
Profundiconus neocaledonicus, Tenorio & Castelin, 2016
|
publication ID |
https://doi.org/10.5852/ejt.2016.173 |
|
DOI |
https://doi.org/10.5281/zenodo.5640347 |
|
persistent identifier |
https://treatment.plazi.org/id/24768796-CD2D-FFF6-FD8F-1329FEF9F84A |
|
treatment provided by |
Plazi |
|
scientific name |
Profundiconus neocaledonicus |
| status |
sp. nov. |
Profundiconus neocaledonicus View in CoL sp. nov.
urn:lsid:zoobank.org:act:6581A3CA-DA2E-44A2-8E6E-2600CCA3B1D3
Figs 15A–J View Fig. 15 , 16A–C View Fig.16 , 17 View Fig. 17
Conus profundorum View in CoL – Rolan & Raybaudi-Massilia 1994: 33, pl. 11, fig. 82 (non Chelyconus ( Profundiconus) profundorum Kuroda, 1956 View in CoL ). — Röckel et al. 1995a: 381, pl. 27, fig. 16. — Röckel et al. 1995b: 563, fig. 22.
Profundiconus cf. profundorum View in CoL – Puillandre et al. 2014: Supplementary Material 1 (unfigured).
Etymology
The epithet of this species makes reference to its distribution in deep water around New Caledonia.
Type material examined
Holotype
NEW CALEDONIA: 45.9 × 21.7 mm, R/V Alis , TERRASSES Expedition, st. DW 3076, Banc Crypthélia, Norfolk Ridge , 23°14'12" S, 168°13'18" E, 390–570 m ( MNHN IM-2009-18227 ; Fig. 15A View Fig. 15 ; GenBank accession number ( cox1 sequence): KJ550428 View Materials ). View Materials GoogleMaps
Paratypes
NEW CALEDONIA: 61.3 × 28.1 mm, R/V Alis , EXBODI Expedition, st. DW 3889, Mont Vauban Partie SE , 22°25' S, 171°41' E, 350 m ( MNHN 2009-31323 About MNHN ; paratype 1; Fig. 15B View Fig. 15 ); View Materials GoogleMaps 52.6 × 24.7 mm, R/V Alis , SMIB 8 Expedition, st. DW 167, Norfolk Ridge , 23°38' S, 167°43' E, 430–452 m ( MNHN IM-2000-30780 ; paratype 2; Fig. 15C View Fig. 15 ) GoogleMaps ; 46.7 × 20.4 mm, R/V Alis , TERRASSES Expedition, st. DW 3060, Banc Stylaster, Norfolk Ridge , 23°39' S, 167°44' E, 440–450 m ( MNHN IM-2009-18225 ; paratype 3; Fig 15G View Fig. 15 ; GenBank accession number ( cox1 sequence): KJ550427 View Materials ) GoogleMaps ; 71.8 × 37.0 mm, R/V Coriolis , CHALCAL 2 Expedition, st. DW 82 , S New Caledonia, 23°14' S, 168°04' E, 304 m ( MNHN IM-2000-30782; paratype 4; Fig. 15D View Fig. 15 ) GoogleMaps ; 92.0 × 42.0 mm, R/V Alis , NORFOLK 1 Expedition, st. DW 1732, Banc P, Norfolk Ridge , 23°20' S, 168°16' E, 347–1063 m ( MNHN IM-2000-30783 ; paratype 5; Fig. 15E View Fig. 15 ) GoogleMaps ; 65.1 × 32.5 mm, R/V Alis , MUSORSTOM 6 Expedition, st. DW 406, Loyalty Ridge , 20°41' S, 167°07' E, 373 m ( MNHN IM-2000-30784 ; paratype 6; Fig. 15H View Fig. 15 ) GoogleMaps ; 54.5 × 25.4 mm, R/V Coriolis , CHALCAL 2 Expedition, st. CC1 , S New Caledonia, 24°55' S, 168°22' E, 500 m (MNHN IM-2000-30785; paratype 7; Fig. 15I View Fig. 15 ) GoogleMaps ; 67.3 × 33.0 mm, R/V Alis , NORFOLK 1 Expedition, st. DW 1657, Banc N, Norfolk Ridge , 23°28' S, 167°52' E, 305–332 m ( MNHN IM-2000-30786 ; paratype 8; Fig. 15J View Fig. 15 ) GoogleMaps ; 61.5 × 28.7 mm, R/V Alis , NORFOLK 1 Expedition, st. DW 1658, Banc N, Norfolk Ridge , 23°26' S, 167°50' E, 320–336 m ( MNHN IM-2000-30787 ; paratype 9) GoogleMaps ; 65.5 × 35.7 mm, R/V Alis , NORFOLK 1 Expedition, st. DW 1652, Banc N, Norfolk Ridge , 23°26' S, 167°50' E, 290– 378 m ( MNHN IM-2000-30788 ; paratype 10) GoogleMaps ; 53.4 × 22.8 mm, R/V Alis , BATHUS 3 Expedition, st. CP 811, Norfolk Ridge , 23°41' S, 168°15' E, 383–408 m ( MNCN 15.05/60172; paratype 11) GoogleMaps .
Type locality
NEW CALEDONIA: Banc Crypthélia, Norfolk Ridge, 23°14' S, 168°13' E, 390–570 m (TERRASSES Expedition st. GoogleMaps DW3076).
Other material examined
NEW CALEDONIA: 39.9 × 18.8 mm, R/V Alis, NORFOLK 2 Expedition, st. DW 2156, Ile des Pins, 22°54' S, 167°15' E, 468–500 m ( MNHN IM- 2007-34866; GenBank accession number ( cox1 sequence): KJ550426 View Materials ). This specimen was databased (http://coldb.mnhn.fr/catalognumber/mnhn/im/2007-34866) and sequenced, but the shell was destroyed in the process and is no longer available.
Additionally, we examined 32 more specimens from 18 uncataloged MNHN lots collected at several stations in Norfolk Ridge and Loyalty Ridge, New Caledonia, in the course of several campaigns.
Description
Morphometric parameters: S L = 45–80 mm; RD = 0.54–0.66; RSH = 0.16–0.22; PMD = 0.81–0.92.
Medium-sized to moderately large (maximum length 92.0 mm). Shell profile ventricosely conical to conical, with rounded shoulder and spire low to moderately high. Spire profile sigmoid, occasionally slightly concave. Protoconch multispiral of 3 or more whorls, white to pale violet-brown ( Fig. 15F View Fig. 15 ). First 4–7 postnuclear whorls nodulose. Teleoconch sutural ramp flat, slightly concave or sigmoid in later whorls, smooth, with cords absent. Last whorl smooth, with fine spiral ribs at base. Ground colour creamy-white to cream. Last whorl with two broad violet-brown, light brown or tan spiral bands above and below midbody, which exhibits broad ground-coloured spiral band. Colour is darker towards base, usually purplish. Narrow ground-colour spiral band often present at height of shoulder. Spire patternless, of ground colour, occasionally showing diffuse pale violet-brown or light brown on top of teleoconch whorls. Aperture light to pinkish brown. Periostracum olive, thin, translucent, smooth. Operculum with lateral serrations.
Radular tooth examined in paratypes 1 ( Fig. 16A View Fig.16 ), 2 and 7 ( Fig. 16B, C View Fig.16 ). 34–45 teeth in radular sac. Radular tooth of rather small relative size: its total length relative to shell length S L/T L = 61–87. Anterior portion shorter than posterior section of tooth (T L/AP L = 3.1–3.4). With one barb and pointed, prominent blade which covers 43–54% of apical portion of tooth. External cusp present, extending between 65 and 88% of length of anterior portion of tooth. External cusp not much widened laterally and serrations can be indistinct, with only 0–3 small blunt denticles. Large adapical opening occupying most of anterior portion of tooth (100AO L/AP L = 62–75). With characteristic fringe of closely spaced projections pointing towards the apex located immediately below waist ( Fig. 16C View Fig.16 ). Shaft fold present. Large and prominent basal spur on top of slanted base of tooth.
Distribution and habitat
Norfolk Ridge and Loyalty Ridge, New Caledonia, at depths from 290 to 1100 m ( Fig. 17 View Fig. 17 ). A couple of empty shells resembling P. neocaledonicus sp. nov. from Aliguay Island, Philippines, have been examined. The identity of these specimens from the Philippines (coming from the John K. Tucker collection, now with the INHS, Illinois, USA) could not be confirmed by radular or molecular studies. This observation might suggest an extension of the distribution range to the Philippines, but would require additional substantiation.
Remarks
Profundiconus neocaledonicus sp. nov. has long been considered a local form of P. profundorum from New Caledonia ( Röckel et al. 1995a). P. profundorum was originally described based upon material from Japan ( type locality: “Off Tosa, Japan; 100+ fathoms”) ( Fig. 1A View Fig. 1 ). In recent times, specimens of P. profundorum have become available from China ( Fig. 1L View Fig. 1 ). These specimens are characterised by their significantly larger mean shell length compared to the Japanese specimens (mean profundorum from Japan: 74.3 mm; mean profundorum from China: 111.05 mm; t = 13.12, p = 0; U SL= 0, p = 0.000028). Apart from differences in SL shell length, ANCOVA indicated no statistically significant differences between the morphometric parameters of P. profundorum from Japan and those from China (ANCOVA on MD: F = 1.94, p = 0.18; on HMD:F = 3.44, p = 0.08; on SH: F = 0.05, p = 0.82). Hence, we consider the Chinese and Japanese populations conspecific. Compared to P. profundorum , the shells of P. neocaledonicus sp. nov. from New Caledonia have a different shape, are consistently smaller in average shell length, and are also paler in colour. Table 3 View Table 3 shows the results of the ANCOVA with the morphometric shell parameters MD, HMD and SH as variables, and using species hypothesis as factor and S L as covariate. Least-squares means are listed, along with average shell lengths for each of the species.
These results confirm that the shell of P. neocaledonicus sp. nov. is significantly broader, more conical (higher HMD) and lower-spired than the shell of P. profundorum , which also has a larger mean S L. A discriminant function analysis (DFA) using MD, HMD, SH and S L as variables and species hypothesis as factor correctly classified 94.4% of the specimens (all correct except two specimens of profundorum misclassified as neocaledonicus and one specimen of neocaledonicus misclassified as profundorum ). A DFA excluding S L from the set of variables correctly classified 92.6% of the specimens (three profundorum were misclassified as neocaledonicus and one specimen of neocaledonicus misclassified as profundorum ). These results indicate that P. neocaledonicus sp. nov. can be separated with a high degree of certainty from P. profundorum based on significant differences in size and shell shape. It was not possible to compare the radula of P. neocaledonicus sp. nov. with that of P. profundorum . The radular tooth morphology of P. profundorum is unknown, since all the published information ( Rolán & Raybaudi-Massilia 1994) actually corresponds to specimens from New Caledonia, here introduced as P. neocaledonicus sp. nov. No preserved specimens of P. profundorum from Chinese or Japanese localities were available for radular or molecular analyses.
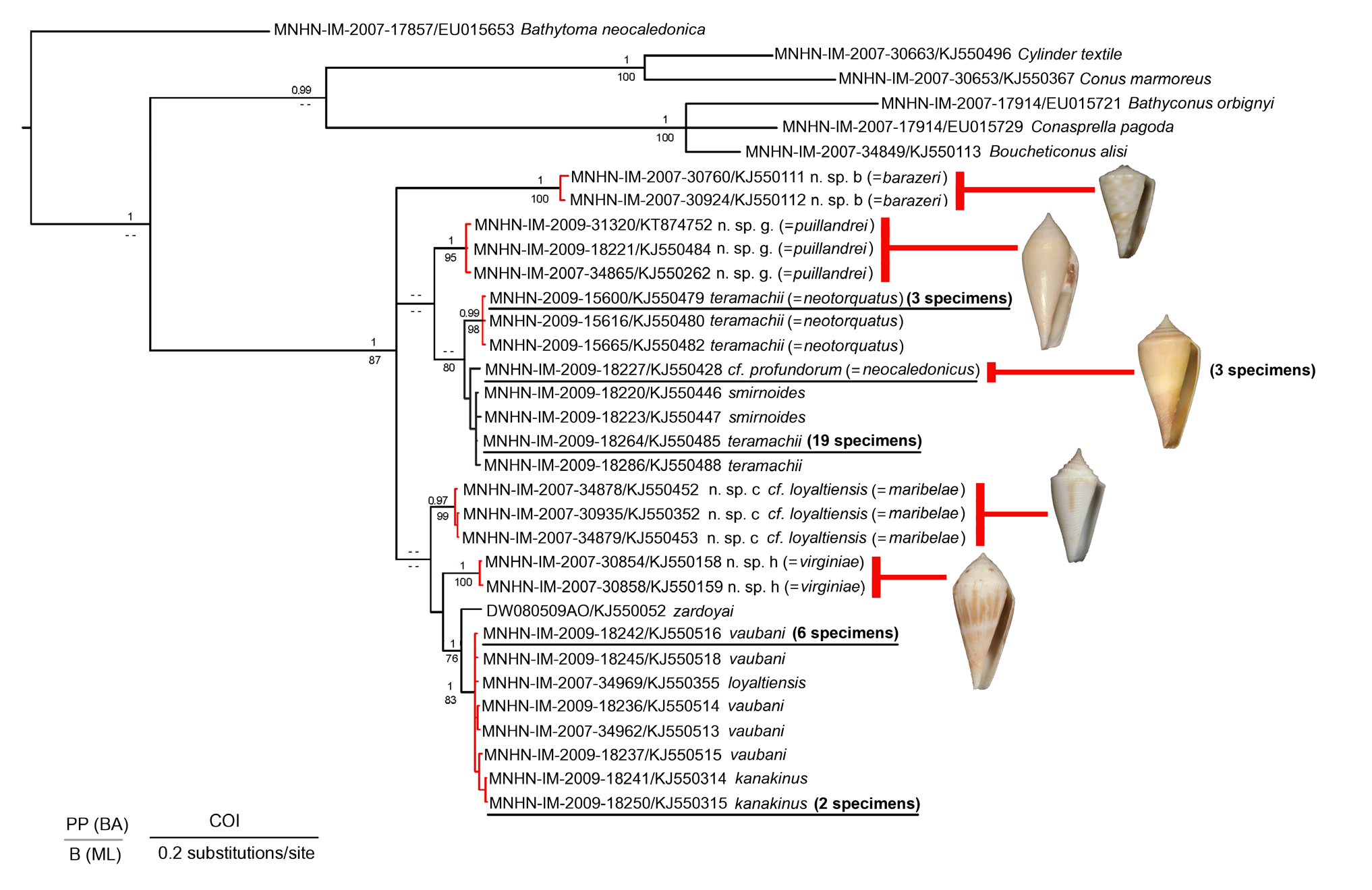
Although the genetic differentiation of P. neocaledonicus sp. nov. from P. smirnoides and P. teramachii was not supported at the nodes on the tree in Fig. 5 View Fig. 5 , P. neocaledonicus sp. nov. is easily separated from the sympatric species P. smirnoides based on shell and radular morphology. The shell of P. smirnoides ( Fig. 1D View Fig. 1 ) is significantly narrower, less conical and much more highly spired than that of P.neocaledonicus sp. nov. (ANCOVA with S L as covariate: on MD: F = 103.91, p = 0; on HMD: F = 62.54, p = 0; on SH: F = 28.37, p = 0). It also has a significantly larger mean shell length ( 76.09 mm for smirnoides versus 57.03 mm for neocaledonicus , t = -4.299, p = 0.00011; U = 285.5, p = 0.00055). P. smirnoides has a pattern of brown spiral bands on each side of centre interrupted by creamy white axial streaks, whereas in P. neocaledonicus sp. nov. is much simpler, consisting of pale purplish-brown broad bands on each side of centre on a creamy-white ground colour. Comparison of the large and elongated radular tooth of P. smirnoides ( Fig. 9H View Fig. 9 ) with that of P. neocaledonicus sp. nov. ( Fig. 16 View Fig.16 ) also allows the straightforward separation of the two species. P. teramachii ( Fig. 1B View Fig. 1 ) has a very different shell, larger in size, and is patternless, with a characteristic stepped spire and with a broadly carinated shoulder, sometimes with densely set rounded tubercles, particularly in smaller adults.
The species P. puillandrei sp. nov., which lives sympatrically with P. neocaledonicus sp. nov., has a significantly smaller shell length (t = 4.296, p = 0.0001; U = 53, p = 0.000066), with differences in morphometric parameters that suggest that P. puillandrei sp. nov. has a narrower shell with a higher spire (ANCOVA with S L as covariate on MD: F = 6.23, p = 0.0168; on HMD: F = 40.19, p = 0; on SH: F = 35.46, p = 0). This species shows a characteristic ridge at the shoulder, sometimes nodulose, whereas in P. neocaledonicus sp. nov. the shoulder is always rounded. The shell of P. puillandrei sp. nov. is patternless and does not exhibit the banding pattern visible in the shell of P. neocaledonicus sp. nov.
Röckel et al. (1995a: pl. 73, figs 17–18) illustrated one specimen identified as “ Conus species no. 32”. This specimen measures 50 × 23 mm and comes from Nazca Ridge. Other specimens like the one illustrated in Röckel et al. (1995a) have been collected in deep-water on several seamounts across Sala y Gómez and Nazca Ridges in the course of several campaigns carried out by Russian research vessels in the 70’s and 80’s. The biota of the Nazca and Sala y Gómez submarine ridges was reviewed in Parin et al. (1997). The fauna of benthic and benthopelagic invertebrates of this area is much more closely related to the Indo-West Pacific than to the Eastern Pacific fauna and is characterized by a very high degree of endemism at the species level (51% among identified bottom invertebrates). We have been unable to examine the specimens from the Nazca Ridge area, but the available photos ( Röckel et al. 1995a) show a striking resemblance to the shells of P. neocaledonicus sp. nov. collected on Norfolk Ridge, New Caledonia, at a distance of c. 10 500 km to the east. Further research might eventually prove their conspecificity, which would imply a considerable range extension for this species, or alternatively disclose a new species of Profundiconus most likely endemic to the Sala y Gómez and Nazca Ridge areas.





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Caenogastropoda |
|
Order |
|
|
SuperFamily |
Conoidea |
|
Family |
|
|
Genus |
Profundiconus neocaledonicus
| Tenorio, Manuel J. & Castelin, Magalie 2016 |
Conus profundorum
| Rolan & Raybaudi-Massilia 1994 : 33 |
| Röckel et al. 1995a : 381 |
| Röckel et al. 1995b : 563 |