
Megalomma interrupta, Capa & Murray, 2009
|
publication ID |
https://doi.org/ 10.3853/j.0067-1975.61.2009.1529 |
|
persistent identifier |
https://treatment.plazi.org/id/2202BF4F-FFC5-5B74-FE9C-52F4FC3AF96F |
|
treatment provided by |
Felipe |
|
scientific name |
Megalomma interrupta |
| status |
sp. nov. |
Megalomma interrupta n.sp.
Figs 2J–L View Figure 2 , 4E–F View Figure 4 , 5B View Figure 5 , 7 View Figure 7 , 8 View Figure 8
Material examined. Queensland. HOLOTYPE AM W35478, One Tree Island , 23°30'S 152°05'E, Stn 10.4, coll. P.A. Hutchings, Oct. 1972 GoogleMaps . PARATYPES: AM W35479, (6 spec., 1 on SEM stub AM 501 ), same locality as holotype GoogleMaps ; AM W35480 (1 spec.), Stn 11.2, same locality and date GoogleMaps ; AM W35481 (1 spec.), Stn 1.1, same locality and date GoogleMaps ; AM W35482 (1 spec.), Stn 5.4, same locality and date GoogleMaps ; AM W35483 (2 spec.), Stn 14.3, same locality and date GoogleMaps ; AM W35484 (1 spec.), Stn 2.4, same locality and date GoogleMaps ; AM W35485 (2 spec.), Stn 10.2, same locality and date GoogleMaps ; AM W35486 (1 spec.), Stn 15.1, same locality and date GoogleMaps ; AM W35487 (1 spec.), Stn 5.1, same locality and date GoogleMaps ; AM W35488 (5 spec., 2 on SEM stubs MI090–91), Stn 10.3, same locality and date GoogleMaps ; AM W35489 (1 spec.), Stn 7.1, same locality and date GoogleMaps ; AM W35490 (1 spec.), off Station Beach , Lizard Island, 14°41'S 145°27'E, 75 LIZ 1–8, coll. P.B. Berents & P. Hutchings, 6 Jan. 1975, from dead Pocillopora , 3 m GoogleMaps ; AM W35491 (3 spec., one on SEM stub), south headland of Turtle Beach , Lizard Island, 14°39'S 145°27'E, 76 LIZ 21-1, coll. P. Berents & P. Hutchings, 27 Aug. 1976, dead branching coral, 3 m GoogleMaps ; AM W35492 (1 spec.) off Granite Bluff , Lizard Island, 14°39'S 145°27'E, 76 LIZ 27-1, coll. P. Berents & P. Hutchings, 31 Aug. 1976, thin plates dead coral, 6 m GoogleMaps ; AM W35493 (2 spec.), off Coconut Beach , Lizard Island, 14°41'S 145°28'27"E GoogleMaps , QLD1185 , coll. K.B.Attwood & P.A. Hutchings, 29 Mar.1995, dead coral at foot of reef, covered in Lithothamnion , 12 m ; AM W35494 (1 spec.), reef front between Bird & South Islands, Lizard Island, 14°41'53"S 145°27'51"E GoogleMaps , QLD1187 , coll. P.A. Hutchings, 30 Mar1995, dead coral, 13 m ; AM W35495 (1 spec.), off North Point Lizard Island , 14°38'51"S 145°27'12"E GoogleMaps , QLD1183 , coll. P.A. Hutchings, 28 Mar. 1995, dead coral with heavy coralline algal growth, 20 m . Western Australia . AM W35496 (2 spec.), Kendrew Island , Dampier Archipelago, 20°28'42"S 116°32'E, coll. P.A. Hutchings, 3 Apr. 1987, crevice fauna, 10 m GoogleMaps ; AM W35497 (1 spec.), 2 km west of Angel Island , Dampier Archipelago, 20°29'46"S 116°47'29"E, WA 639, coll. P.A. Hutchings & L. Avery, 4 Aug. 2000, pinnacle, dead coral substrate & large bivalves heavily encrusted and bored, 10 m GoogleMaps ; AM W35498 (3 spec., 2 on SEM stub MI089 and MI090 ), west of Angel Island , 20°29'03"S 116°47'50"E, WA619, coll. P.A. Hutchings & L. Avery, 25 Jul. 2000, dead coral, 6 m GoogleMaps ; AM W35499 (2 spec.), East Montlivet Island , Kimberley region, 15°06'S 125°18'E, Stn 50, coll. P.A. Hutchings, 16 Jul. 1988, 6 m GoogleMaps ; AM W35500 (1 spec.), south side of Long Reef , Kimberley region, 14°01'S 125°44'E, Stn 60, coll. P.A. Hutchings, 18 Jul. 1988, 20 m. Indonesia GoogleMaps . MAGNT W5975 View Materials (1spec., Bay of Maumere , Pasir Sari, 8°37'00"N 122°13'59"E, 24–27 m, rubble E, coll. B.C. Russell, 6 Nov. 1991. GoogleMaps
Additional material. Queensland (identification is tentative due to poor preservation): AM W198006 (2 specimens, 1 on SEM stub MI086 ), Calliope River, Gladstone , 23°51'S 151°10'E, coarse sand, 3.9 m, coll. P. Saenger, Aug. 1982 GoogleMaps ; AM W198007 (1 spec.), same locality and date, silty sand, 5.6 m GoogleMaps .
