
Bombus keriensis Morawitz, 1887
|
publication ID |
https://doi.org/10.5852/ejt.2020.719.1107 |
|
publication LSID |
lsid:zoobank.org:pub:A4500016-C219-4353-B81C-5E0BB520547F |
|
DOI |
https://doi.org/10.5281/zenodo.14372078 |
|
persistent identifier |
https://treatment.plazi.org/id/252087CA-1F1B-9565-FDCA-FB5DD92EFEBA |
|
treatment provided by |
Valdenar |
|
scientific name |
Bombus keriensis Morawitz, 1887 |
| status |
|
Bombus keriensis Morawitz, 1887 View in CoL
Figs 16 View Figs 14‒16 , 163–172 View Figs 139–180 , 203 View Figs 199‒204 , 209 View Figs 209–210 , 212 View Figs 211–212
Bombus keriensis Morawitz, 1887: 199 View in CoL .
Lapidariobombus separandus subsp. meridialis Skorikov, 1914a: 127 View in CoL .
Bombus trilineatus View in CoL S.-F. Wang, 1982: 441.
Pyrobombus keriensis subsp. karakorumensis Tkalců, 1989: 57 View in CoL .
The taxonomic limits of the species B. keriensis have been a particular long-standing or persistent taxonomic problem.
Vogt (1909: 41, 62) treated B. keriensis s. str. as a species separate from his taxon separandus . Vogt (1911: 58) characterised and distinguished B. keriensis s. str. from B. separandus by having from a few yellow hairs on the face to having the hair of the face entirely pale.
Skorikov (1931) listed B. keriensis s. str. and B. separandus as two species but discussed them together under a joint heading, “ Lapidariobombus keriensis (F. Mor.) 1886 und Lapidariobombus separandus (Vogt) 1909 ”. While he mentioned the difference in face colour, he described B. keriensis s. str. and B. separandus as hardly distinguishable (his Lapidariobombus anargumentosus Skorikov, 1931 , on the same page is recognised here from the pattern of punctures on the ocello-ocular area of the head of the syntypes in the ZIN as conspecific with B. ( Pyrobombus) biroi Vogt, 1911 , syn. nov., and is not a species of the subgenus Melanobombus ).
Reinig (1935) included the taxon keriensis s. str. as a part of a broad species B. keriensis s. lat. that also included the taxa separandus + incertoides + alagesianus . This interpretation was followed by Williams (1991, 1998).
Wang (1982) described her taxon trilineatus without reference to B. keriensis , comparing it instead to the more distantly related “ B. occulatus var. haemorrhous Richards ” [sic] (= B. simillimus ) for colour pattern and details of morphology.
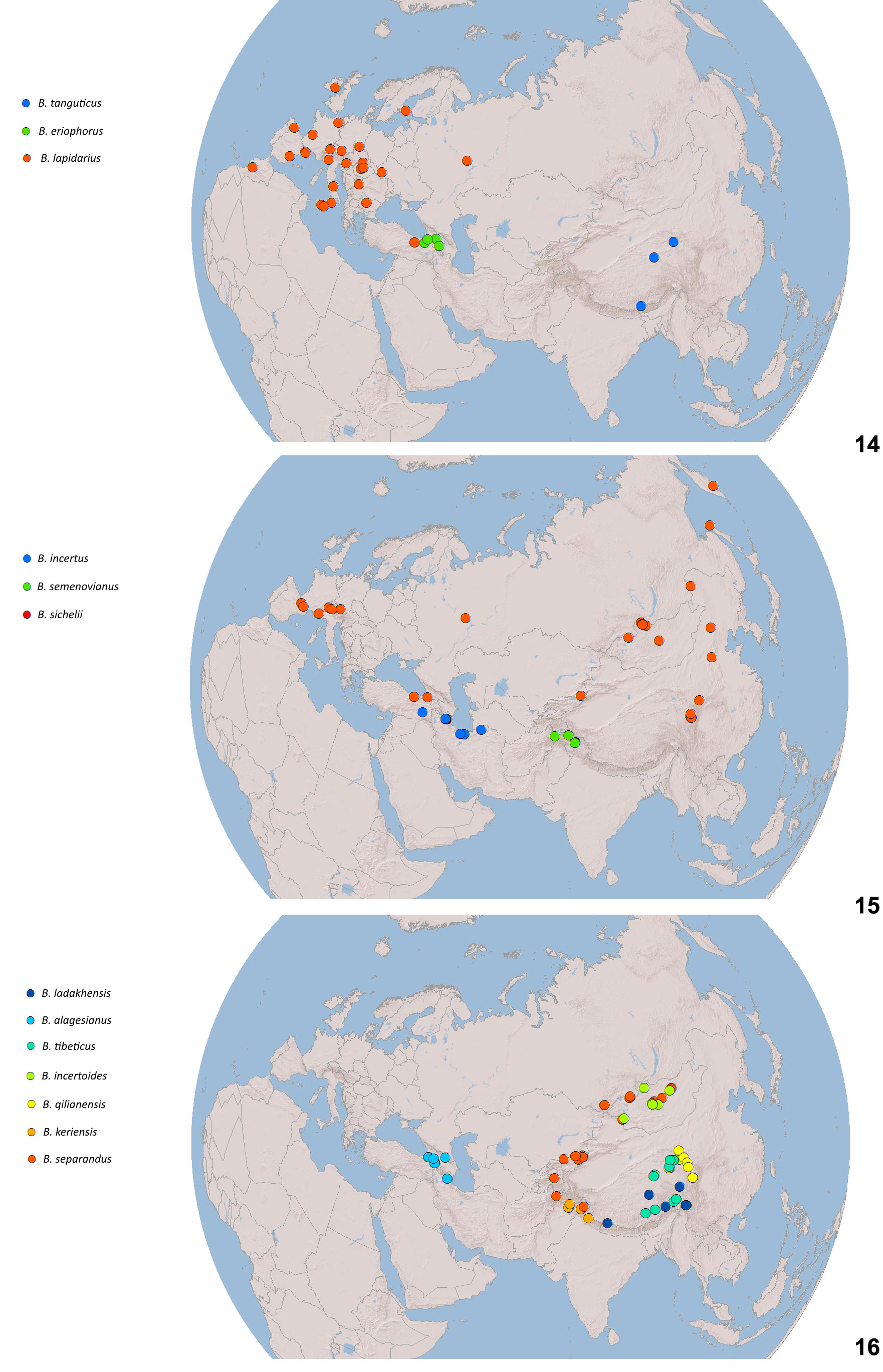
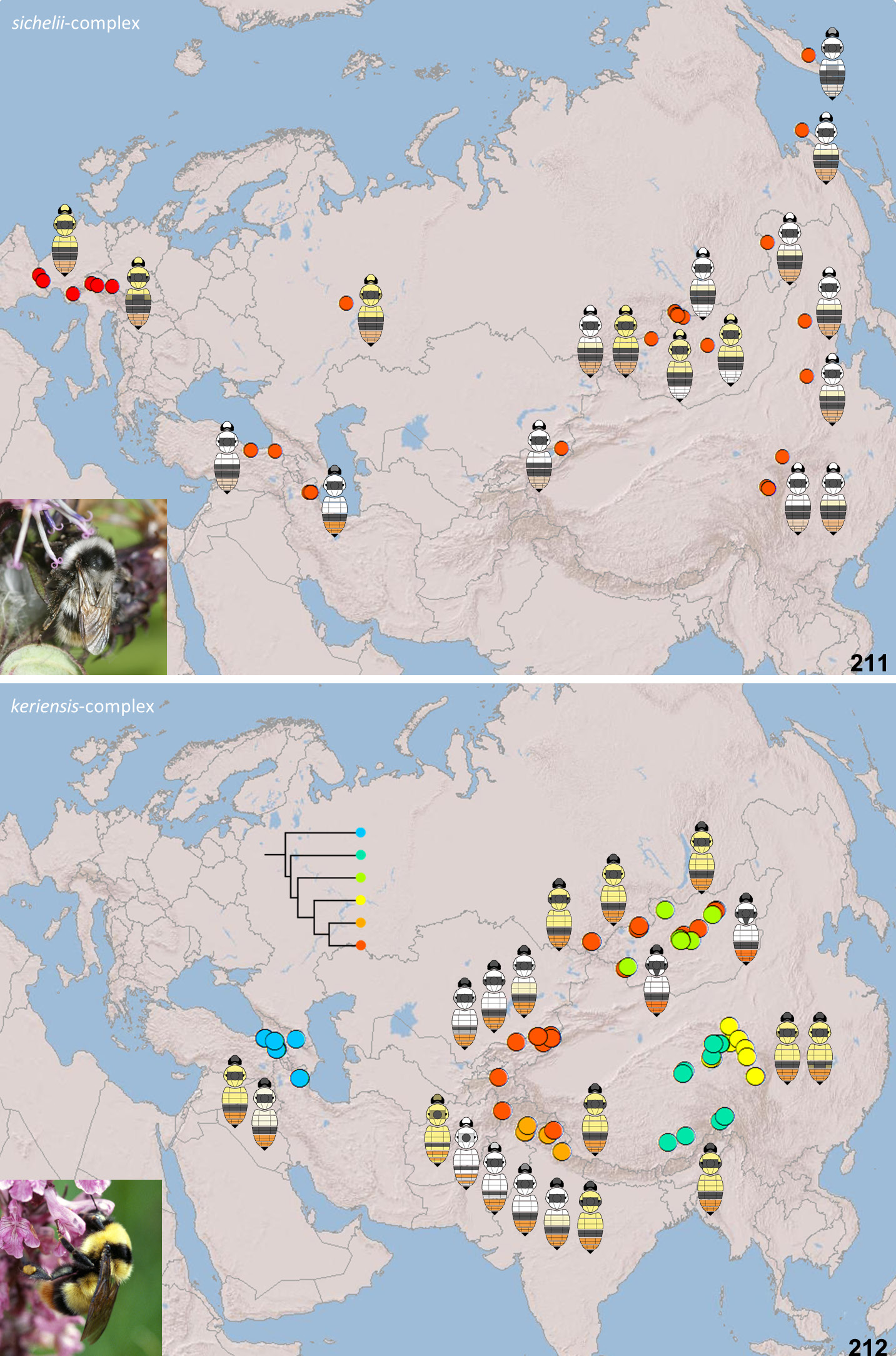
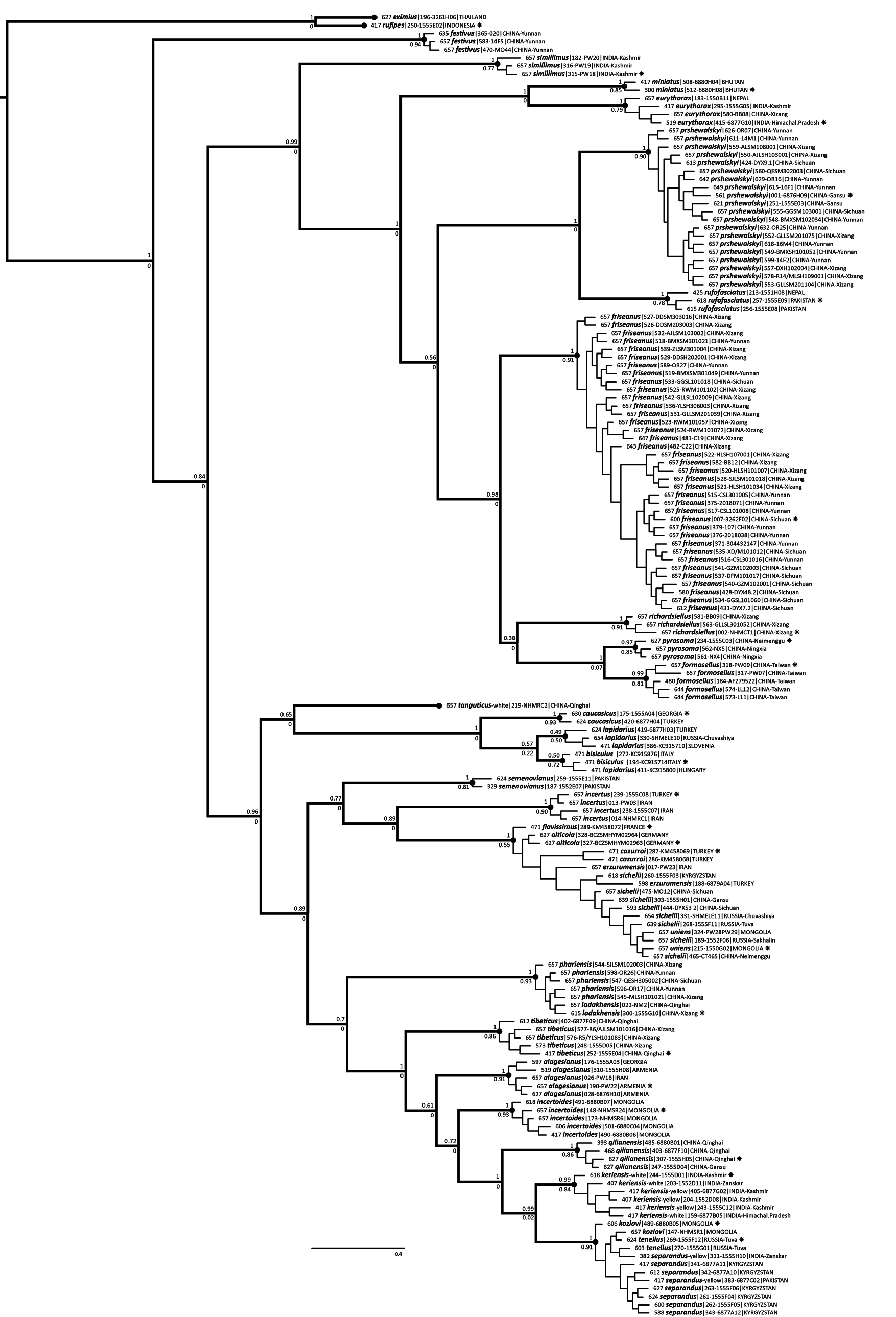
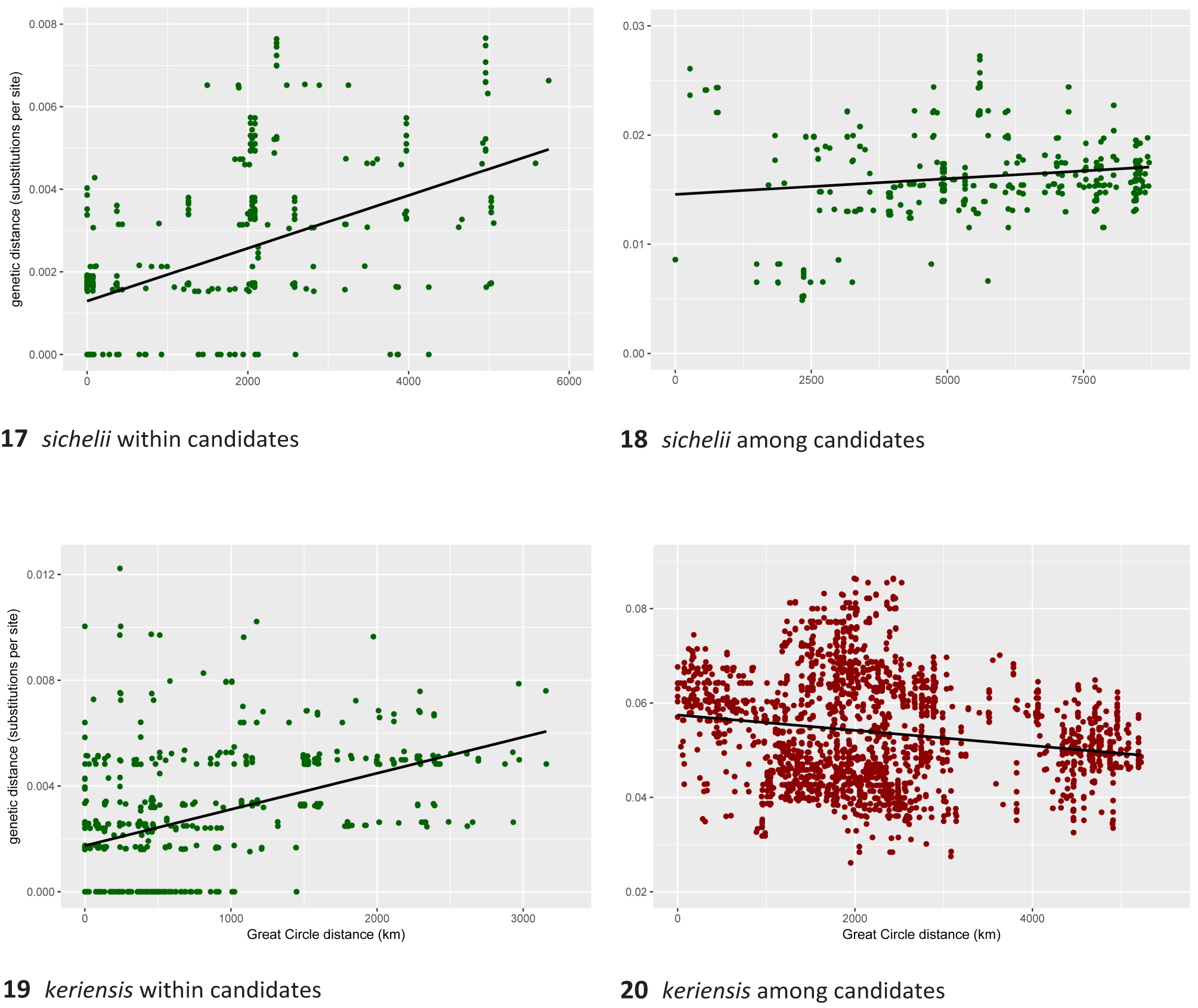
Our PTP analysis ( Fig. 10 View Fig ) of coalescents in the COI gene within the keriensis -complex supports six species including B. keriensis s. str., corroborated by differences in morphology. These species are also supported by the absence of a positive divergence-with-distance relationship among them ( Fig. 20 View Figs 17–20 ) (see Divergence and geographical distance, page 12).
From morphology, after examining>1000 specimens of the keriensis -group from across Asia and after comparing the distribution of states of several characters as part of this study (PW), Vogt’s character state of at least a few yellow hairs on the face does appear to remain one of the most consistently diagnostic character states for B. keriensis s. str. (by comparison with COI sequences), at least for queens in most parts of the species’ range (but see the comments on B. separandus regarding the Mongolian taxon kozlovi ). Several aspects of the colour pattern vary strongly within the species: (1) B. keriensis s. str. includes both yellow-banded and white-banded colour patterns; (2) the extent of pale hair on the face varies; (3) the extent of the black hair between the wing bases varies; (4) the extent of the pale hair on the side and ventral area (leg bases) of the thorax varies; (5) the extent of the pale hair posteriorly on T3 varies (the ‘ciliation’ described by authors); (6) the contrast of the pale posterior fringes on T4–5 varies. Many of these characters might appear to show a continuum of variation that is also continuous with the variation within B. separandus ( Williams 1991) , which might then appear consistent with the two taxa being parts of the same species ( Reinig 1935). However, evidence from COI barcodes shows that the two gene-coalescent groups differ most consistently in the presence of pale hairs on the face of B. keriensis s. str., supporting Vogt’s characterisation of the species, and providing integrated morphological support for its status as a separate species. The overlapping colour variation of B. separandus but only in Mongolia (where B. keriensis s. str. does not occur) does not negate the fact that B. keriensis s. str. is recognisable by morphological characters throughout its geographical range.
There are some yellow-banded queens from the Pir Panjal and Great Himalaya ranges (part of B. keriensis s. str. according our COI coalescent results, Fig. 10 View Fig ) that have very few pale hairs on the face, although these specimens still have a few yellow hairs on the leg bases and have a few black hairs in the pale bands of the thoracic dorsum. Many of the available specimens have failed repeated attempts at sequencing, so the species’ diagnosis remains tentative. Particularly influential in the interpretation of these individuals is one sequenced yellow-banded queen with very few pale hairs on the face, on the underside of the thorax, and on T3, from the Pir Panjal range (PW: ML405, Figs 170 View Figs 139–180 , 209 View Figs 209–210 ) that groups with B. keriensis s. str. ( Fig. 10 View Fig ) rather than with B. separandus (diagnostic base positions 267T, 286T, 301T, 306C, 339C, 363T, 423C, 498T, 529A, 540C, 541T, 556T). This is one of the specimens that most closely resembles some yellow-banded B. separandus also from Kashmir ( Figs 177 View Figs 139–180 , 210 View Figs 209–210 ) (the difference in the breadth of the black band between the wings is not diagnostic for these species).
Diagnosis
Females
Queens medium-sized body length 17–21 mm, workers 9–14 mm. Can be distinguished in Central Asia by the combination of hair of the face usually with some pale hairs, the leg bases usually with some pale hairs, T2 posteriorly entirely pale without black hairs, with T3 laterally usually with some pale hairs (cf. B. sichelii , B. separandus ).
Males
Body length 11–15 mm. Can be distinguished reliably at present only by their COI sequence, although in the western Himalaya they may be distinguished by yellow hair between the wing bases and on T3 posterio-laterally. Genitalia ( Fig. 203 View Figs 199‒204 ) with the gonostylus shorter than broad, its inner basal projection reduced to a short stub (cf. rufipes- group, festivus- group, rufofasciatus -group); volsella with the inner distal corner broadly produced but without a narrow hook (cf. rufipes- group, festivus- group, rufofasciatus -group); eye unenlarged relative to female eye.
Material examined
Lectotype designation
CHINA • ♀ (queen), lectotype of Bombus keriensis Morawitz, 1887 ; Keria Mts ; 9000 ft a.s.l.; Przewalsky leg.; ZIN.
Morawitz’s original description of several females of B. keriensis gives the type locality as “Keria” [= Keriya/Yutian, east of Hotan /Hetian / Khotan], presumably not in the low-lying town but higher in the neighbouring Karakax Shan (of the western Kunlun Shan [shan = mountains], southern Xinjiang, and probably near the village of Polu / Pulu/ Pulucun [cun = village], 36.1850° N, 81.4827° E. This village was visited by N. Prshewalsky while searching for a pass southwards into Ladakh during his fourth (and final) Central Asian expedition in the summer of 1885 ( Rayfield 1976). If the “9000” on the syntype specimen label (below) were an elevation in feet then this is substantially higher than the elevation of Keriya (ca 4500 feet), so a collecting site near Pulu is highly likely. These mountains are very arid, so likely sites with a reliable water supply and food plants every summer sufficient to support bumblebee colonies are restricted ( Williams et al. 2015a).
The ZIN collection contains a queen that agrees with the original description and carries seven labels: (1)gold disc;(2)handwritten white label“Keria / Gebirg./ 9000”reverse“2/10/[illegible]”;(3)handwritten white label “Przewalsky”; (4) handwritten white label “ Bombus / keriensis ♀. / F. Morawitz.”; (5) printed white label in Cyrillic “[K. F. Morawitz]”; (6) red label partly printed in black “ Lectotypus Bombus / keriensis F. Morawitz / design. Podbolotsk.” (unpublished); (7) red label printed in black “ LECTOTYPE ♀ / Bombus / keriensis / Morawitz, 1887 / det. PH Williams 2017”. This specimen, which is complete, is recognised as one of Morawitz’s syntypes and is designated here as the lectotype in order to reduce uncertainty in the identity and application of the name.
Material sequenced ( 7 specimens)
INDIA – Kashmir • 1 ♀ (queen); Gulmarg, Mt Apharwat ; 34.0548° N, 74.3856° E; Aug. 1985; P. Williams leg.; BOLD seq: 6877G02; PW: ML405 GoogleMaps • 1 ♀ (queen); Gulmarg, Mt Apharwat ; 34.0212° N, 74.3208° E; 21 Jul. 2009; BOLD seq: 1552D08; RR: ML204 GoogleMaps • 1 ♀ (queen); Zanskar, Padum ; 33.4746° N, 76.8871° E; 30 Jul. 2007; BOLD seq: 1552D11; RR: ML203 GoogleMaps • 1 ♀ (worker); same collection data as for preceding; 31 Jul. 2011; R. Raina leg.; BOLD seq: 1555C12; RR: ML243 GoogleMaps • 1 ♀ (worker); Razdaan pass; 34.5390° N, 74.6377° E; 23 Jul. 2012; R. Raina leg.; BOLD seq: 1555D01; RR: ML244 GoogleMaps • 1 ♀ (worker); Dawar ; 34.6464° N, 74.7171° E; 24 Jul. 2012; R. Raina leg.; BOLD seq: 1555H03; RR: ML305 . — Himachal Pradesh GoogleMaps • 1 ♀ (worker); Saichu Nulla ; 31.6° N, 78.267° E; 8 Sep. 1986; A. Hutchings leg.; BOLD seq: 6877B05; PW: ML159 GoogleMaps .
Global distribution
(West Qinghai-Tibetan-Plateau species) Himalaya: AFGHANISTAN, PAKISTAN, INDIA: Kashmir, Himachal Pradesh. – East Asia: CHINA: Xizang. (IOZ, NHMUK, PW, RR, ZIN, ZSM.) The species is usually not common ( Fig. 212 View Figs 211–212 ).
Behaviour
Food-plant generalists ( Williams 1991). Male mate-searching patrolling behaviour ( Williams 1991).



| ZIN |
Russian Academy of Sciences, Zoological Institute, Zoological Museum |
| PW |
Paleontological Collections |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
|
SubGenus |
Melanobombus |
Bombus keriensis Morawitz, 1887
| Williams, Paul H., Altanchimeg, Dorjsuren, Byvaltsev, Alexandr, Jonghe, Roland De, Jaffar, Saleem, Japoshvili, George, Kahono, Sih, Liang, Huan, Mei, Maurizio, Monfared, Alireza, Nidup, Tshering, Raina, Rifat, Ren, Zongxin, Thanoosing, Chawatat, Zhao, Yanhui & Orr, Michael C. 2020 |
Pyrobombus keriensis subsp. karakorumensis Tkalců, 1989: 57
| Tkalcu B. 1989: 57 |
Bombus trilineatus
| Wang S. - F. 1982: 441 |
Lapidariobombus separandus subsp. meridialis
| Skorikov A. S. 1914: 127 |
Bombus keriensis Morawitz, 1887: 199
| Morawitz F. F. 1887: 199 |