
Plestiodon
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2011.00801.x |
|
persistent identifier |
https://treatment.plazi.org/id/25042F0C-4A01-1D41-15D7-1097FB7EFC70 |
|
treatment provided by |
Marcus |
|
scientific name |
Plestiodon |
| status |
|
OF PLESTIODON
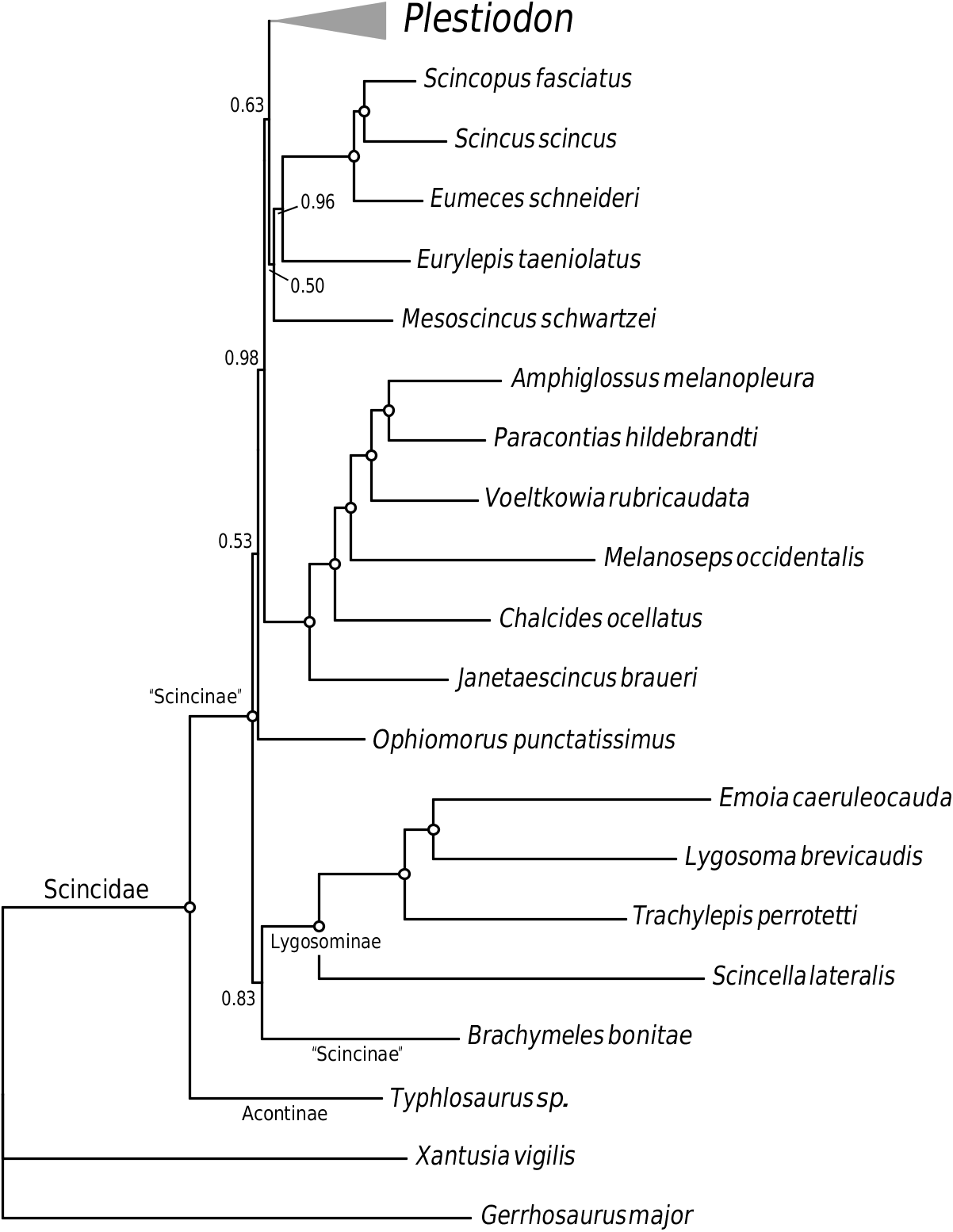
Although they constitute the largest lizard family in terms of species, skinks have only recently been the subject of molecular phylogenetic analysis. These studies ( Whiting et al., 2003; Brandley et al., 2005, 2011; Austin & Arnold, 2006; Siler & Brown, 2011; Siler et al., 2011; Skinner et al., 2011) have both supported and refuted many of the relationships proposed by previous morphological analyses ( Taylor, 1935; Greer, 1970a, b). However, many relationships, especially ‘deep’ relationships among the major skink lineages, have remained poorly supported, or in at least one case (the monophyly of Lygosominae ), completely conflicting ( Greer, 1986; Whiting et al., 2003; Brandley et al., 2005, 2011; Siler & Brown, 2011; Siler et al., 2011; Skinner et al., 2011). In his pioneering evolutionary taxonomy of skinks, Greer (1970a, b) identified four scincid subfamilies: Acontinae , Feylinae, Lygosominae , and Scincinae , and assumed that Scincinae was a group from which the three other subfamilies were derived (thereby rendering it paraphyletic). Subsequent studies have demonstrated conclusively that the enigmatic Feylinae (not included in this study) is closely related to the southern African ‘scincines’ Melanoseps and Typhlacontias ( Whiting et al., 2003; Brandley et al., 2005). However, a wellsupported phylogenetic placement of the lygosomines with major ‘scincine’ lineages has remained elusive. Our study does much to revise the existing phylogenetic framework of skinks resolving several additional ‘deep’ relationships, including the placement of Acontinae , Lygosominae , and ‘Scincinae’.
Our results strongly support a basal split within Scincidae between the limbless acontines (represented by Typhlosaurus sp. in Figs 2 View Figure 2 and S 1 View Figure 1 ) and all other skinks, thereby corroborating the results of Whiting et al. (2003) and Skinner et al. (2011). This phylogenetic relationship has bearing on the evolution of limb reduction in skinks. Complete limblessness has evolved independently ~25 times among squamate reptiles, with the majority of these derivations ocurring within Scincidae ( Greer, 1991; Wiens et al., 2006; Brandley et al., 2008; Siler et al., 2011). Although we lack sufficient phylogenetic evidence to evaluate the ancestral body plan of scincid lizards, that acontines represent one of the two earliest lineages of crown Scincidae suggests that limb reduction may have been a feature of scincid evolution for a very long time (79–114 Mya; Brandley et al., 2008, 2011).
Lygosomines represent the bulk of species diversity in skinks. Although our sampling of lygosomines is low, we sampled four of its five major lineages: the Eugongylus group [represented by Emoia caeruleocauda (De Vis, 1892) ], the Lygosoma group [ Lygosoma brevicaudis Greer, Grandison, & Barbault, 1985 ], the Mabuya group [ Trachylepis perrotetii (Duméril & Bibron, 1839) ], and the Sphenomorphus group [ Scincella lateralis (Say, 1823) ]; but not the Egernia group. This therefore allows us to make a cursory evaluation of competing hypotheses of ‘deep’ lygosomine relationships. Molecular studies that have focused specifically on lygosomine relationships have supported the Sphenomorphus group as the sister lineage to all other lygosomine skinks ( Honda et al., 2000, 2003; Reeder, 2003; Austin & Arnold, 2006; Linkem, Diesmos & Brown, 2011; Skinner et al., 2011), a result congruent with our analysis of the concatenated data ( Fig. 2 View Figure 2 ). However, the relationships of the remaining groups differ among these studies. With the caveat that we did not sample the Egernia group, our results support Reeder (2003) and Skinner et al. (2011) who inferred strong support for a clade composed of ( Mabuya ( Lygosoma + Eugongylus )) groups. That Austin & Arnold (2006) did not sample the Lygosoma group makes comparison with our study uninformative. With the exception of the placement of the Sphenomorphus group, our results are completely incongruent with Honda et al. (2000, 2003), but we note that these relationships were not strongly supported in those studies.
With one exception, the individual gene tree analyses either support the same relationships as the concatenated data or are not strongly incongruent; the mtDNA gene tree ( Fig. S2 View Figure 2 ) supports a sister relationship between the Mabuya and Sphenomorphus group. We speculate that this relationship is explained by homoplasy resulting from a combination of the relatively rapid evolution of mtDNA and the relatively old age of lygosomines (see Brandley et al., 2011; Skinner et al., 2011): a problem that is probably exacerbated by our low level of taxon sampling. Even with explicit model-based methods (e.g. maximum likelihood and Bayesian), extreme homoplasy can nonetheless lead to high support for incorrect relationships ( Felsenstein, 1978, 1985; Brandley et al., 2006, 2009).
When compared with previous molecular phylogenetic studies of scincid relationships, perhaps the most notable result in the current study is an increased resolution among the ‘scincine’ genera. Our multilocus phylogenetic analysis reveals multiple, well-supported novel ‘scincine’ relationships. Although previous studies have inferred a close phylogenetic affinity of the ‘scincine’ genera inhabiting Africa, Madagascar, and the Seychelles, ours is the first to infer very strong support for the interrelationships of many of these lineages. We find the Seychellois Janetaescincus , North African and Mediterranean Chalcides (and presumably Sphenops ; Brandley et al., 2005; Carranza et al., 2008), Southern African Melanoseps , and Malagasy Voeltzkowia and Amphiglossus + Paracontias form progressively more exclusive clades. Only the mtDNA gene tree ( Fig. S2 View Figure 2 ) supports a strongly incongruent relationship by supporting a clade that is exclusive of Melanoseps . The geographical distribution of these genera suggests that the break-up of Gondwana played a major role in the phylogenetic history of the clade; however, inclusion of the Indian and Sri Lankan genera in future analyses will be critical for testing this hypothesis. We note that we did not fully sample other African, Malagasy, and Mauritian ‘scincine’ genera, but we can infer from other studies that they too are members of this larger clade ( Whiting et al., 2003; Brandley et al., 2005; Schmitz et al., 2005). We also infer strong support for the sister relationship of this putatively Gondwanan clade with the primarily Laurasian-distributed Eumeces s.l., Scincus , and Scincopus .
The phylogenetic affinities of Ophiomorus and Brachymeles are complex. Although our concatenated data analysis did not infer strong support for the placement of either genus, inspection of the 95% credible set of unique topologies reveals that 837 of 2816 trees are compatible with ‘scincine’ monophyly (not shown). In other words, although we cannot strongly support the placement of these genera, we also cannot statistically reject their placement in a monophyletic Scincinae . The RAG1 gene tree ( Fig. S 1g View Figure 1 ) also strongly supports ‘scincine’ monophyly. However, the SNCAIP tree strongly supports Brachymeles in a clade containing the four lygosomine genera ( Fig. S1h View Figure 1 ). Brandley et al.’s (2011) timecalibrated analysis of a smaller data set (see Brandley et al., 2011: appendix IV) infers strong support for the sister relationship of Brachymeles and Lygosominae (PP = 1.0), and the sister relationship of Ophiomorus and all other ‘scincines’ (PP = 0.96).
Different taxon and gene sampling may explain the discrepancies between Brandley et al. (2011) and the current study. An alternative explanation is that, unlike the current study, Brandley et al. (2011) used a relaxed molecular clock model of evolution that attempts to correct for rate heterogeneity amongst lineages for the purposes of divergence date estimation. Regardless, because the relationships of these two genera are strongly supported in Brandley et al. (2011), and our present phylogenetic results do not strongly conflict with that study, we argue that the Brandley et al. (2011) tree may be a better estimate of the relationships of Brachymeles and Ophiomorus in the absence of more phylogenetic evidence.
We infer strong support for the hypothesis that Plestiodon and other Eumeces s.l. genera do not represent the earliest diverging lineage of skinks. These results therefore refute Greer’s (1970a) hypothesis that ‘Morphologically, Eumeces [s.l.] is very possible the most primitive living skink taxon and may, in fact, be quite similar to the ancestor of all skinks’. Although the genus ‘ Eumeces ’ was long considered to be monophyletic, numerous recent studies have rejected this hypothesis ( Griffith et al., 2000; Schmitz et al., 2004; Brandley et al., 2005). These studies are also in concordance with karyotypic studies that have demonstrated that three of the four genera possess unique shared, derived karyotypes, 2 N = 32 in Eumeces s.s. ( Gorman, 1973; Caputo et al., 1993; Caputo, Odierna & Aprea, 1994), 2 N = 28 in Eurylepis ( Ivanov & Bogdanov, 1975; Kupriyanova, 1986; Eremchenko, Panfilov & Tsarinenko, 1992), and 2 N = 26 in Plestiodon (e.g. Deweese & Wright, 1970; McDiarmid & Wright, 1976; Kato et al., 1998). The karyotype of Mesoscincus is unknown. However, these molecular and karyotype studies are only able to reject monophyly, and are unable to elucidate with strong support the phylogenetic affinities of the four genera that were once part of Eumeces s.l. ( Eumeces s.s., Eurylepis , Mesoscincus , and Plestiodon ). The concatenated data tree, and all nine gene trees, support a clade composed of Eumeces s.s., Scincopus , and Scincus , to the exclusion of all other skink genera. Moreover, the concatenated data also support Eurylepis as the sister lineage to this clade. The precise phylogenetic affinities of Mesoscincus and Plestiodon remain elusive, although we note that our concatenated data tree at least excludes them from lygosomines, acontines, Ophiomorus , and Brachymeles .
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.