
Corotoca melantho, Schiodte, 1853
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4691.1.3 |
|
publication LSID |
lsid:zoobank.org:pub:30167B32-C19D-4723-9399-C9BC65522B6D |
|
DOI |
https://doi.org/10.5281/zenodo.4323778 |
|
persistent identifier |
https://treatment.plazi.org/id/216387BA-1569-FFD7-AAE1-F9BBFC38FBD8 |
|
treatment provided by |
Felipe |
|
scientific name |
Corotoca melantho |
| status |
|
Morphological notes on stenogastric individual of Corotoca melantho View in CoL : does stenogastry recapitulate phylogeny?
The post-imaginal growth in Corotocini is caused by the expansion of membranous abdomen and secondary sclerotization of sclerites. Although this fact was well known even before Seevers’s monograph on termitophilous beetles ( Seevers 1957), this phenomenon was treated only in terms of body size difference between physogastric and stenogastric individuals. That author was the first to acknowledge that the post-imaginal growth can lead to other considerable changes besides body size. Among those differences are the possible functional wings in stenogastric individuals of Termitomimus Trägårdh, 1907 , Termitozophilus mirandus Mann, 1923 and T. laetus Silvestri, 1901 (Zilberman 2019) ; and also the unusual and distinctive hind coxae and metasternum of stenogastrics Thyreoxenus major Mann, 1923 and Nasutimimus emersoni ( Seevers, 1957) .
In general, Seevers (1957) describes stenogastric individuals as frail, slender and winged forms with abdomen of typical aleocharinae appearance. Indeed, the author showed concern about the matter of identification, noting that due to several morphological discrepancies, stenogastric individuals of Thyreoxenus major and N. emersoni could even mislead someone to classify them as another subtribe than Corotocina, although perhaps as Corotocini . His notes on stenogastric forms of these two species demonstrated that some features, such as the leg thickness, are modified during post-imaginal development. In case of T. major several intermediate stages are known and suggest that the changes from stenogastric to physogastric condition are gradual. Seevers (1957) also added some remarks on Dioxeuta microps Sharp, 1899 (Termitopaediini) , and it helped us to understand the range of this transformation. Silvestri examined a series of Jacobsonella termitobia that included stenogastric and physogastric individuals; the stenogastrics were classified as “ovigera form” and physogastrics as “gestans form”. Franssen (1932) concluded that the “gestans forms” individuals belong to a different species, which he named Jacobsonella oudemansi , however, the current name of the species remains as Neodioxeuta oudemansi ( Franssen, 1932) . He observed that the apparent stages were actually males and females. Kistner (1982) describes similar changes between the different forms of Nasutimimus emersoni as noted in Seevers (1957). In this case, an intermediate stage between the stenogastric and physogastric forms was also presented. This intermediate stage appears to be present in the development of physogastric beetles, and it is illustrated for Thyreoxenus major by Emerson (1935).
The morphology discrepancies between the different forms of the same individual is probably an important factor for identification of the species. Besides, the majority of the taxonomic keys for termitophilous species, probably due to lack of material, do not consider characteristics of stenogastric individuals (Casey 1889; Seevers 1957; Fontes 1977; Caron et al. 2018; Zilberman 2019). Some characters of various stenogastric forms were included in the Seevers’ identification keys in 1957.
Seevers concerns commented above on the uncertainty of the identity of stenogastric individuals allow the raising of interesting questions about the taxonomic work on those termitophilous. Throughout the history, little attention was given to the chaetotaxy of species across the genera, but we believe that it may be the best character to identify stenogastric individuals of some Corotocini beetles. Recent taxonomic works on Corotoca highlight chaetotaxy as a great diagnose character for the group ( Zilberman 2018a; Zilberman & Casari 2018; Zilberman 2019). The fact is that whatever differences the stenogastric has in comparison to physogastric individuals, it will be limited to the matter of secondary sclerotization, while chaetotaxy remains conservative.
The stenogastric individual discussed in this paper is an example of how the chaetotaxy can help identify the specimen to a genus, even with all the other differences regarding secondary sclerotization when compared to physogastric ones. Some diagnostic features for the genus Corotoca also found in stenogastric individuals are the row of bristles on vertex, a pair of bristles on pronotum (exceptionally, C. araujoi Seevers, 1957 has no bristles on pro- notum and C. pseudomelantho Zilberman, 2018 in Zilberman & Casari (2018) has two pairs) and a comb of bristles on posterior tibia. Additionally, the eyes occupying more than 2/3 the lateral region of the head, scarcity of bristles on elytra and a V-shaped depression on pronotum, made it possible to identify the individual as a Corotoca species. Along with the long bristles on sternites V, VI and VII, and only one pair of bristles on pronotum, it is possible to conclude that the stenogastric individual is a C. melantho specimen and it is a female ( Zilberman & Casari 2018). It is noteworthy to add, that the stenogastric individual analyzed has an additional bristle of asymmetrical distribution on vertex, but it is probably an anomaly because one of the authors (Zilberman, Obs. Pers.) observed similar phenomenon in physoga stric sp ecimens of C. fontesi Zilberman, 2018 .
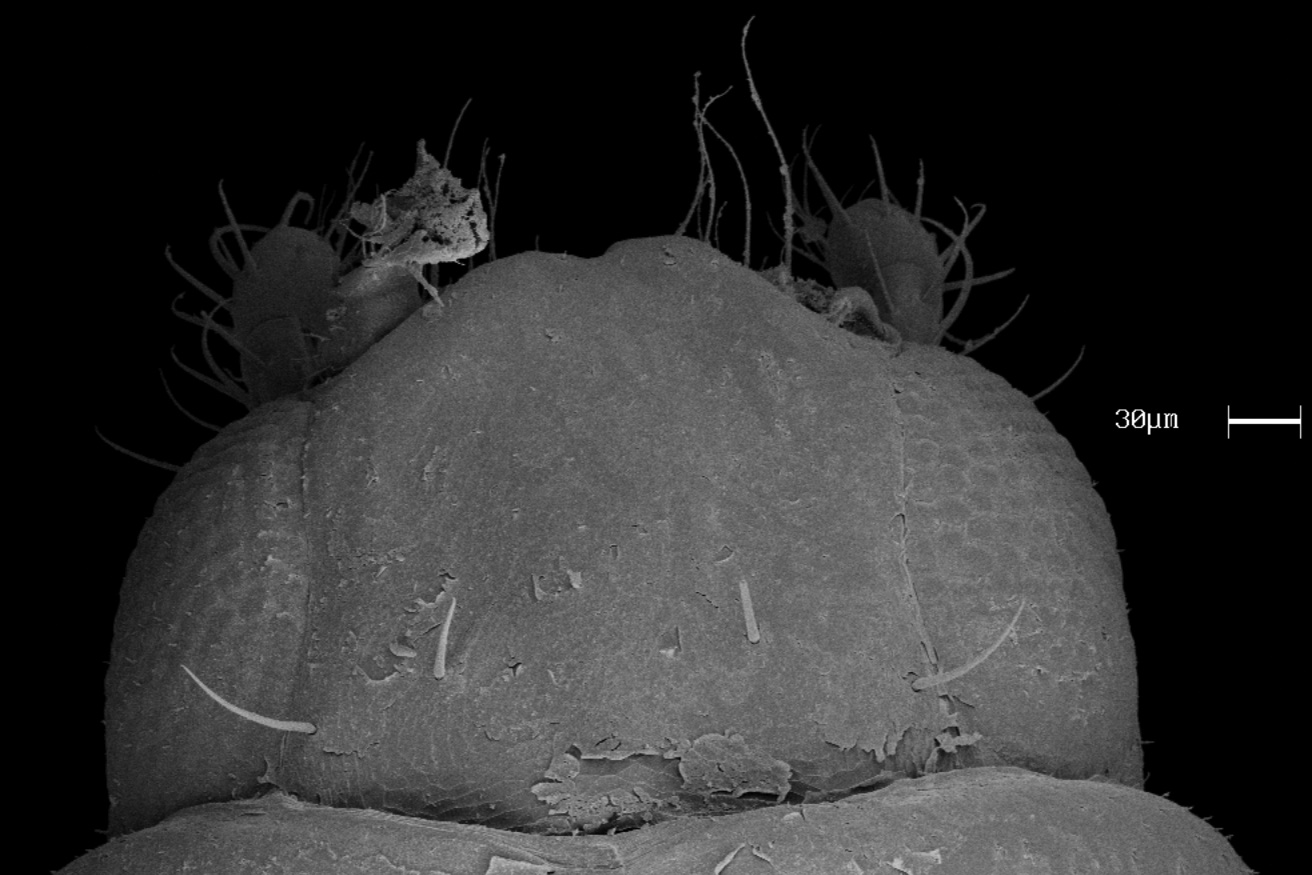
The slender, fragile and size reduced body (1.35 mm), when compared to physogastric individuals (~ 3.5 mm), and the different size of legs (being longer on stenogastric individual), fit Seevers’ description for stenogastric individuals of other species ( Figs. 1–6 View FIGURES 1–4 View FIGURES 5–6 ). Some features present in Corotoca species are cryptic on steonogastric individual, as the absence of characteristic secondary sclerotization and pigmentation. A conspicuous feature absent in stenogastric individuals is the long lateral extensions on tergites IV and V that are fused along their length and nearly encircle the abdomen.
According to Seevers (1957), the presence of wings seems to be a characteristic that also disappears throughout the development of individuals of certain species. For example N. emersoni has functional wings in stenogastric specimens and “thin-walled non-functional sacs” in physogastric ones. Furthermore, the dispersive cycle of Corotoca suggests that inside the nest the wings are of no use, since the dispersion occurs passively by immatures ( Oliveira et al. 2018). Once inside the colony (the nest), a stable environment, the dispersion and wings are no longer necessary ( Wagner & Liebherr 1992). However, the way that Corotoca specimen finds a new nest to host itself in, is still subject to discussions and uncertainties. It is expected that stenogastric individuals may provide more clues about social parasitic relationship and integration than physogastric individuals ( Seevers 1957), because Corotoca can be found inside the nest in its emergent form.
Although it is expected that all species within the genus Corotoca possess the same kind of dispersion strategy, keeping the wings throughout their life is not a character shared by all members of the group. In physogastric Corotoca , the species that have wings in that stage are C. melantho , C. guyanae Mann, 1923 , and C. pseudomelantho . In the other species, C. phylo Schiødte, 1853 , C. fontesi Zilberman, 2018 , and C. araujoi Seevers, 1957 , the wings are vestigial. Although both forms of C. melantho (steno. and physo.) are winged, the long lateral extensions on tergites IV and V that are fused along their length, possibly constrain the flexibility of the abdomen and perhaps prevents them from exposing their wings (non-functional) as the stenogastric form does.
The stenogastric form of Corotoca melantho has membranous wings that are maintained throughout the development of the individual. Whether or not physogastric wingless specimens of other species across Corotocini lose their wings during the development of physogastry and integration into the host colony is an open issue and possibly vary in different groups. Even if the adult of C. melantho does not use its wings while inside the termite nest, it is unknown if the wings can be used by the stenogastric forms to find a new nest since it is unknown how Corotoca finds a new colony. Within Corotocini , the stenogastric individuals collected by Seevers (1957) had wings (even if they did not have them as physogastric), and the same could be expected for the non-winged species in Corotoca , and perhaps this plays an important role in the search for another colony. Zilberman (2019) stated that in Termitozophilus laetus Silvestri, 1901 stenogastric individuals have wings and physogastric forms have them partially or totally removed. The author’s hypothesis is that it is reasonable to infer that stenogastric forms are the dispersal individuals and then inside the nest, they “lose their wings because they were not needed anymore”. Similar situation occurs with parasite Hippoboscidae flies, which lose their wings after locating a suitable host ( Wagner & Liebherr 1992). Another possibility is that the wings are being absent during the whole development of the species, which is also a subject to be explored across the wingless species in the genus Corotoca . Seevers (1957) was the first to show that the differences could be more than just the body size, but here it is suggested that these differences may also have a significant phylogenetic implications. These conclusions are based on a taxonomic revision and cladistic analyses performed by Zilberman (unpublished data).
The degree of secondary sclerotization may have evolved over time in certain taxa and possibly created these structures or conditions across different evolutionary branches. Corotoca melantho specimen has the long lateral extensions on tergites IV and V are caused by secondary sclerotization and this structure is a synapomorphy of the group of Corotoca species associated with C. cyphergaster living in Brazilian Cerrado and Caatinga, and Argentinean Chaco (which means all species excluding C. guyanae , this one is associated with C. cavifrons Holmgren, 1910 living in forests). The stenogastric individual of C. melantho do not have character state of this structure, which is considered plesiomorphic.
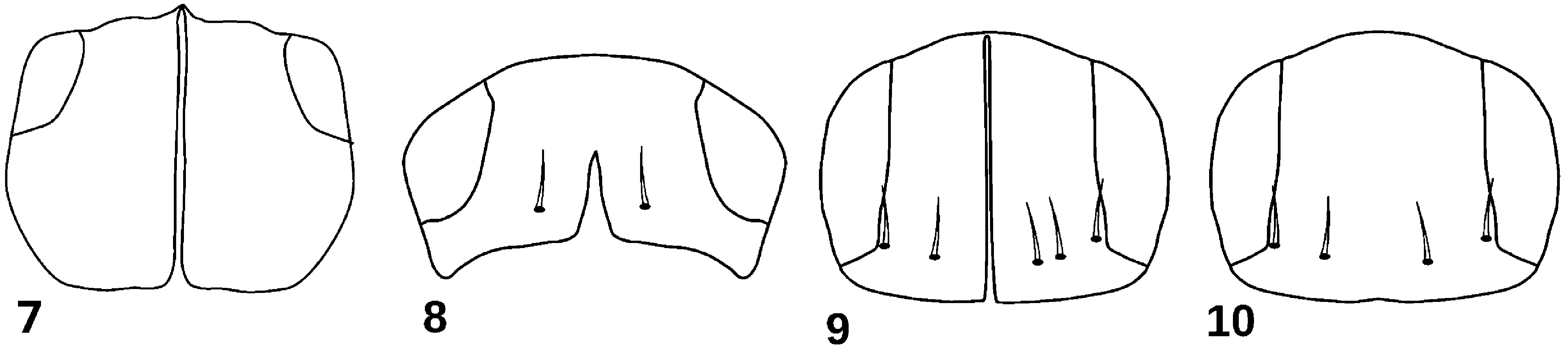
The longitudinal suture present on head’s surface of physogastric Corotoca guyanae is absent on physogastric specimens of remaining Corotoca species. The suture, however, is present in the stenogastric individual analyzed in this study. It is noteworthy that the suture occupies almost entirely the length of the head surface, which is different from the C. guyanae , in which it reaches half the length of the head ( Fig. 7–10 View FIGURES 7–10 ). The suture found is similar to that of physogastric Spirachtha eurymedusa Schiødte, 1853 , the sister group to Corotoca . It is possible that the suture was present in the common ancestor of Spirachtha + Corotoca and it was similar to that found in S. eurymedusa , being reduced over time in C. guyanae until be completely covered by secondary sclerotization in remaining species of Corotoca ( Fig. 11 View FIGURE 11 ), remaining visible only in stenogastric individuals. Therefore, we wonder if the stenogastric individuals of C. guyanae have a longer suture similar to that of stenogastric C. melantho and physogastric S. eurymedusa .
The pattern of ontogenetic changes occurs in such a way that if one would find a stenogastric beetle, and compared it to its physogastric form without knowing the true identity of the species, one could consider as dealing with two different species. The results of morphological study by Zilberman (unpublished data) on the phylogeny of the genus Corotoca , along with the morphology of the stenogastric individual of C. melantho , strongly suggest the importance of study of stenogastric specimens of different Corotocini groups. When stenogastry is taken into consideration, the evolutionary history narrative of certain characters could be changed entirely. In fact, this type of development needs to be thoroughly studied. There are probably many triggers for the first changes in stenogastric individuals. Studies on physogastry and its physiological role in termitophily are necessary for a better understanding of this transformation.
Identifying stenogastric specimens: a new or known species?
Given the discussed morphological discrepancies between stenogastric and physogastric individuals, a possible question to address is why we consider the former belonging to an already known rather than a new species. Following the collection of stenogastric individuals, there are two facts to consider to lead to correct identification and possibly it is a general rule for physogastric termitophiles: the rarity of finding stenogastrics specimens in nests and the morphological changes resulting from the secondary sclerotization process.
The shortage of stenogastric individuals in nests is evident not only from practical experience, but also from the rarity of stenogastric individuals in termitophile collections. The Museu de Zoologia da Universidade de São Paulo has an extensive collection of Corotocini , with over a thousand specimens of mainly Neotropical species but the number of stenogastric individuals is only ten (nine for Termitozophilus laetus and one C. melantho ). The difficulty in collecting stenogastric individuals means that when it occurs, it is unlikely that the physogastric of this species will remain undiscovered during nest investigation procedures. Additionally, known morphological changes resulting from the secondary sclerotization process are a rule rather than an exception, and radical differences in morphology are expected.
The first step after collecting a stenogastric sample is to determine the species of all physogastric representa- tives of all species within a termite nest; chances are this individual belongs to one of these species. Subsequently, it is essential to know that the secondary sclerotization process undermines characters that may be useful only for identifying physogastrics. It is recommended to use characters that are expected to be conservative at both stages, and here it is believed those related to chaetotaxy are continuous among the adult semaphoronts of a given species. The less the chaetotaxy is addressed in a given description the greater is the difficulty in determining the species of its stenogastric variation.
Behavioral notes
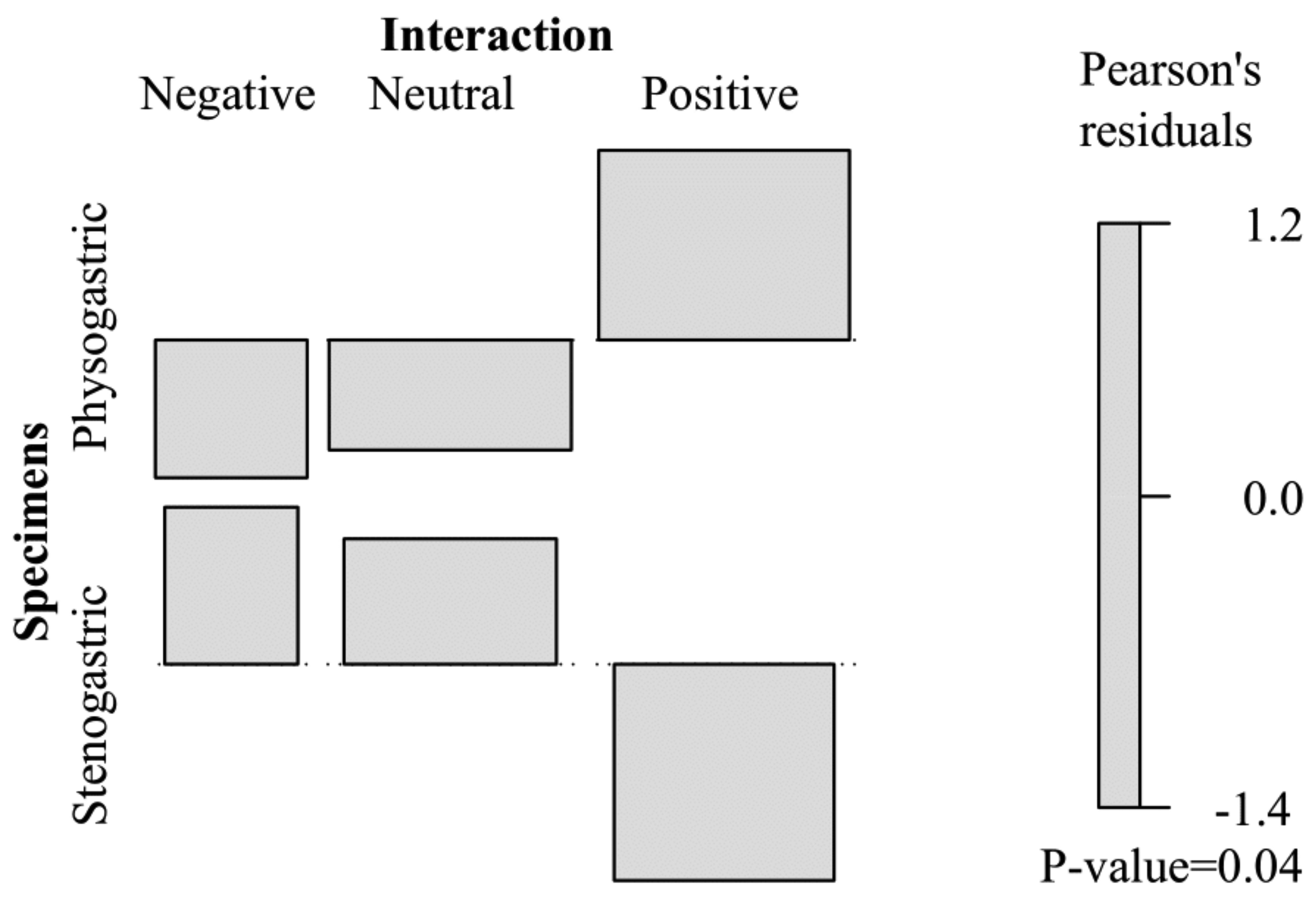
It is expected that the same species display similar behavior, but it is evident that the stenogastric form does not behave similarly to the physogastric one (χ² =6.4299, df=2, P =0.04).
The stenogastric Corotoca melantho beetle shows a particularly curious behavior: frequently displaying and pulling his wings using the hind legs. The absence of intersegmental membranes in the abdomen, coupled with the absence of secondary sclerotization that surrounds the outermost part of the body is probably responsible for this action. The bioassay revealed no statistical differences in frequency of wing pulling in the presence of termites (Wilcoxon signed rank V=20.5, P>0.05), and C. melantho does not lose its wings along lifespan, and therefore we are not sure of the reasons behind this action. Zilberman (2019) found multiple T. laetus lacking a wing or having torn wings, which suggests that wing puling might be common among Corocotini beetles. The probable reason why C. melantho exhibits this behavior, despite not losing its wings, may be simple genetic memory from wing pulling ancestors.
Interactions between termites and termitophiles take place at any moment inside the nest, which helps in understanding how these organisms integrate into the termite society. Unlikely the physogastric form, stenogastric C. melantho is much more agile among termites, moving faster, and do not interact actively with their hosts. Contact avoidance can be hypothesized as a defense strategy, since the beetle may not be fully integrated into the colony, as with some mymercophiles that avoid ant contact for days before colony invasion ( Hölldobler & Kwapich 2019). The high agility and faster movements can be explained by the absence of a greatly developed abdomen that carries a larva inside, increasing its weight and being the cause of the slowness and fragility of the beetles. Similar agility is present in species of limuloid beetles, which prefer to go effectively unnoticed within their host society ( Cai et al. 2017).
Avoiding contact with the new colony seems to be a good option when one is not fully integrated with society. In some mymercophiles the "odor" of the colony can be acquired gradually. Kistner’s (1979) observations of Mimocetes flying from one side to another for about 10 cm, on a Dorylus Fabricius, 1793 trail, and then entering thick lines of Dorylus (Anomma) suggest that it is acquiring odors. However, even if the odor of the species does not seem to be of high importance, Kistner (1979) shows that experimental transfers of termitophiles from one termite nest to another conspecific nest have always been successful, suggesting that this is not regarded as an integrating mechanism but as an adaptation to the nest as a niche.
Chemical integration provides an adequate elucidation for social interactions. Cuticular hydrocarbons in termites can mediate the nest mate recognition, and the same mechanism can be understood when treating termophiles as members of the colony since the recognition in social insects is mediated by chemical affinity ( Rosa et al. 2018).
When Corotoca pupates, their way for reintroduction in termite colony is not yet well understood. Zilberman et al. (2019) explains that the stenogastric form can enter (invade a new colony) in the nest different from birth origin, and this change of hosts can influence the adaptation of Corotoca inside the colony, as well as its interactions, since the beetle would come from a nest with different chemical profile than the current. The probable chemical profile differences between Corotoca and C. cyphergaster ( Rosa 2018) may be the reason for the stenogastric defensive behavior ( Fig. 12 View FIGURE 12 ), avoiding contact with its host, since its integration into society would occur gradually.
The presence of stenogastric individuals of Corotoca melantho within the termite nest reinforces the hypothesis raised by Zilberman et al. (2019) about the way that C. melantho invades the nest of its host. After the pupal stage, which occurs outside the nest (see Oliveira et al. (2018)), C. melantho is probably guided by trail scent (chemical cues) of its host and follows it to the corresponding nest, invading it. Only females of Corotoca spp. are encountered accompanying termites in foraging trails ( Oliveira et al. 2018; Moreira et al. 2019). The males live exclusively within the nests, leading to believe that its only purpose is mating. The life within the stable and resource-rich nest termitarium ( Kistner 1969), plus the reduced spermatheca leads to the belief that there is frequent reproductive contact between males and females of Corotoca species ( Pisno et al. 2019).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.