
Scalacurvichthys naishi, Cawley & Kriwet, 2018
|
publication ID |
https://doi.org/ 10.1080/14772019.2017.1330772 |
|
publication LSID |
lsid:zoobank.org:pub:04383E2A-551A-4F57-8996-68E06EFA52E0 |
|
DOI |
https://doi.org/10.5281/zenodo.10903612 |
|
persistent identifier |
https://treatment.plazi.org/id/23236B4B-1F3B-FFBB-FEBA-FDF95802FC94 |
|
treatment provided by |
Felipe |
|
scientific name |
Scalacurvichthys naishi |
| status |
sp. nov. |
Scalacurvichthys naishi sp. nov.
( Figs 1–5 View Figure 1 View Figure 2 View Figure 3 View Figure 4 View Figure 5 )
Type specimen. Holotype SMNK-PAL. 8613. Complete specimen.
Age. Bet Meir or the slightly younger Amminadava Formation, middle part of the Judea Group, early to middle Cenomanian, early Late Cretaceous.
Type locality. Limestone quarry near the village of Beit El, Binyamin Region, West Bank, Israel.
Geographical distribution. Only known from the type locality.
Stratigraphical range. Early–middle Cenomanian, Late Cretaceous.
Diagnosis. As for the genus. Only known species.
Derivation of name. The name of the new species is dedicated to Dr Darren Naish who is currently writing a book on the entire vertebrate fossil record and is prolific in publishing research on dinosaurs, pterosaurs and marine reptiles amongst many other groups of tetrapods.
Description.
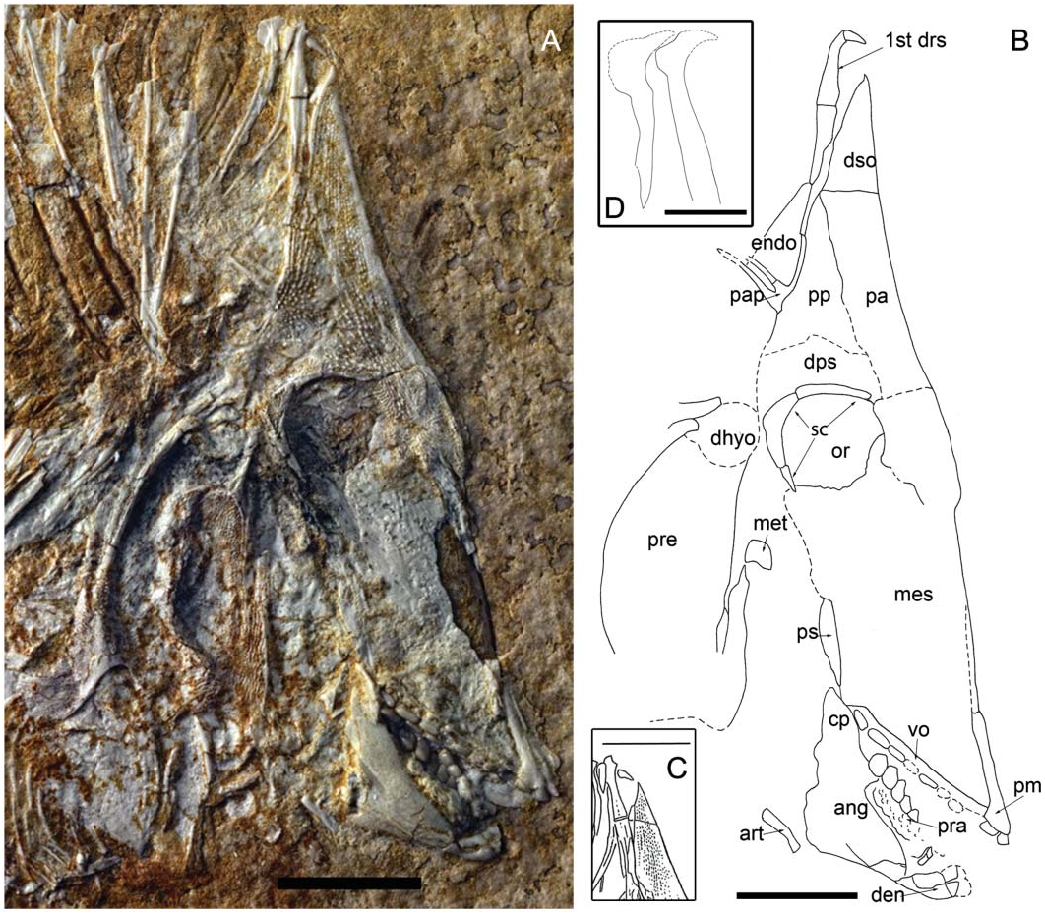
Size and general morphology. This new taxon is a small-sized pycnodont ( Fig. 1 View Figure 1 ). The SL of the new taxon is 12.3 cm. Its general body shape is oval with a deeply sloped skull. The snout is very deeply turned down. At a maximum body height of 6 cm it is relatively high bodied but distinctly less high than other pycnodonts such as Gyrodus from the Late Jurassic. The highest point of the body is at the first dorsal ridge scale which would have protruded 6 mm above the dermosupraoccipital when the animal was still alive. This dorsal ridge scale is spine-like and elongate, and curves anteriorly at the tip.
The dermosupraoccipital is triangular in form and is directly dorsal to the parietal bone, the only bone in the skull roof that it contacts. The dermocranial fenestra is absent from this specimen. The ventral apex appears to be at the posterior edge of the post-coelomic bone just before the anal fin. Conversely, the dorsal apex appears to be absent. The flank scales are reduced to scale bars, with dorsal flank scales covering only the abdomen while ventral flank scales cover both the abdomen and trunk. Scales are absent from the body posterior to the post-coelomic bone. The cleithrum appears to be a large bone that encircles the opercular skeleton, but the bone mixing with the rocky matrix makes its true form difficult to discern. Other than three fragmented hypochordals in the caudal fin and neural spines in the caudal peduncle, all of the body posterior to the post-coelomic bone is preserved as an imprint in the rock only. The dorsal and anal fins are extremely sickle shaped with no evidence of either fin contacting the caudal fin. The body tapers posteriorly to the rather thin and un-keeled caudal peduncle. The caudal fin has a height of 4 cm and has a convex margin at the distal border. There is no evidence of pectoral or pelvic fin preservation in this specimen.
Skull roof. The boundary between cranial roof bones is difficult to discern due to taphonomic processes that are characteristic for fossil fishes from ‘Ein Yabrud. Additionally, several bones seemingly were lost post-mortem.
The parietal ( Figs 1 View Figure 1 , 2 View Figure 2 ) is a large wedge-shaped bone ventral to the dermosupraoccipital and anterior to the post-parietal and dermopterosphenotic. The dorsal margin of the parietal is flat. A faint suture in front of the orbit seems to indicate the ventral margin of the parietal, which is dorsal to the mesethmoid. The posterior margin of the bone slopes to the right in an antero-ventral direction and expands posteriorly over the dorsal margin of the orbit. This bone is heavily ornamented with tubercles. Tubercles on the anterior border are highly organized into tightly packed rows, in contrast to the widely spaced tubercles found elsewhere on the bone. On the ventral portion of the parietal where tubercles are absent, the surface is dotted with pits.
The post-parietal ( Figs 1 View Figure 1 , 2 View Figure 2 ) is situated ventral to the first dorsal ridge scale and posterior to the parietal bone. The suture between the parietal and the post-parietal is uneven. Tubercles are present all over the bone, which form into ridges at the postero-dorsal border of the post-parietal. A parietal process protrudes from its posterior border. Two long, thin branches jut out facing in a posterior direction. The distal ends of the branches are obscured by rocky matrix, which may also obscure additional branches of the parietal process.
The ventral boundary of the post-parietal is hard to observe but is assumed to occur where the bony surface becomes smoother. We interpret this bone to be the dermopterosphenotic due to the change in ornamentation and its location, being ventral to the post-parietal and dorsal to the posterior margins of the orbit.
The dermosupraoccipital ( Figs 1 View Figure 1 , 2 View Figure 2 ) is a tall and broadly triangular bone placed on top of the skull roof. It is dorsal to the parietal. The tip of the bone is pointed and has a slight convex expansion on the right side and concave margin on the left. Tubercles cover the anterior portion of the dermosupraoccipital. Posterior to the dermosupraoccipital is the first dorsal ridge scale which is incorporated into the skull roof as seen in all known pycnodonts ( Nursall 1996b; Poyato-Ariza & Wenz 2002) with the exception of Gladiopycnodontidae ( Taverne & Capasso 2013) .
Orbit and neurocranium. Bones surrounding the orbit are fragmented and only the dorsal and posterior portions are preserved. The dorsal sclerotic ring fragments have rounded anterior margins and flat dorsal margins. Sclerotic rings widen in a posterior direction. The posterior sclerotic ring is crescent shaped and dotted with pits on its surface. The antero-ventral corner of the posterior sclerotic ring ends in a distal point, and has wavy posterior margins and flat anterior margins.
Inside the orbit, an elongate, curved bone with rounded distal ends is present in the ventral half of the orbit ( Figs 1 View Figure 1 , 2 View Figure 2 ). In the postero-dorsal corner of the orbit, rocky matrix obscures the remaining portions of this bone. The bone widens near the rocky matrix. The shape and position of the bone indicates a posterior orbital bone from the left-hand side of the fish.
In the dorsal half, a large bone gently curves down in an antero-ventral direction to a point. From the point, the bone has a concave margin until the fragmentation obscures the true nature of the margin. The identity of the bone is unknown but it may possibly represent the orbitosphenoid. This bone is not heavily ossified and has numerous fractures along its ventral margin but is otherwise unornamented. The bone overlying the probable orbitosphenoid has a concave posterior margin and a lateral medial bony process slightly raised from the surrounding bone. The anterior margin of the bone is convex. The bone is heavily intermixed with rocky matrix so its identification is difficult. No infraorbitals are preserved.
A smooth, unornamented bone appears to be overlain by the limbs of the post-parietal process. Due to the anteriorly extreme shortening of the skull roof in this specimen, it is probable that this could be the posterior margin of the endocranium. However, due to the intermixing of this probable endocranial portion with the rocky matrix, it is difficult to identify the shape and true identity of this bone. Doing preparatory work to remove this sediment could be problematic due to the fact that the post-parietal process limbs overlie this bone and there is a high chance they could be damaged. The presence/absence of a postcephalic lacuna thus cannot be confirmed. Exposing the specimen to UV light, however, reveals smooth bony remains dorsal to the post-parietal process and posterior to the dermatocranium ( Fig. 3 View Figure 3 , arrow). The morphology and position of these remains correlate well with the posteriorly exposed endocranium in pycnodontids.
Jaw bones and dentition. The premaxilla ( Figs 1 View Figure 1 , 2 View Figure 2 ) has a long, thin process protruding dorsally from the toothbearing portion. The true length of this process is unclear due to fragmentation in front of the skull. Two incisiform teeth are present and have a flat base with a pointed anterior margin and rounded posterior margin. The maxilla is not preserved in this specimen.
The dentalosplenial ( Figs 1 View Figure 1 , 2 View Figure 2 ) is a small jaw bone with a broad anterior margin that narrows in an antero-posterior direction. Two teeth are present on the dentalosplenial. The incisiform teeth have a flat base with a rounded posterior margin. The posterior margin is expanded in the first tooth. On the second tooth, the anterior margin narrows in a dorso-ventral direction.
Two rows of teeth are preserved in the vomer in the roof of the mouth ( Figs 1 View Figure 1 , 2 View Figure 2 ). The left lateral row shows six teeth preserved. There is a hollow between the second and fourth tooth, which indicates a tooth was present there while the fish was alive. This brings the total of teeth in the left lateral row to seven. The shape of these teeth is an elongate oval with a smooth surface, increasing in size in an antero-posterior direction. The median row has six teeth preserved. Teeth start out as elongate ovals but become much broader the farther back in the jaw they go. A shallow hollow is present medially in all median-row teeth with the exception of the first one, which is fully flattened. Crenulated ridges are only present around the outer margin of the final tooth of the main middle row; all other teeth are smooth.
There are three rows of teeth present in the prearticular bone ( Figs 1 View Figure 1 , 2 View Figure 2 ). The right lateral row has four teeth preserved, which are globular with a rounded contour and smooth surface. Seven teeth are preserved in the principal median row. Fully preserved teeth are elongate with an oval contour and smooth surface. Rocky matrix has formed over parts of the teeth as well as uplifted sections of the right lateral row. There are only three teeth preserved on the left lateral row, which are mostly embedded in rocky matrix. Teeth that are preserved show a smooth, flattened, elongate oval shape similar to those of the principal row.
The angular is a large bone, which makes up the bulk of the lower jaw ( Figs 1 View Figure 1 , 2 View Figure 2 ). It is positioned posterior to the dentalosplenial, ventral to the prearticular bone and anterior to the articular bone. The coronoid process has a rounded margin, which is posterior to the vomer. Ridges are present in the postero-dorsal region of the angular.
The articular bone is a small, thin and elongate bone located at the postero-ventral corner of the angular ( Figs 1 View Figure 1 , 2 View Figure 2 ). The dorsal portion of the articular has a rounded margin and faces in a cephalo-caudal direction. The ventral margin is flat and located closer to the ventral border of the angular. The articular is broad at the ventral margin and narrows in a cephalo-caudal direction. It is a smooth bone without ornamentations.
The mesethmoid is a large bone, but mostly preserved only as imprint so that possible ornamentation patterns remain unknown ( Figs 1 View Figure 1 , 2 View Figure 2 ). This bone seemingly takes up most of the skull. It is ventral to the orbit and parietal and dorsal to the vomer. The posterior border is ventral to the posterior portion of the orbit and dorsal to the coronoid process of the angular.
The parasphenoid is overlain by the mesethmoid and can be observed only partially at the postero-ventral corner of the mesethmoid. The observed portion of this bone has a posterior convex margin and a smooth surface.
The bones of the palatoquadrate arcade are incompletely preserved. The metapterygoid is a small bone with a flat base, convex anterior margins and a flat posterior margin. It has a smooth surface with no ornamentation. This bone is located postero-ventral to the orbit and posterior to the mesethmoid.
Opercular series. Much of the opercular skeleton is heavily crushed and sutures cannot be seen so the extent of the bone size and its shape remain unknown at present.
Even though much of the opercular skeleton is fragmented or eroded, we interpret its structure thus: the preoperculum is a large, deep, broad bone taking up much of the antero-ventral portion of the opercular skeleton. It is located postero-dorsal to the metapterygoid ( Figs 1 View Figure 1 , 2 View Figure 2 ) and dorsal to the singularly preserved branchiostegal ray. The preoperculum is heavily ornamented with ridges occurring in a criss-cross pattern.
A large process protrudes c. 4 mm antero-dorsally from the preoperculum and is also ornamented with ridges. The distal margin of the process is flat and a posterior lateral process protrudes from the distal tip. This process has a flat dorsal margin and mixes with rocky matrix in its ventral region so the shape of the ventral margin remains unknown. The process is found postero-ventral to the posterior plate of the orbit.
Between the dorsal preopercular process and the antero-dorsal extension of the cleithrum lies the dermo-hyomandibular. The dermohyomandibular is located postero-dorsal to the metapterygoid. This is a smooth bone with a convex dorsal border. A small gap appears between the ventral border of the dermohyomandibular and the dorsal border of the preoperculum. This suggests that the preoerculum was displaced ventrally during fossilization and would have been located directly ventral to the dermohyomandibular in life.
The large hollow between the preoperculum and the cleithrum also suggests anterior displacement of the preoperculum. In between the dermo-hyomandibular/preoperculum complex and the cleithrum would have been the operculum. However, there is no evidence of an operculum preserved on this specimen.
A bone that is interpreted to be a branchiostegal ray is located ventral to the antero-ventral margins of the preoperculum/dermohyomandibular complex. It is a singular ray and is heavily disarticulated. The anterior margin is flat with a slightly crushed antero-ventral border due to preservation bias. The dorsal margin is concave while the ventral margin is slightly convex. The rocky matrix overlies the posterior end of the bone so the true length of the bone is unknown, but the width of the ray is broad.
Pectoral girdle. A narrow, slightly crescent-shaped bone found posterior to the probable preoperculum/hyomandibular complex is most likely the cleithrum ( Fig. 1 View Figure 1 ). The cleithrum is a very large bone, which encircles the whole opercular skeleton ( Fig. 1 View Figure 1 ). Much of this bone is obscured by rocky matrix, making its morphology difficult to discern. The dorsal margin is posterior to the orbit. The bone widens to a large medial extension and a similar extension is found ventral to the opercular skeleton. Some parts of the bone that are exposed are located postero-medially and are ornamented with pits and ridges, the latter becoming more common closer to the posterior margins of the cleithrum.
The antero-dorsal arm of the cleithrum is found posterior to the caudal plate of the orbit. This arm is ornamented with small vertical ridges and pits. Two ridges are present along the outer margins throughout the entire bone. The ventral tip of this arm appears to be pointed judging by how the bone narrows towards the tip, but the rocky matrix between the sutures obscures this area. The cleithrum arm broadens towards the dorsal margin. The dorsal margin appears to be fragmented but seems convex forming two bifurcations. It is dorsal to the dermo-hyomandibular. These are broad with rounded margins.
Axial skeleton. There are seven autogenous neural spines preserved ( Fig. 1 View Figure 1 ). They are located posterior to the post-parietal process, dorsal to the cleithrum and anterior to the preserved imprint of notochord. These spines are long, thin bones with a large anterior flange, which tapers to a point distally, but no tips of the autogenous neural spines are preserved. The first four spines are overlain by the tips of the dorsal flank scales, confirming the decrease in size of the scales as they continue into a cephalo-caudal direction.
There are 21 neural spines excluding those in the caudal peduncle ( Fig. 1 View Figure 1 ). All are preserved as imprints in the rock, with a single exception. This spine is the first neural spine in the series and it is located posterior to the last autogenous neural spine. It is an elongate and thin bone with a broad anterior flange.
No haemal spines are preserved as bone ( Fig. 1 View Figure 1 ). Twelve imprints of haemal spines are observed but since there is a 1:1 distribution of haemal to neural spines there is presumably a minimum of at least 21 haemal spines in this specimen. Anterior haemal spines are obscured by the rocky matrix and ventral flank scales. No articulation between the neural and haemal spines is preserved along the open notochordal canal. Although the arcocentra of neither haemal nor neural spines are preserved, the impressions on the fossil hint at these spines having an expanded base that would have contacted the arcocentra of the adjacent neural and haemal spines, suggesting that there was a simple contact between these arcocentra.
Dorsal and anal fins. All remains of both the dorsal and anal fin are preserved as imprints only ( Fig. 1 View Figure 1 ). The preserved dorsal fin is comprised of seven fin rays, which are supported by 11 axonosts. The dorsal fin faces in an anterior direction and the fin rays simultaneously increase in size in an anterior direction. The fin is greatly elongate and has a falcate shape. Due to the very low number of fin rays and the position of the fin, disarticulation seems likely. A large V-shaped gap is located between the neural spines underneath the first three dorsal axonosts. This gap is most likely due to disarticulation.
The anal fin is located posterior to the post-coelomic bone. It is comprised of six fin rays. The fin rays increase greatly in size in a cephalocaudal direction. The anal fin has an elongate, falcate shape similar to the dorsal fin. There is no evidence of further fin rays being preserved, which would make this a very distinctive anal fin for a pycnodont fish. The number of axonosts is unknown as they are not preserved.
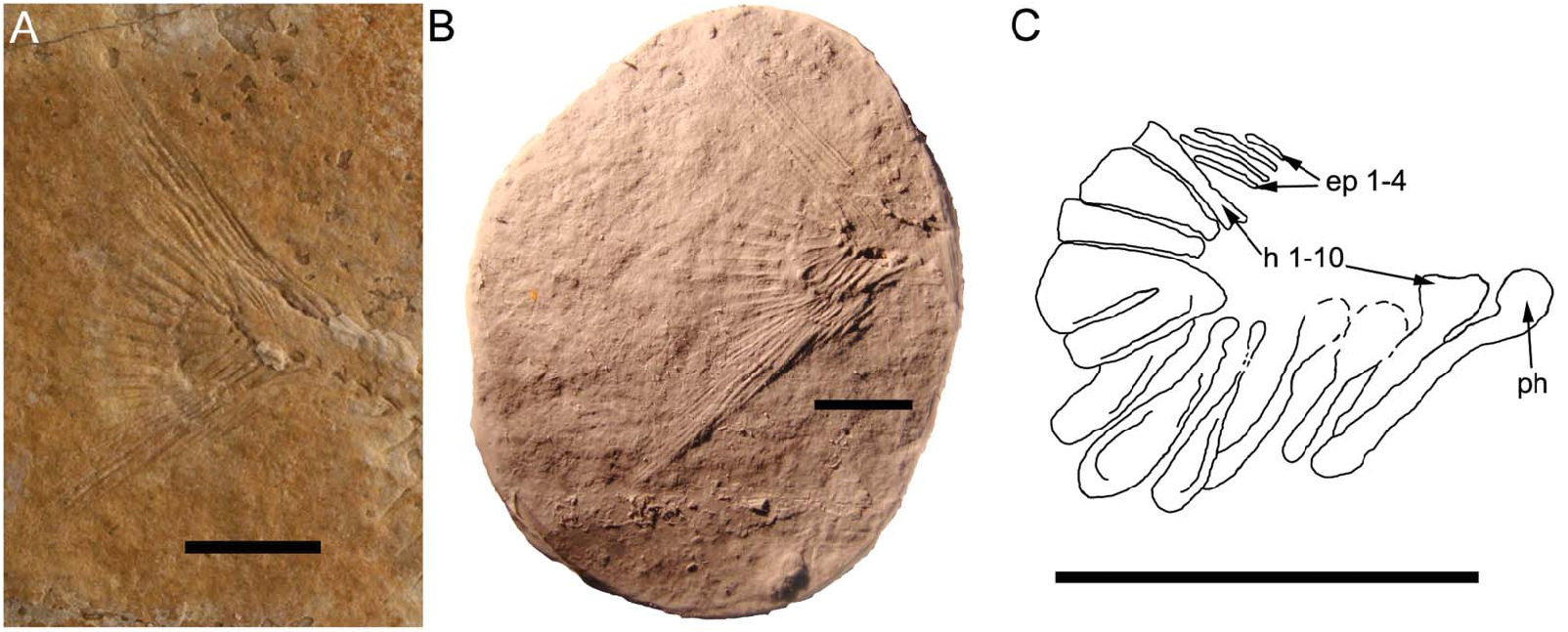
Caudal endoskeleton and fin. The caudal peduncle contains four neural spines preserved as bone, and four haemal spines preserved as imprints in the rock ( Figs 1 View Figure 1 , 4A View Figure 4 ). All neural spines curve backwards and have a large fan-shaped base with crenulations on its ventral margins. The second most posterior spine has a concave dorsal margin. The most posterior spine has a large anterior lateral process protruding from the antero-dorsal margin. No tips of spines are preserved, but the imprints in the rock show that they extended in a posterior direction and that the tips were as long as the remainder that was preserved.
The cast of the caudal fin imprint reveals additional details about its morphology ( Fig. 4B, C View Figure 4 ). There is a large parhypural located anteriorly to the hypochordal series. It is a club-shaped bone with a large ventral arcocentrum that curves backwards and appears to have overlapped the base of the first hypochordal. There are 10 hypochordals in total. Hypochordal 1 has a large expanded head with a narrow body that is inclined in an anterior direction. Hypochordals 2 and 3 are spoon shaped, with hypochordal 3 being the longer of the two. Hypochordals 4–7 have narrow heads with rounded margins and are fan-shaped bones which show evidence of hypertrophy due to the presence of longitudinal crests. The head of hypochordal 6 is overlain by hypochordal 7 but it still retains the fan shape of the two preceding hypochordals. Hypochordal 7 appears to be a compound bone as it shows a separation of the bone medially and at the anterior margin but not at the base. Hypochordal 8 is fan shaped but narrower than the preceding fan-shaped hypochordals, while hypochordal 9 is significantly broader. Hypochordal 10 appears to have a similar shape to hypochordal 8 with the exception of having a broader base. There are no longitudinal crests present on hypochordals 8–10.
There are only four epichordals observed. They are located ventral to the fin rays and anterior to hypochordal 10. The imprints reveal that they were elongate, thin bones that reach at least to the antero-medial border of the last hypochordal in length. Epichordal 1 is the smallest of the bones while epichordals 2–3 are far longer, being twice the length of the first epichordal. Epichordal 4 is only moderately longer than the first epichordal.
No fin rays are preserved as bone ( Figs 1 View Figure 1 , 4A View Figure 4 ). Thirty to 31 fin rays are preserved as imprints in the rock. The general shape of the fin suggests that dorsal and ventral outer fin rays extend outwards c. 3 cm from the caudal peduncle. The median fin rays barely extend outwards from the hypochordal bones (c. 4 mm). The imprint of the caudal fin margin suggests that it has a convex median distal border.
Post-coelomic bone. Located posterior to the cloaca is the post-coelomic bone ( Figs 1 View Figure 1 , 5 View Figure 5 ). The bone itself is not preserved but the shape is clearly imprinted into the rock. It is a long, thin bone, which broadens to a club near its distal tip. The anterior portion of the club-like bone has convex dorsal and ventral margins while the medial margin is flat. The posterior margin of this bone is flat. It can be seen that the bone curves in an anterior direction.
Squamation. Posterior to the dermosupraoccipital is a large spine (twice as tall as preceding ossification) with a broad base, which narrows in a ventro-dorsal direction towards the distal tip. The ridge present on its anterior margin disappears as it reaches the distal tip of the scale. The tip of the spine curves anteriorly with the dorsal and ventral margins having a convex contour and a concave contour, respectively. The spine is broken off at tip, which is displaced ventrally to the concave margin of the spine ( Fig. 2A, C View Figure 2 ). The tip of the spine has a rounded margin. This spine represents the first dorsal ridge scale. This scale originates from the posterior base of the dermosupraoccipital where it is incorporated into the skull roof ( Figs 1 View Figure 1 , 2A–C View Figure 2 ).
The second dorsal ridge scale shows that the posterior portion has a large rectangular margin. The first dorsal ridge scale also has this rectangular extension but it is overlain by a displaced scale. The second dorsal ridge scale also appears to be narrowing to a distal tip anteriorly but this is not observed directly, instead being deduced from the concave notch present in the anterior portion of the scale before it is broken off ( Fig. 2D View Figure 2 ).
Three dorsal ridge scales are preserved as bone and six are preserved as imprints in the rocky matrix, making a total of nine scales ( Fig. 1 View Figure 1 ). The preserved scales posterior to the first dorsal ridge scale are smooth with a flat dorsal margin and no spines. The scale has a medial ridge surrounded by a posterior rectangular extension and a narrowing anterior portion with a concave notch. There is a long, thin pointed antero-ventral process protruding from the dorsal ridge scale while it overlies the flank scales. The few scales that are preserved in bone increase in width in a cephalo-caudal direction.
Postero-ventral to the cleithrum are the ventral ridge scales ( Fig. 1 View Figure 1 ). There is a total of 10 scales preserved in bone. Like their dorsal ridge scale counterparts, an increase in scale width can be observed in a cephalo-caudal direction. The best-preserved scale is located posterior to the cloaca. It has a smooth surface and two spines not in contact with each other. Flank scales located anterior to the cloaca are slimmer in width, and two appear dorsal to the anterior and posterior ends of the ventral ridge scale. On the scales where the spines are preserved, there appears to be a maximum number of three. The spines are more visible in scales when observed in a cephalo-caudal direction.
The flank scales are thin, elongate scale bars that are ventral to the dorsal ridge scales and dorsal to the ventral ridge scales ( Fig. 1 View Figure 1 ). The dorsal flank scales have a medial ridge going down the length of the scale bar. The flank scales curve posteriorly. These flank scales decrease in length the closer they are to the dorsal fin. On the ventral side, flank scales are singular, thin and elongate ventral to the cleithrum. Posterior to the cleithrum, they become much broader with numerous ridges (usually two to three) on the surface. These reach the notochord and are not preserved in the medial portion of the fossil. No articulation is seen between the dorsal and ventral flank scales, so the number of complete scale rows remains unknown.
The cloaca is not roofed by a bifid scale. A fragment of the scale that forms the roof of the cloaca can be observed in an antero-ventral position. A slight concave notch is present ventrally on the preserved portion of the scale. No modified cloacal scales as originally described by Poyato-Ariza & Wenz (2002, p. 203) are present in this specimen. Instead, there are what appear to be bifurcating scales located anterior and posterior to the cloaca ( Figs 1 View Figure 1 , 5 View Figure 5 ). The anterior cloacal scale bifurcates in a vertical, slightly anterior direction. The left branch is ornamented with small ridges while the right branch is smooth. Bifurcation begins closer to the anterior portion of the scale that roofs the cloaca. The anterior margin is concave while the posterior margin is convex. The posterior cloacal scale begins to bifurcate vertically above the cloaca. The ventral base is broad and curves anteriorly revealing a convex margin. Both branches are preserved as an imprint in the rocky matrix. The left branch is significantly longer than the right and becomes narrower at its tip. Bifurcation of the posterior cloacal scale is also preserved as an imprint. The true length of the right branch is unknown as the tip is overlain by disarticulated flank scales. We have reconstructed it to be of the same height as the left branch ( Fig. 5B View Figure 5 ). Both bifurcated cloacal scales are dorsal to the ventral ridge scales. No scales are preserved posterior to the post-coelomic bone.



| UV |
Departamento de Biologia de la Universidad del Valle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Order |
|
|
Family |
|
|
Genus |