty two pterygiophores bear short segmented and branched rays. The first two pterygiophores are broader than the following ones.
The anal fin is strip-like in shape (type A2 of Poyato-Ariza & Wenz 2002: fig. 34). The origin of the fin is located at the ventral apex. There are 45 strong pterygiophores bearing 47 rays. The first five pterygiophores abut against the postcoelomic bone and are progressively lengthened from the first to the fifth. The first three rays are reduced to short spines. The other rays are segmented and branched. The very short first pterygiophore supports two little spiny rays and the last pterygiophore two soft rays.
5. The caudal skeleton ( Figs 6 View Fig , 7 View Fig )
There is no caudal peduncle because the dorsal and anal fins reach the caudal fin.The caudal endoskeleton contains 1 urodermal, 7 epichordal and 11 hypochordal elements. The first epichordal neural arch bears a long and narrow neural spine but the length of the neural spines progressively decreases from the first to the seventh epichordal element, which has only a very short neural spine. Some elements in the hypochordal series are broadened but there is no real hypertrophy. This broadening is more important on the eighth and the tenth hypochordal elements than on the other parts of the series.
The contour of the caudal fin is double emarginated ( Poyato-Ariza & Wenz 2002: fig. 36 E) but the median convex part of the fin is greatly enlarged. There are 30 principal rays, 3 dorsal and 4 ventral procurrent rays.
6. Squamation ( Figs 2 View Fig , 4 View Fig , 5 View Fig , 8 View Fig )
The dorsal ridge and ventral keel scales are notably differentiated from the flank scales.
There are 19 dorsal ridge scales between the dermosupraoccipital and the origin of the dorsal fin but only the first and the seventh to the tenth are well preserved. The first dorsal scale is small, triangular in shape and located just behind the dermosupraoccipital. The five following scales are badly crushed. Only fragments of the last nine are visible in a fissure of the substratum at the dorsal apex level. Each upper margin of the seventh to tenth dorsal scales bears up to six small spines.
The total number of ventral ke el scales is unknown. The ventral keel begins with 3 scales located under the cleithrum ( Fig. 4 View Fig ). The first one bears a few very small spines. The second one has a large spine and the third one two large spines. A few ventral keel scales bearing very small spines are visible on the
ventral contour between the cleithrum and the pelvic girdle, but some elements of this series are lost because of the taphonomic events.
There are 3 pelvic scales associated with the pelvic bones and 2 postcloacal scales are located just before the postcoelomic bone.
There are flank scales only in the abdominal region of the body, anterior to the origin of the dorsal and anal fins. In the ventralmost area of the situs viscerum, between the cleithrum and the postcoelomic bone, the scales are complete, thick, deep, broad, slightly ornamented with small tubercules and articulated together. There are 11 rows of these large ventral flank scales. The other body scales are reduced to scale bars. In the dorsal area of the abdominal region the scale bars are badly preserved and only fragments are visible between the neural spines. Scale bars also are associated with the first eight dorsal ridge scales. The scales linked to the eleven other dorsal ridge scales are progressively broader and longer.
The first scale of the lateral line is visible beneath the brush-like process of the parietal.





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |