
Glyphocrangon taludensis, Hendrickx, 2010
|
publication ID |
https://doi.org/ 10.11646/zootaxa.2372.1.27 |
|
persistent identifier |
https://treatment.plazi.org/id/207687FE-BF4B-475C-FF62-FC2CFED68E0B |
|
treatment provided by |
Felipe |
|
scientific name |
Glyphocrangon taludensis |
| status |
sp. nov. |
Glyphocrangon taludensis View in CoL n. sp.
( Figs. 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 )
Material examined. Holotype: ov. female (cl 34.3, tl 128.1), TALUD XII, St. 25 (18°26’45”N, 104°16’10”W) 1.IV.2008, 1858– 1879 m depth, benthic sledge, muddy bottom ( EMU –8595) GoogleMaps . Allotype: 1 male (cl 21.2, tl 85.2), TALUD VII, St. 13B (23°30.3’N, 107°44.0’W) 6.VI.2001, 1400–1450 m depth, benthic sledge, muddy bottom ( EMU –8596) GoogleMaps . Paratypes: 2 ov. females (cl 32.4–35.1, tl 124.2–128.2), TALUD XII, St. 25 (18°26’45”N, 104°16’10”W) 1.IV.2008, 1858– 1879 m depth, benthic sledge, muddy bottom ( EMU –8597; 1 ov. female (cl 39.3, tl 141.5), TALUD XII, St. 25, (18°26’45”N, 104°16’10”W) 1.IV.2008, 1858– 1879 m depth, benthic sledge, muddy bottom ( EMU –8598) (preserved in ethanol) GoogleMaps .
