
Cucurbita, 1973
|
publication ID |
https://doi.org/ 10.1016/j.phytochem.2022.113110 |
|
DOI |
https://doi.org/10.5281/zenodo.8257224 |
|
persistent identifier |
https://treatment.plazi.org/id/220DD551-FFAC-FF80-E950-EBB0FBC685AB |
|
treatment provided by |
Felipe |
|
scientific name |
Cucurbita |
| status |
|
2.2. Pigment synthesis in Cucurbita seedlings
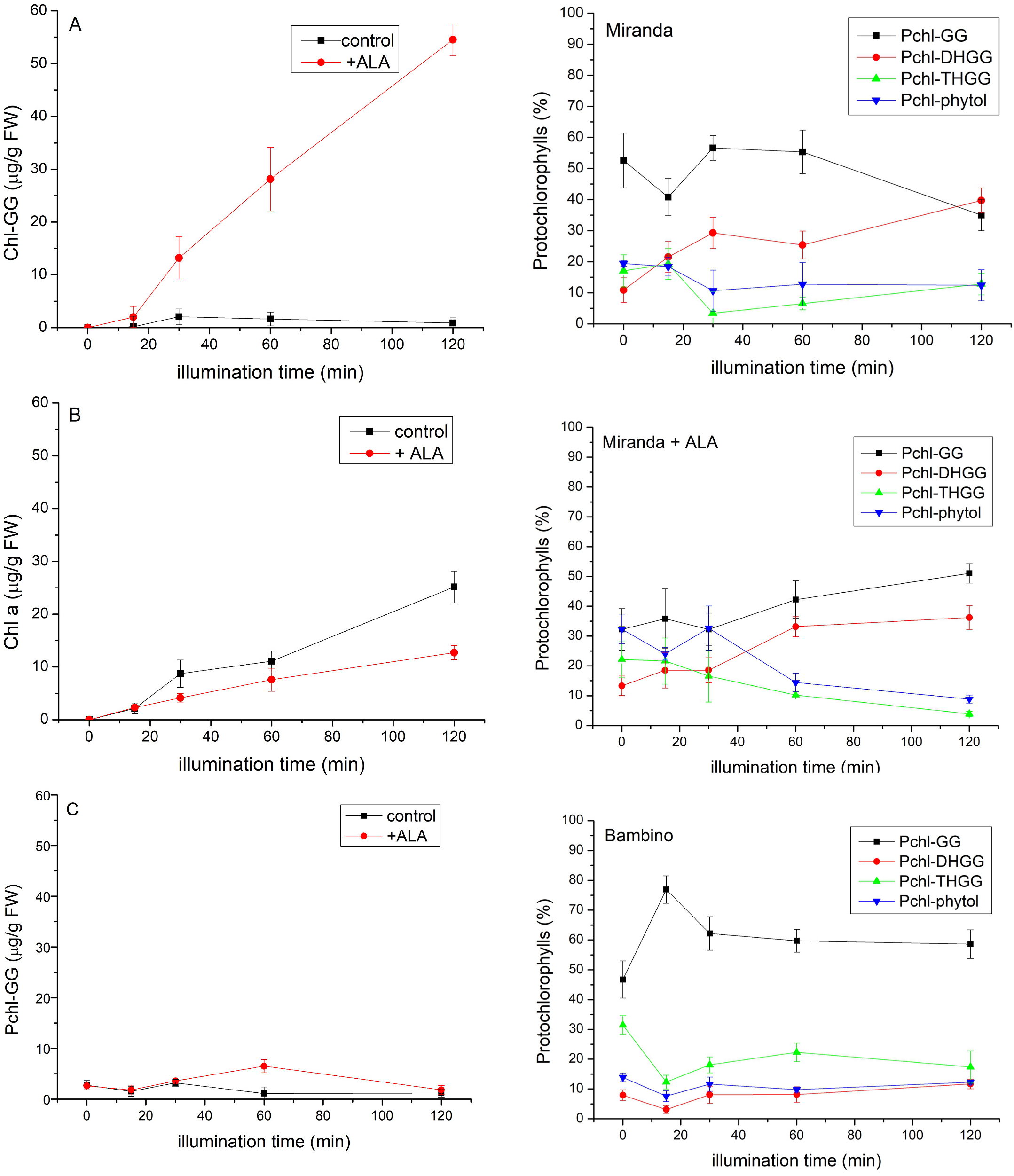
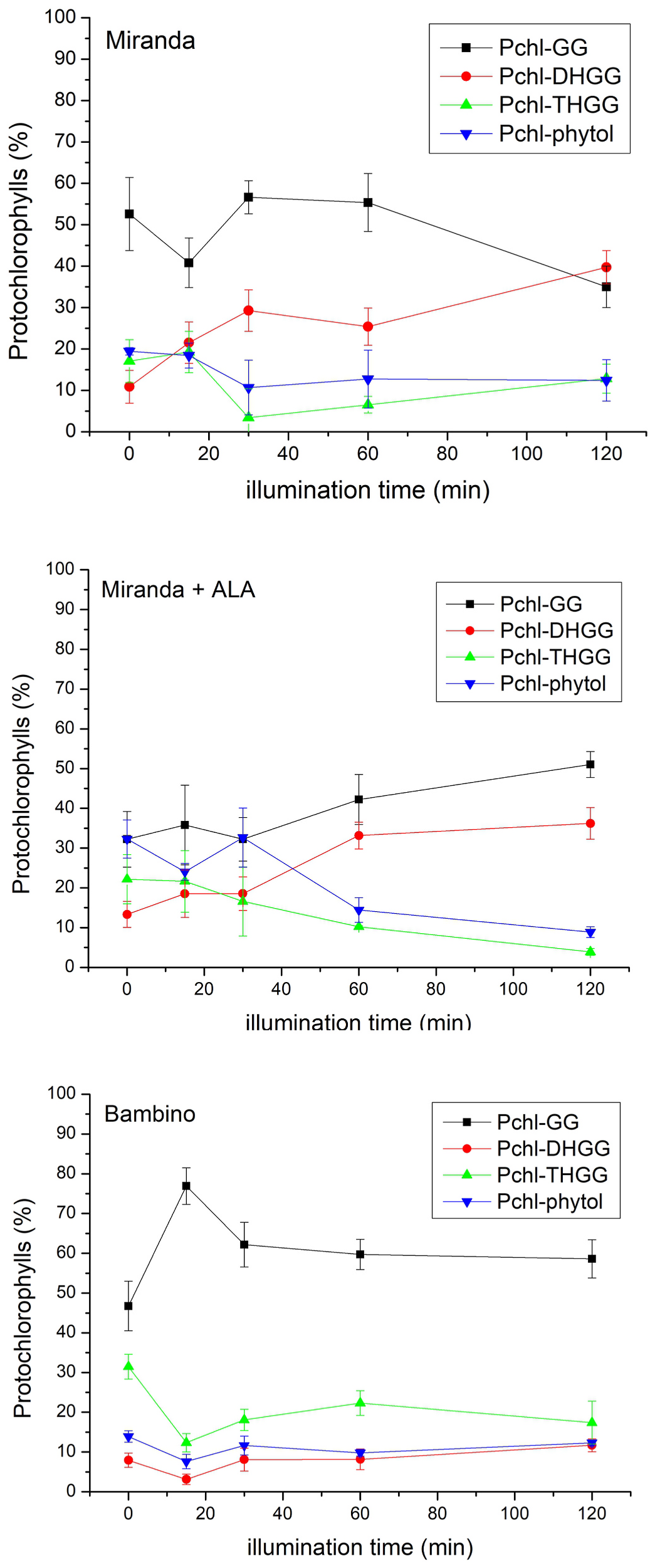
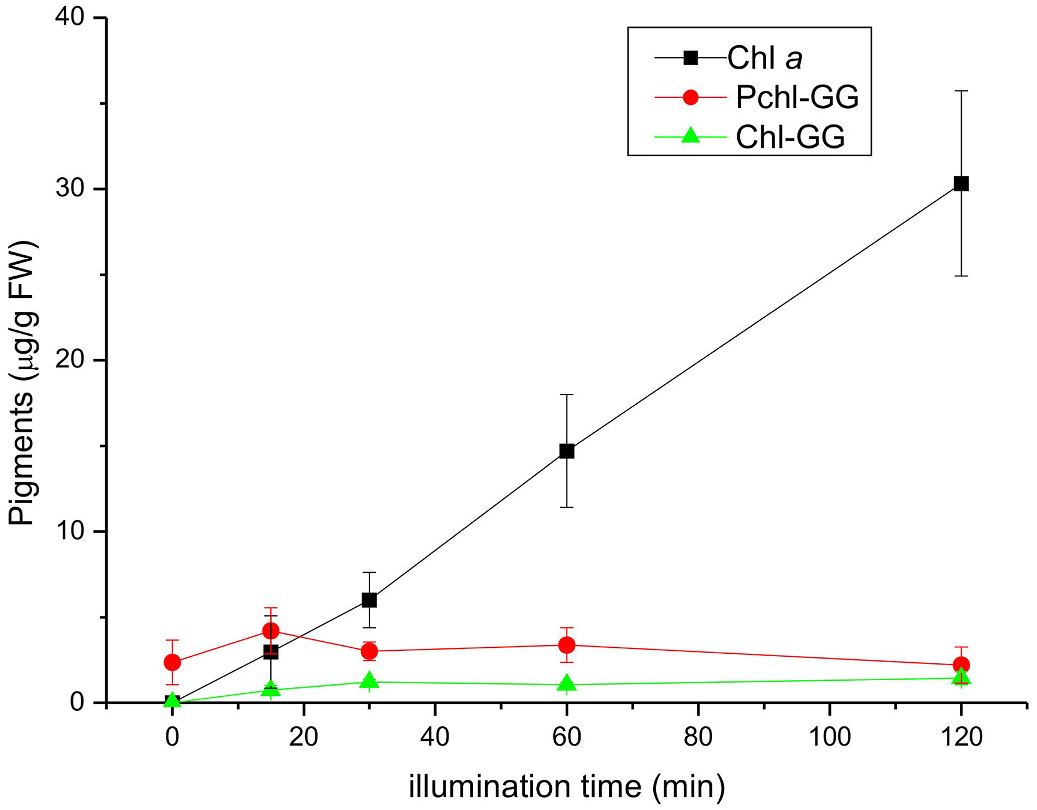
An analysis of porphyrin pigments in etiolated C. pepo ‘Miranda’ seedlings revealed that when subjected to continuous illumination ( Fig. 4 View Fig ), Chl a content in the control plants increased gradually, as expected, while the level of Chl-GG, a precursor of Chl a, was low and did not change significantly during illumination, like that of Pchl-GG. However, when the seedlings were incubated with ALA (precursor of the porphyrin ring) before illumination, the Chl-GG content increased dramatically with illumination time, while the synthesis of Chl a was inhibited. The synthesis of Pchl-GG was not affected very much ( Fig. 4 View Fig ). It is supposed that Chl a is synthesized by Chl synthase from Chlide a and both phytyl-PP or GG-PP, depending on the plant species, light conditions (darkness or light), the age of the plants and other factors ( Schoefs et al., 1998; Schoefs and Bertrand 2000, Schmid et al., 2001 and 2002). n.d. – not determined. These data suggest that Chl a is not synthesized from GG-PP via Chl-GG but rather directly from phytyl-PP in C. pepo seedlings. This conclusion is supported by the lack of other Chl a precursors (Chl-DHGG, Chl-THGG) which are intermediates in the pathway from Chl-GG to Chl a (data not shown). The observation that the Chl a level did not increase after ALA feeding suggests that the phytyl-PP supply is limited. An analysis of the composition of Pchls in primary leaves during illumination ( Fig. 5 View Fig ) shows that Pchl-GG in the C. pepo ‘Miranda’ control plants was the dominating species and the content of individual compounds did not change very much during illumination, apart from Pchl-DHGG whose contribution increased fourfold. In the presence of ALA, a gradual, but not very pronounced, increase in Pchl-GG and Pchl-DHGG levels occurred at the expense of the two other Pchls. These data suggest that the synthesis of Pchls in the leaves of developing seedlings is a relatively slow process, but not limited by the supply of substrates, such as Pchlide ( Fig. 4 View Fig ). It is also probable that the enzyme is only active at the very early stages of the seedlings’ development and its expression decreases quickly with the age of the seedlings.
When an analogous experiment was performed with a different species, C. maxima ‘Bambino’, the pigment content changes were similar to those for C. pepo ‘Miranda’ in the control plants, i.e., Chl a content increased gradually, while that of Chl-GG and Pchl-GG did not change significantly during illumination ( Fig. 6 View Fig ). The content of individual Pchls was dominated by Pchl-GG and did not change very much during illumination ( Fig. 5 View Fig ).
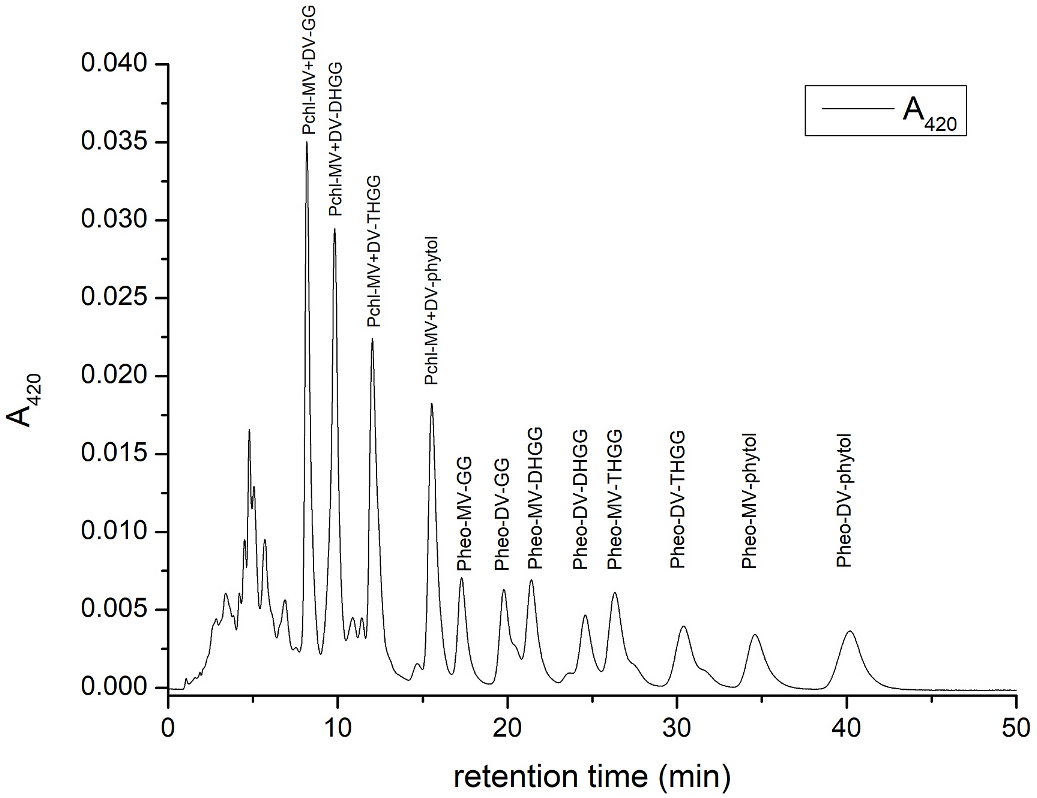
Next, we tried to follow Pchl synthase activity using different preparations, namely homogenates of seed coats, the inner seed coats of C. maxima ‘Bambino’, prolammelar bodies (PLB) isolated from etiolated leaves of C. maxima ‘Bambino’ and homogenates of etiolated leaves of C. pepo ‘Miranda’. Different isolation buffers and additional preparation treatments, such as repeated freeze-thawing, were also used. We applied a broad substrate concentration range of both Pchlide and side-chain precursors (GG-PP and phytol-PP) and incubation periods (see Experimental section). However, in none of the preparations was Pchl synthase activity found. This suggests the latency of the synthase and/or that the reaction rate of the enzyme is slow, like what was demonstrated in the case of Chl synthase ( Lindsten et al., 1990). It is also possible that in seeds the enzyme is active only at the early stages of seed development which were not analyzed in our case. Nevertheless, the homogenates of etiolated leaves of C. pepo ‘Miranda’ were active in Chl-GG and Chl-phytol synthesis, using Chlide and appropriate side-chain substrates ( Fig. S3 View Fig ), indicating that the assay method, at least for the chlorophyll synthase activity, is appropriate. Both Chl-GG and Chl-phytol were found as double peaks, showing identical absorption maxima to those of Chl a, and result probably from the presence of both cis/trans isomers of the side-chain substrates.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.