Diagnosis. This species is characterized by a combination of features: radiolar eyes present in dorsalmost and lateral radioles only, with approximately four radioles in between without eyes; radiolar skeleton with approximately 10 cells in cross section; collar margins not fused to the faecal groove, shallow inconspicuous pockets present, caruncle absent, and inferior thoracic broadly hooded chaetae with distal end narrowing abruptly (type A).
Description. Holotype 22 mm long and 2.5 mm wide with body flattened dorsoventrally, thorax wider than abdomen and abdominal segments diminishing in size posteriorly. Eight thoracic and 78 abdominal chaetigers. Crown longer than thorax, with 15 radioles in each lobe semicircular lobe. External margin of radioles quadrangular at the base ( Figs 2I View Figure 2 , 7C View Figure 7 ) and rounded towards the tips, without lateral flanges but with two longitudinal bands of cilia. Tips of radioles tapering, shorter than pinnules ( Fig. 2J View Figure 2 ). Basal membrane absent. Radiolar skeleton with about 10 cells in transverse section at the base of radioles ( Fig. 5C View Figure 5 ). Dorsalmost pair of radioles longer than the rest, each with a large subdistal compound eye almost surrounding the whole radiole, and with visible ommatidia ( Figs 2J View Figure 2 , 7K View Figure 7 ), adjacent next five pairs of radioles without eyes, next two pairs of lateral radioles with small, similarly-sized subdistal eyes. Dorsal lips with radiolar appendages as long as two thoracic chaetigers, with lateral lamellae almost reaching the tip of appendage; three dorsal pinnular appendages each. Caruncle absent. Ventral lips rounded and well developed. Ventral sacs and parallel lamellae present ( Fig. 7A View Figure 7 ). Collar with pointed ventral lappets ( Figs 2I View Figure 2 , 7A View Figure 7 ); lateral margins of collar smooth with dorsolateral U-shaped indentations on both sides forming inconspicuous pockets and continuing as ridges to the middorsal faecal groove ( Figs 2I View Figure 2 , 7B–C View Figure 7 ). Ventral shields quadrangular with parallel lateral margins, not in contact with the neuropodial tori ( Figs 2K View Figure 2 , 7A View Figure 7 ). First ventral shield longer than the rest, with a midanterior incision (m-shaped anterior margin). First chaetiger with superior and inferior elongate narrowly hooded notochaetae; superior row longer than the inferior one ( Fig. 7D View Figure 7 ). Rest of thoracic chaetigers with superior elongate narrowly hooded and inferior broadly hooded notochaetae (about 20, type A) arranged in two rows ( Fig. 7H View Figure 7 ). Notopodia of thoracic chaetigers with elongated lobe between superior and inferior chaetae ( Fig. 7B View Figure 7 ). Neuropodial tori becoming slightly shorter more posteriorly. Thoracic uncini with several rows of small teeth, all similar in size above main fang ( Fig. 7F,G View Figure 7 ); uncinus with well developed breast and handle about twice the length of the distance from breast to main fang ( Fig. 4E View Figure 4 ). Companion chaetae with asymmetrical membrane covered proximally with rows of small teeth ( Fig. 7E View Figure 7 ). Abdominal neuropodia with elongate, broadly hooded chaetae arranged in two rows ( Fig. 7I View Figure 7 ). Notopodial abdominal uncini similar to the thoracic ( Fig. 7J View Figure 7 ) but with shorter handle ( Fig. 4F View Figure 4 ). Pygidium as a rounded papilla without pygidial eyes.
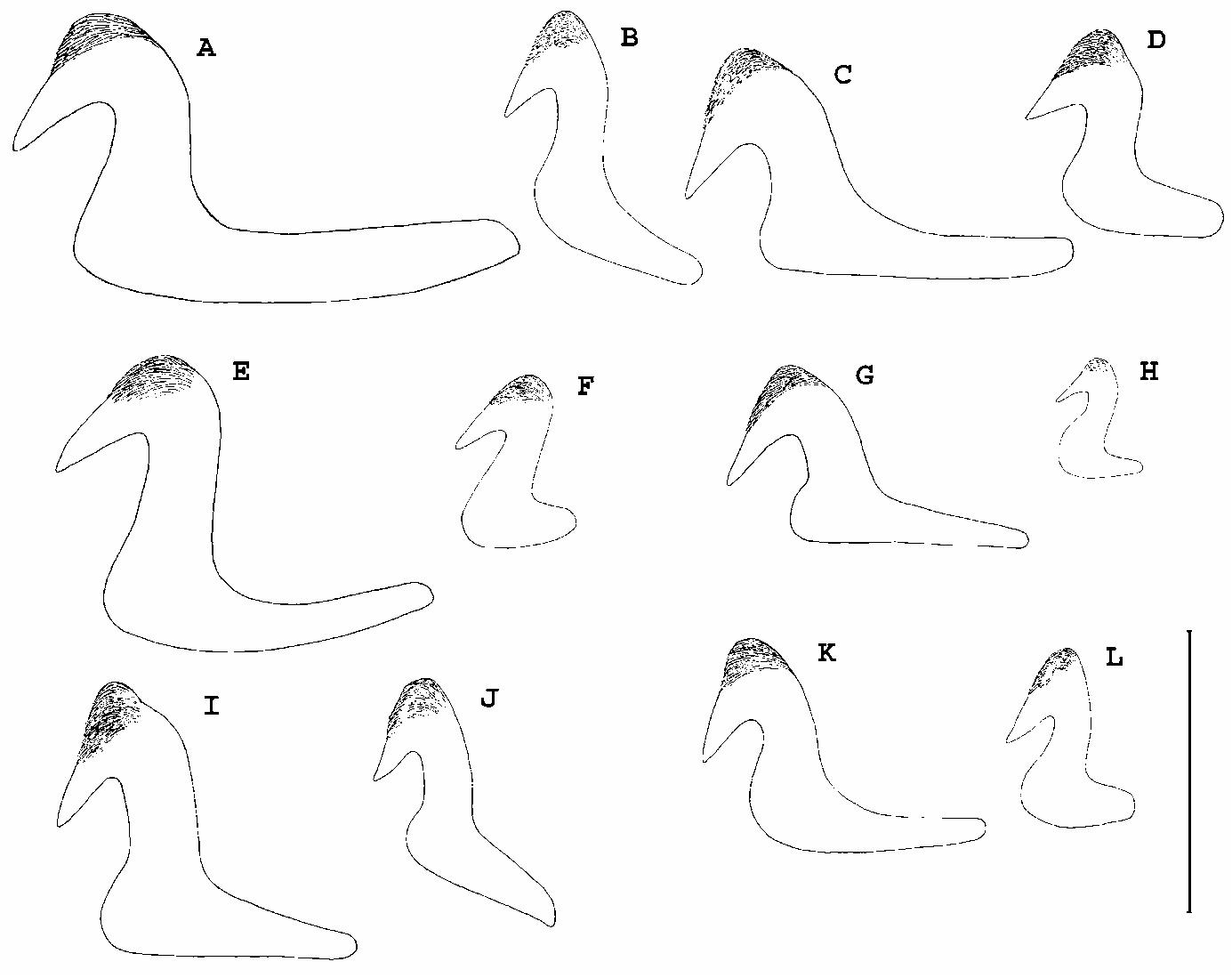
Variation. There is some variability in the number of thoracic chaetigers, with some paratypes having nine or ten. The number of radioles varies between 10 and 18 pairs and phyllisae n.sp.; (B) M. interrupta n.sp.; (C) M. cf. miyukiae ; (D) M. inflata n.sp.; (E)
Megalomma sp. 1 ; (F) M. quadrioculatum ; (G) M. trioculatum ; (H) M. lanigera ; (I)
M. suspiciens . Scale: 0.1 mm.
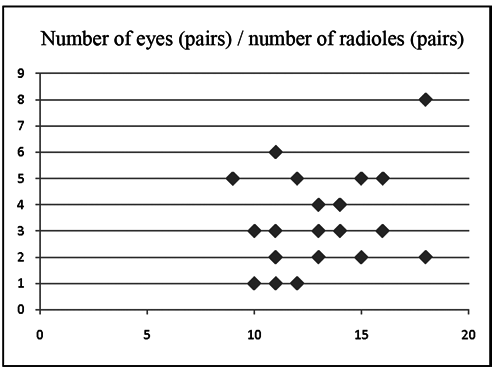
the number of compound subdistal eyes also differs among specimens and is not related to the number of radioles ( Table 1, Fig. 9 View Figure 9 ). All specimens share the same arrangement of a large radiolar eye on each of the dorsalmost radioles, but the small, similarly-sized subdistal radiolar eyes on a few lateral radioles can be absent entirely from some specimens. Dorsal lips bear long radiolar appendages that in some specimens reach the length of four thoracic chaetigers. In all specimens the peristomial collar has dorsal margins separated from the faecal groove, but in some specimens under the dissecting microscope, it is not clear whether pockets are absent or present, as some specimens exhibit low membranes as continuations of the dorsal margins of the collar which could be considered as vestigial pockets (according to Knight- Jones, 1997) (see Fig. 2M View Figure 2 ), a feature that is made very clear using scanning electron microscopy (see Fig. 7B,C View Figure 7 ). Also, the ventral lappets of the collar may be observed as overlapping or not, depending on whether they are erect or folded back. Specimens show variability in the size of the gap between ventral shields and thoracic tori, even within the same sample. In some specimens (e.g., holotype, paratypes, and AM W35498) shields and tori are separated by a wide gap, in others (e.g., AM W35496) they are separated only in posterior chaetigers, while in other specimens they are in contact in all chaetigers (e.g., AM W35497).
After observing the variability within this morphospecies under both dissecting and compound microscopes, some specimens were selected from several different localities covering the species’ distribution and which also displayed the variability in the distribution of radiolar eyes, and these were studied under SEM. Thoracic and abdominal chaetae appear to be similar in shape, size and arrangement within their fascicles ( Fig. 8B,C,F,G,J,M,N View Figure 8 ) but variability was observed among thoracic and abdominal uncini ( Fig. 8A,D,E,H,I,K,L,O View Figure 8 ). All specimens have similarly-sized teeth covering half the surface of the main fang in thoracic uncini, and a slightly greater surface area of abdominal uncini, but some specimens show relatively larger teeth ( Fig. 7A,D,E,H,I,K View Figure 7 ) than others ( Fig. 8L,O View Figure 8 ); however these differences do not seem to be correlated with the specimens’ differing geographical localities, as variability is observed among specimens from the same sample. Until further studies reveal if this is, in fact, a complex of species that coexist in the same habitats in the same localities, we consider this variability to be intraspecific.
Colour pattern. The intensity of the pigmentation varies among the specimens and could be due to different methods or lengths of time of preservation. The specimens from Western Australia are more pigmented in general that those from Queensland ( Figs 2J–M View Figure 2 ). The thorax is anteriorly pigmented, particularly dorsally and ventrally around tori. The base of the branchial crown is also pigmented, there is a longitudinal medial line basally on each radiole, and some specimens also exhibit irregular, variable-width, transverse bands of pigment on the crown.
Etymology. The name of this species refers to the discontinuous arrangement of eyes on the radioles.
Remarks. These specimens do not fit into the classification proposed by Knight-Jones (1997) as her categories do not consider the combination of the following features: dorsal margins of collar not fused to the faecal groove, pockets present and compound eyes in dorsalmost and also in some lateral radioles. These characters, common to all these specimens, justify the inclusion of a new category (called Group 2E herein) in Knight-Jones’ classification. The collar pockets could be considered as vestigial or even absent in some specimens and therefore this species should also be compared with those previously placed in the categories 2B, 2C and 2D which also lack dorsolateral collar pockets. The new species resembles M. messapicum Giangrande & Licciano, 2008 , from the Mediterranean, in the particular arrangement of compound subdistal radiolar eyes, with a large eye almost surrounding the tip of the dorsalmost radioles, as well as some lateral radioles bearing small, similarly-sized subdistal radiolar eyes, and with those radioles between the dorsalmost and lateral ones lacking eyes (see Table 1). These two species, confined to distant and isolated geographic areas, are distinguished by the difference in the degree of fusion of collar margins with the faecal groove, separated in M. interrupta n.sp. and fused in M. messapicum ; moreover M. interrupta n.sp. is characterized by the presence of large interramal lobes in thoracic notopodia and a radiolar skeleton with around 10 cells in transverse section, whereas M. messapicum does not possess enlarged interramal notopodial lobes, and has a radiolar skeleton of four cells in cross-section (Giangrande, pers. comm.).




| AM |
Australian Museum |
| MAGNT |
Museum and Art Gallery of the Northern Territory |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |