
Afronycteris, 2020
|
publication ID |
https://doi.org/ 10.1093/zoolinnean/zlaa087 |
|
publication LSID |
lsid:zoobank.org:pub:71737F08-2938-4403-8385-5438B2E5EABE |
|
DOI |
https://doi.org/10.5281/zenodo.4451456 |
|
persistent identifier |
https://treatment.plazi.org/id/25458781-FFAA-E413-533C-FF34FF1F52C3 |
|
treatment provided by |
Admin |
|
scientific name |
Afronycteris |
| status |
gen. nov. |
AFRONYCTERIS MONADJEM, PATTERSON & DEMOS View in CoL GEN. NOV.
LSID: http://zoobank.org/ urn:lsid:zoobank.org:pub:71737F08-2938-4403-8385-5438B2E5EABE
Synonymy
Vespertilio Peters, 1852 View in CoL (part, not Linnaeus, 1758).
Hypsugo Kolenati, 1860 View in CoL (part, not Kolenati, 1856).
Vesperugo Dobson, 1875 (part, not Keyserling & Blasius, 1839).
Pipistrellus Miller, 1900 View in CoL (part, not Kaup, 1829).
Myotis Matschie, 1907 View in CoL (part, not Kaup, 1829).
Neoromicia Shortridge, 1934 View in CoL (part, not Roberts, 1926).
Eptesicops Roberts, 1951 (part, not Roberts, 1926).
Complete synonymic histories for the species placed herein in Afronycteris are given in the African Chiroptera report ( AfricanBats NPC, 2019).
as Neo. tenuipinnis ). However, its distribution beyond western Kenya is not known. It seems to be associated with the high plateau of western Kenya, which extends into eastern Uganda; presumably, it also occurs there. Thorn & Kerbis Peterhans (2009) recorded ‘ Pipistrellus tenuipinnis ’ as occurring widely in Uganda. The cranial measurements of specimens from Budongo, Entebbe and Sango Bay (at elevations similar to those we report from Kenya) all fall neatly within the range of Pse. nyanza and are generally larger than those for Pse. tenuipinnis . It would be instructive to re-examine these specimens (in the collections of the BMNH and LACM) to confirm their identities and help to determine the western limits of the distribution of Pse. nyanza . However, records from the eastern Democratic Republic of the Congo apparently refer to true Pse. tenuipinnis , owing to their small size, with total length ‘about 72 mm’ ( Allen et al., 1917). We speculate that, despite the limited geographical range of Pse. nyanza (even if Uganda is included), this species is currently not threatened because it survives in human-altered habitats, and therefore we recommend the IUCN conservation status of ‘Least Concern’. The type specimen echolocated at a peak frequency (start and end frequencies) of 40.4 kHz (56.4–39.3 kHz). The mean (± SD) peak frequency for 16 individuals at the type locality was 40.4 ± 0.84 kHz (55.1 ± 7.91 to 39.5 ± 0.68 kHz).
Type species: Afronycteris nana (Peters, 1852) View in CoL .
Included species: Afronycteris helios (Heller, 1912) .
Etymology: From the Greek word νυχτερίδα, bat, and the prefix Afro- referring to the African continent, referring to the wide distribution of the type species A. nana . This species ranges, without obvious breaks in distribution, from Senegal in the west, east to Ethiopia and south to South Africa, being absent only from the more arid desert and semi-desert environments associated with the Sahara, Sahel and Chalbi Desert in the north and the Namib and Kalahari deserts in the south-west ( Happold, 2013a).
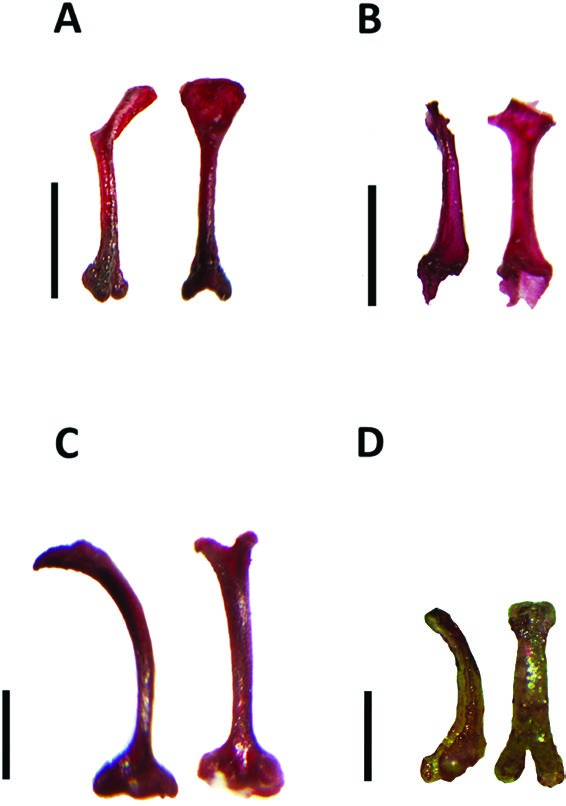
Diagnosis: Small-sized vespertilionids with the simple muzzle characteristic of this family. The cranium in lateral view is distinctly inflated, more so than any other member of the tribe Vespertilionini . The tragus is characteristically hatchet shaped, with the posterior margin having an abrupt angle and lacking a notch at its base, as illustrated by Van Cakenberghe & Happold (2013). The tragi of Laephotis , Neoromicia and Pseudoromicia all have a notch at the base of the posterior margin. The pelage of the upper and under parts is bicoloured, with the basal portion of each hair darker than the terminal portion. There is a distinct thumbpad at the base of the thumb, thought to be useful in climbing on smooth leaves. The outer incisor I 2 is well developed, reaching almost the same length as the I 1, the latter being slightly bicuspid or unicuspid; in Laephotis , Neoromicia and Pseudoromicia , I 2 is typically half the length of I 1 or shorter. The P 1 is present and relatively large, whereas this tooth is absent in Laephotis , Neoromicia (except Neo. bemainty and Neo. anchietae ) and Pseudoromicia . The baculum (~2.0 mm in length) is shorter than in Pseudoromicia and similar in length to that of Laephotis and Neoromicia . It has a distinctly and deeply bilobed base and a gently curved shaft leading to a spatulate tip ( Fig. 5D View Figure 5 ).
Distribution: This genus is endemic to sub-Saharan Africa, probably occurring in suitable habitats across its wide range. It occurs throughout the Upper Guinea rainforest zone, extending northward into Sudanian savanna, possibly extending into the Sahel along major rivers and wetlands ( Happold, 2013a). It occurs throughout mesic portions of Central and East Africa, but records are sparser in the Horn of Africa ( Lanza et al., 2015). It is widespread in the wetter parts of southern Africa, avoiding the dry southwestern region of South Africa, much of Botswana and Namibia ( Monadjem et al., 2010).
Systematic relationships: Afronycteris is sister to Pseudoromicia , but the two genera can be distinguished easily by external characteristics, cranial features and the shape of the baculum (see ‘Diagnosis’ above for details).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Afronycteris
| Monadjem, Ara, Demos, Terrence C, Dalton, Desire L, Webala, Paul W, Musila, Simon, Kerbis Peterhans, Julian C & Patterson, Bruce D 2020 |
Eptesicops
| Roberts 1951 |
Neoromicia
| Shortridge 1934 |
Myotis
| Matschie 1907 |
Pipistrellus
| Miller 1900 |
Vesperugo
| Dobson 1875 |
Hypsugo
| Kolenati 1860 |
Vespertilio
| Peters 1852 |