Additional material examined. One female (cl 27.5, tl 104.5), 2 ov. females (cl 28.1–28.4, tl 107.0– 107.6) ( EMU –8601, 8604), TALUD VII, St. 13B (23°30.3’N, 107°44.0’W) 6.VI.2001, 1400–1450 m depth, benthic sledge, muddy bottom; 1 ov. female (cl 28.3, tl 104.9), TALUD VII, St. 25 (24°51.8’N, 108°58.0’W) 8.VI.2001, 780– 850 m depth, benthic sledge, muddy bottom ( EMU –8602); 1 male (cl 19.0, tl 79.9) ( EMU – 8600), 3 ov. females (cl 26.5–29.4, tl 106.3–111.6) ( EMU –8599, 8603), TALUD XII, St. 29 (19°19’37”N, 105°26’20”W) 2.IV.2008, 1609– 1643 m depth, benthic sledge, muddy bottom GoogleMaps .
Type locality. Off Guerrero, Mexico (18°26’45”N, 104°16’10”W), 1858–1879 m depth GoogleMaps .
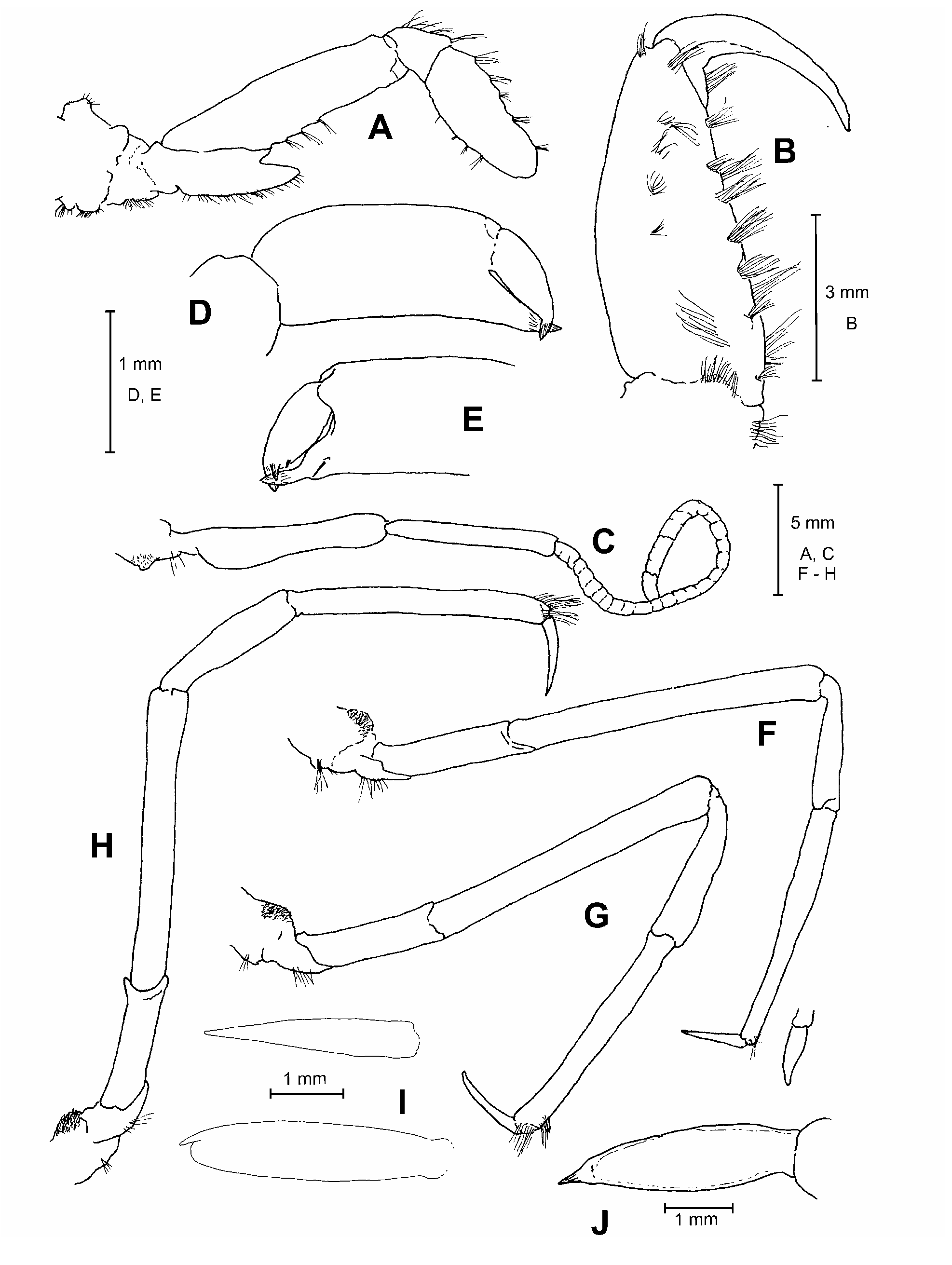
Description. Body ( Figs. 1–2 View FIGURE 1 View FIGURE 2 ) robust, integument firm, with very short pubescence, not visible to nakedeye. Rostrum slightly upturned anteriorly, overreaching anterior margin of scaphocerite (0.57 times as long as carapace in holotype, 0.62 in allotype and 0.60–0.67 in paratypes), dorsally concave, not septate; dorsolateral margins with 2 pairs of subequal teeth, anterior pair at proximal 1/3 of rostrum, posterior pair at posterior margin of orbit; median carina extending from apex of rostrum to between the two pairs of lateral teeth, low, not higher than lateral carinae; dorsolateral and ventrolateral margins ridged; ventral surface flattened, with short distal, median carina.
Submedian (first) carina on carapace composed of blunt tubercles, slightly pointing forward, 7 or 8 anterior and 5 or 6 posterior to cervical groove, area between submedian carinae smooth except for few anterior tubercles. Anterior intermediate (second) carina composed of 3 strong tubercles and strong triangular tooth continuous with dorsolateral carina of rostrum; posterior intermediate carina strong, not entire, margin divided into 5 or 6 lobes, a pair of small, rounded tubercles anterior to carina. Anterior antennal (third) carina very short, not continuous with antennal tooth, reduced to row of close-set 2 or 3 small eroded tubercles; posterior antennal carina entire, strong, eroded, except for lower anterior portion, without anterior lobe or tooth, a couple of strong tubercles oblique to carina beyond cervical groove. Anterior lateral (fourth) carina not continuous with antennal tooth, a distinct notch at about midlength, anterior section forming large tooth not reaching level of posterior margin of orbit, posterior section terminating anteriorly in low, blunt tooth; posterior lateral carina entire, eroded. Anterior sublateral (fifth) carina prominent, eroded; posterior sublateral carina less distinct. Submarginal carina less distinct, separated into some sections. Fifth and sixth carinae margins beyond cervical groove subparalel to lower margin of carapace, ending in a strong posterior, triangular lobe just above posterolateral angle of carapace. Space between anterior first and second carinae with a few tubercles; space between posterior first and second carinae with scattered tubercles; spaces between posterior parts of second, third and fourth carinae smooth. Antennal teeth unarmed, about 3/4 as long as, and diverging more than branchiostegal teeth. Branchiostegal teeth not overreaching anterior margin of cornea, falling short of proximal segment of antennular peduncle, slightly divergent, two low lateral and one ventral carinae.
First abdominal somite with some obscure tubercles along posterior margin, 1 strong flattened tubercle slightly produced beyond anterolateral margin of tergum; median carina thick, dorsal margin ridged, overhanging anterior section of first somite. Median carina on each somite posterior to first sharp, divided into anterior and posterior sections by shallow notch (somites 2–4) or v-shaped incision (somites 5–6). Fifth somite with posteriorly divergent submedian carina on posterior half. Posterior margins of fourth to sixth somites convexly produced. Pleuron of somites 2–5 with mesial, vertical protuberance, bearing several irregular tubercles, the lower one sharper, tooth-like in somites 3–4. Pleuron of first somite tapering anteroventrally to blunt point, and those on somites 2–5 with 2 ventral teeth; teeth on somite 2 directed ventrally, anterior slightly larger than posterior tooth; teeth on somites 3–5 directed posteroventrally, anterior stronger than posterior in somites 3–4, weaker than posterior one in fifth somite. Pleuron of sixth somite with strong posteroventral tooth directed posteriorly. Telson ( Fig. 1 View FIGURE 1 ) elongate, triangular, gradually tapering to sharp point, 0.63 times as long as carapace, posterior part slightly uptumed; dorsolateral margin sharply ridged; dorsal surface concave, with strong, acute median tubercle proximally.
Eye moderately large, about 1.28 rostral width at its base, about 0.25 carapace length, with dark cornea.
Antennule with peduncle falling slightly short of anterior margin of scaphocerite; proximal segment with stylocerite showing as acute lobe; combined length of 2 distal segments less that 2/3 length of proximal segment. Antennular flagella about as long as peduncle.
Scaphocerite ( Fig. 4D–E View FIGURE 4 ) ovate, 1.55 times as long as wide, less than half the length of carapace, with minute lateral tooth at about midlength, lateral margin proximal to lateral tooth bearing short setae. Carpocerite not reaching distal margin of blade.
Third maxilliped ( Fig. 4A–B View FIGURE 4 ) stout, reaching anterior margin of scaphocerite; distal 2 segments with strong spines on ventromesial margin and mesial face, mesial face concealed by long setae; ultimate segment terminating in sharply pointed apex; exopod with articulated distal lash.
First pereopod ( Fig. 3A–B View FIGURE 3 ) incompletely subchelate; palm narrowing distally in dorsal view, with row of tufts of stout setae mesially; carpus short, bearing setae dorsomesially; ischium with wide ventral laminar expansion, distally pointed. Right second pereopod slightly longer than left ( Fig. 3C View FIGURE 3 ) and with more carpal articles (28 vs. 25) (in paratypes, right with 25–27, left with 20–23); chela ( Fig. 3E–D View FIGURE 3 ) about as long as ultimate article of carpus; fixed finger short, terminating in corneous spine; dactyl broad, terminating in 2 unequal corneous spines; ischium distinctly longer than merus, ventral margin somewhat expanded. Third pereopod ( Fig. 3F View FIGURE 3 ) with dactyl flattened, simple, less than 1/3 propodus length, without setae except for a few terminal setae on propodus; carpus 0.5–0.6 times as long as propodus. Fourth ( Fig. 3G View FIGURE 3 ) and fifth pereopods ( Fig. 3H View FIGURE 3 ) with dactyl similar to that of third pereopod, proportionally slightly longer, without setae; propodus without setae, except for terminal and subterminal tufts of setae; carpus 0.6 times as long as propodus, without setae. Male dactyl of fourth and fifth pereiopods bifid ( Fig. 3I View FIGURE 3 ).
Male first pleopod with endopod ( Fig. 4F–G View FIGURE 4 ) slightly less than half length of exopod, mesial margin concave; appendix interna defined mesially by wide V-shaped sinus, oval. Male second pleopod ( Fig. 4H View FIGURE 4 ) with appendix masculina longer than appendix interna, by over 1/3 appendix interna length, bearing about 40 long spines.
Uropod ( Fig. 1 View FIGURE 1 ) not reaching posterior end of telson; exopod equal in length to endopod, with transverse suture, lateral margin convex, terminating posteriorly in acute tooth.
Eggs large, ovate. Number of eggs increasing with carapace size: 110 (cl 28.1, EMU–8601); 134 (holotype, cl CL); 172 (cl 39.3, EMU–8598). Size of eggs increasing with embryonal development (estimated by direct microscope observation): 2.30 × 2.97 in eggs with no sign of embryo; 2.60 × 3.29 in eggs with small embryo; 2.83 × 3.71 in eggs with large embryo.
Etymology. The species is named after the name of the cruises and the research project during which it was collected (TALUD cruises).
Ecology. The material examined was collected in depths between 780 and 1879 m. Epibenthic temperature and dissolved oxygen concentration values varied from 2.4 to 4.7°C and from 0.10 to 1.85 ml O 2 / l. The lowest oxygen value was measured at station 25 of the TALUD VII cruise, in depths of 780–850 m, a bathymetric fringe where severe hypoxy is known to occur in the southern Gulf of California and off the coast of western Mexico (see Hendrickx 2001; Hendrickx & Serrano 2007).
Remarks. Glyphocrangon taludensis sp. nov. is morphologically close to G. sicaria Faxon, 1893 and G. vicaria Faxon, 1896 , both occurring in the east Pacific. It differs from both species, however, by the absence of tubercles between the lateral carinae, the presence of a large tooth at the anterior end of the second anterior lateral carina, the size of the eye (proportionally smaller in G. vicaria ), the presence of a deep V-shaped notch on dorsal carina of somites 5–6 (absent in G. vicaria , little pronounced in G. sicaria ), and the presence of two large rostral lateral teeth (proximal obsolete in G. sicaria , reduced in G. vicaria ). The absence of tubercles between the lateral carinae also distinguish G. taludensis sp. nov. from two other species similar to G. vicaria , namely G. saintlaurentiae Komai, 2004 (West Pacific), and G. atlantica Chace, 1939 (Atlantic Ocean). The characters used by Komai (2004a: key) to distinguish the Indo-Pacific species and comparison of other species subsequently described by this author ( Komai 2006, 2007; Komai & Chan 2008) allow for the distinction of G. taludensis sp. nov. from all other known species in the genus, by the following characters: the carapace and abdomen feature very short, sparse setae, not visible to the naked-eye; the intercarinal regions on the carapace are devoid of rows or clusters of tubercles (except for occasionally a few, isolated tubercles); there is a single lateral rostral tooth above the orbital margin and the rostrum is not septate; the pterygostomial tooth is distinctly longer than the antennal tooth; the eye is large, about 0.25 times length of carapace; the first (submedian) carina harbours blunt tubercles; the anterior lateral (fourth) carina is divided into teeth or lobes and does not reach the orbital margin; the anterior (third) carina is confined to the antennal spine, with small tubercles forming a reduced row on the upper hepatic region; the dactylus of pereiopods 4–5 is subspatulate, terminating in a single; acuminate unguis, there is no median carina on ventral face of the fourth pereiopod dactylus, the later being about 0.4 times the length of the propodus.
The new species, however, is strikingly similar to G. fimbriata Komai & Takeuchi, 1994 , from the Mid- Pacific Mountains. It differs from the latter species by the presence of a conspicuous lateral tooth or tubercle on the pleura of abdominal somites 3–5; a shorter branchiostegal spine; by pereiopods 3–5 and antennal peduncle segments somewhat different in their respective proportion when compared to similar proportions in G. fimbriata ; and the absence of curved setae on the distal part of the dactylus of pereiopods 3–5. When comparing the male pleopods 1–2 (but see below) of both species, the shape of the male endopod of first pleopod is oval in G. taludensis vs. tapering in G. fimbriata ; the relative size (proportionally shorter in G. fimbriata ) and spination of the appendix interna (a row of 20 plumose setae in G. taludensis sp. nov., vs. only 4 in G. fimbriata ) and appendix masculina (ca 40 spines in G. taludensis sp. nov., vs. ca 20 spines in G. fimbriata ) also allow to separate the two species.
In his study of the Indo-West Pacific species of Glyphocrangon, Komai (2004a: 487) also compared G. fimbriata to G. megalophthalma De Man, 1918 , reported from Indonesia, and which it closely ressembles. Among differences noted between these two species by Komai (2004a), G. megalophthalma features a conspicuous pubescence on carapace, and an antennular peduncle that clearly overeaches the anterior margin of the scaphocerite. In both G. fimbriata and G. taludensis sp. nov. pubescence is very much reduced and the antennular peduncle is shorter than the scaphocerite. In addition to this, the presence of a simple, acuminate dactylus on pereiopods 4-5 of mature females and of a bifid dactylus in males and young females of G. taludensis sp. nov. provides easy separation between the latter and both G. fimbriata and G. megalophthalma (see below).
Komai & Takeuchi (1994: 463) noted the similarity between G. fimbriata and the two species described by Faxon (1893, 1896) G. sicaria , and G. vicaria , but they separated them on roughly the same criteria as those used in the present contribution. A large specimen of G. vicaria (tl 160) was illustrated by Wicksten (1979: Fig. 4 View FIGURE 4 ) and it is clearly distinct from both G. fimbriata and G. taludensis sp. nov., and also from G. megalophthalma . The specimen of G. sicaria illustrated by Faxon (1895: Plate 34) also was a large specimen (tl 124), very distinct from G. taludensis sp. nov. As all were large, mature specimens, confusion due to examination of characters in immature specimens was avoided.
Closeness of G. fimbriata and G. taludensis sp. nov. merits attention. As related taxa (see Komai 2004a, 2006), these two species probably represent a case of vicariance for the Pacific Ocean.
All large females (cl 32.4–39.3) used herein as type material feature a simple dactylus on pereiopods 3–5, uncleft, without bifurcation ( Fig. 3F View FIGURE 3 ). The additional material examined includes 2 males (cl 19.0–21.2) and 7 smaller females (cl 21.2–29.4). These specimens and the male allotype feature a bifid dactyl on pereiopods 4– 5 ( Fig. 3I View FIGURE 3 ). A very close examination of these smaller specimens, reveals the spines and tubercles to be sharper compared to the large females used as type material, but no other significant difference could be found between the external morphology of these two series of specimens. Sex-related variation of the dactylus of pereiopods 4–5 in some Glyphocrangon species has been reported by Komai (2004a, 2004b). In mature and spawning females of G. caecescens Wood-Mason, 1891 [in Wood-Mason & Alcock, 1891a], G. rimapes , G. dimorpha Komai, 2004 , and G. unguiculata Wood-Mason & Alcock, 1891b , for instance, the dactyl is deeply cleft or bifid while it is simple and acuminate in males and young, non-spawning females (see Komai 2004a, 2004b for further details). As noted earlier, the presence in G. taludensis sp. nov. of a bifid dactyl in pereiopods 4–5 of young females and males further provides separation between the new species and both G. fimbriata and G. megalophthalma (dactyl simple in both males and non-spawning females).
Other small variations were noted. In smaller specimens the posterior sublateral and submarginal carinae of the carapace are almost obsolete. In some specimens, there are a few tubercles between the posterior first and second carinae.



| EMU |
Eastern Michigan University, T. L. Hankinson Vertebrate Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |