
Bulla, LINNAEUS, 1758
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2008.00369.x |
|
persistent identifier |
https://treatment.plazi.org/id/255D87DA-FF9E-FFC3-895A-176BFD45D15F |
|
treatment provided by |
Felipe |
|
scientific name |
Bulla |
| status |
|
GENUS BULLA LINNAEUS, 1758 View in CoL
Bulla Linnaeus, 1758: 725 View in CoL View Cited Treatment (type by subsequent designation, Gray, 1847; Bulla ampulla Linnaeus, 1758 View in CoL ). Nux View in CoL da Costa, 1776: 173 (non binominal).
Bullus Montfort, 1810: 331 (type by monotypy Bulla ampulla View in CoL ).
Bullaria Rafinesque, 1815: 142 View in CoL (type species not designated).
Bullea Blainville, 1825: 478 View in CoL (type by monotypy Bulla ampulla View in CoL ).
Vesica Swainson, 1840: 360 View in CoL (type here designated Bulla ampulla View in CoL ).
Quibulla Iredale, 1929: 349 View in CoL (type by original designation Bulla botanica Hedley, 1918 View in CoL ).
The genus Bulla View in CoL has a complex taxonomic history, and there has been persistent controversy about the definition of the genus and designation of a type species. The name Bulla View in CoL was introduced by Linnaeus (1758) for a genus of 23 species that are now classified in diverse genera belonging to families of marine, land and freshwater molluscs. Of these original species, only B. ampulla View in CoL remains today in the genus. Among the other species, three are impossible to assign to known taxa and the remainder are distributed among 18 genera ( Dodge, 1955).
Linnaeus (1758: 725) provided only a brief and vague definition of the genus Bulla . Furthermore, he gave this same name to a subgroup of orthopteran insects of the genus Gryllus ( Linnaeus, 1758: 427) . This gave rise to some confusion, as authors claimed that the name Bulla was preoccupied by its use as a subgenus of insects ( Dodge, 1955). This was finally settled by the ruling of the International Commission of Zoological Nomenclature that the subdivisions of genera published by Linnaeus do not have subgeneric status (Anon., 1936: Opinion 124).
The first modification of Linnaeus’ concept of Bulla was suggested by da Costa (1776). He referred to the complexity of the genus and redefined the group to exclude ovulids, leaving only species bearing, among other features, an umbilicate oval shell. da Costa designated this group by the name Nux , and his figured example undoubtedly belongs to the genus Bulla ( da Costa, 1776: pl. 3, figs 4, 5). da Costa did not employ any specific names in this work, where he seems to have followed a uninominal nomenclature.
Bruguière (1792) also acknowledged that Linnaeus had united several unrelated groups in Bulla , and introduced more fundamental changes to the concept of the genus. Bruguière likewise excluded all the Ovulidae , and restricted the genus to marine species. According to this author only species with an aperture the full length of the shell and spire hidden inside the shell or in the top of it should be included in the genus Bulla . Differences in the spire were used to split the genus into two (unnamed) groups. In the first group he included species now recognized as Cylichna and Bulla in the modern sense, whereas the other group was a repository for diverse species belonging to genera such as Atys , Haminoea , Philine , Retusa , Volvulella , Hydatina , Akera , Scaphander , Aplustrum and Micromelo .
The concept of Bruguière (1792) was slightly refined by Lamarck (1801: 90; 1822: 27), who also emphasized the weaknesses of Linneaus’ definition of Bulla . Lamarck admitted only species with an external, globose and involute shell, removing for example Philine to a different genus ( Bullaea ), and leaving only six of the original 23 Linnaean species ( Lamarck, 1822). Lamarck was also the first to describe the external morphology of the animals. By this time the genus was so modified from its original Linnean concept that Gray (1847) attributed the genus to Lamarck (1801). Gray also designated Bulla ampulla as the type species.
de Montfort (1810) further narrowed the concept of Bulla , although under a different genus name ( Bullus ), but again took B. ampulla as the type species. He included a brief diagnosis and one figure of B. ampulla and introduced the generic names Scaphander and Atys for B. lignarius and B. naucum , until then included in Bulla .
Further complexity was introduced by Rafinesque (1815) who proposed the name Bullaria and considered Bulla its synonym. He defined this group in vague terms as bearing a dorsal shell, never conic, with a slightly rounded or spiralled base, which does not clearly match the features of Bulla . This name was ignored for a long time, but resurrected by Suter (1913) and Hedley (1916, 1918) for the Australian species B. vernicosa (as B. adamsi ) and B. quoyii (as B. australis , B. botanica and B. tenuissima ), and has also been used for the Atlantic species B. striata ( Fischer-Piette, 1942; Nicklès, 1950; Marche-Marchad, 1958; Nordsieck, 1972) and B. occidentalis ( Peile, 1926) , and for the eastern Pacific species B. gouldiana and B. punctulata ( Baker & Hanna, 1927) .
de Blainville (1825) employed the name Bulla for Haminoea hydatis , whereas Bulla ampulla was assigned to a new genus Bullea , together with Philine aperta (Linnaeus, 1767) and others. The situation became even more confused after Swainson (1840) once more ascribed Scaphander lignarius to Bulla and created the new genus Vesica for Bulla ampulla and Atys naucum ( Linnaeus, 1758) .
The major contributions that stabilized the concept of the genus were those of Adams (1850), Sowerby (1868) and finally Pilsbry (1895), to whom the modern concept of the genus should be attributed. Since then, non- Bulla species have hardly ever been included in the genus.
Nevertheless, Iredale (1929) claimed that the confusion surrounding the definition of this group and choice of a type species was such that only a new name could settle the issue. He therefore introduced Quibulla , with type species Bulla botanica Hedley, 1918 . However, this proposal was not widely accepted and only a few works dealing with Australian species have used the name Quibulla , and even then with uncertainty (e.g. Rudman, 1971).
Since Linnaeus (1758: 726) in his lists of synonyms only quoted the name Bulla once, to refer to the species Bulla naucum (presently known as Atys naucum ) as used by Rumphius (1705), B. naucum should therefore be regarded as the type species of Bulla by Linnaean tautonomy (ICZN, 1999: art. 68.5). If this argument were to be followed, this name would not be available for the Bulla ampulla group to which it has traditionally been applied, and Atys Montfort, 1810 would become a synonym of Bulla Linnaeus, 1758 . To avoid this profound nomenclatural change the International Commission of Zoological Nomenclature ruled that Bulla ampulla Linnaeus, 1758 should be designated as the type species of the genus Bulla and placed both names on the Official List of Generic and Specific Valid Names in Zoology (ICZN, 1954: Opinion 196).
During the second half of the 20th century the name Bulla became established in the literature (e.g. Habe, 1950; Willan, 1977, 1978; Wells, 1985; Fukuda, 1995; Hori, 2000; Malaquias, Martínez & Abreu, 2002; Valdés & Camacho-García, 2004; Ekawa & Sakashita, 2005), and other generic names such as Bullaria and Quibulla were seldom used (e.g. Rudman, 1971; Nordsieck, 1972). Only Nordsieck & García-Talavera (1979) included several species of Bulla in the genus Cylindrobulla ; this, however, is a genus of sacoglossan opisthobranchs.
Accounts of anatomical features of the genus Bulla can be found in Vayssière (1885: general anatomy of B. striata ; 1906: radula, gizzard and male genital system of B. arabica , as B. ampulla ), Pilsbry (1895: general anatomy of Bulla ), Bergh (1900: jaws, gizzard plates and spines, and male genital system of B. ampulla ; gizzard plates of B. gouldiana , as B. adamsi ; radula of B. arabica , as B. ampulla var. M. rubri ; jaws, radula, gizzard plates and spines of B. occidentalis , as B. media ; gizzard plates and spines of B. striata , as B. media var. mediterr .), Si (1931: general anatomy of B. striata ), Eales (1938: central nervous system of B. arabica , as B. ampulla ), Risbec (1951: general anatomy of B. ampulla ), Pruvot-Fol (1954: general anatomy of B. striata , radula and gizzard), Marcus (1955: radula and gizzard of B. occidentalis , as B. striata ; 1957: general anatomy of B. occidentalis , as B. striata ; 1961: radula and male genital system of B. gouldiana ), Rudman (1971: general anatomy of B. quoyii , as Quibulla ), Robles (1975: reproductive anatomy of B. gouldiana ), Willan (1977: radula of B. quoyii ), Mikkelsen (1996, 2002: general anatomy of Bulla ), deMaintenon & Mikkelsen (2001: reproductive anatomy of B. occidentalis , as B. striata ), Malaquias et al. (2002: radula and male genital system of B. mabillei ), Valdés & Camacho-García (2004: radula and gizzard of B. punctulata ), Ekawa & Sakashita (2005: radula and gizzard of B. ampulla ), Malaquias & Reid (2008: female glands of B. striata ).
Diagnosis: Shell ovoid, solid, mottled in drab colours. Spire involute. Aperture slightly longer than shell height. Cephalic shield produced antero-laterally, forming pair of siphon-like tentacles. Penis invaginable, formed by a long duct continuous with coiled prostate, ending in a narrow terminal blind caecum; all structures enclosed in thick muscular sheath, contiguous with a long tubular vestibular area; retractor muscle attached to proximal area of penial sheath. Anterior gonoduct slightly helicoid; fertilization chamber rounded, located at mid-length; seminal receptacle placed antero-ventrally, attached to wall of common genital duct. Rachidian tooth broad, denticulate, with central cusp usually smaller. Large gap between rachidian and laterals. Inner lateral clawshaped, usually symmetrical with major central cusp and 3–4 lateral cusps on either side. Outer lateral with smooth vertical internal edge and denticulate convex outer edge with 4–10 cusps; base with denticle. Second outer lateral teeth reduced, plate-like. Gizzard plates uncalcified, smooth, with longitudinal keel.
Shell: ( Figs 1A, C View Figure 1 , 2 View Figure 2 , 5 View Figure 5 ) External, solid, ovoid to slender, smooth or with few spiral striae, mottled in drab colours. Columella smooth and thick. Parietal callus present. Spire involute, ornamented with concentric striae, with or without pronounced sutures. Visible area of protoconch convex and smooth. Aperture inverted comma-like shape, slightly longer than shell height.
External body: ( Fig. 1C View Figure 1 ) Cephalic shield produced antero-laterally to form a pair of siphon-like tentacles, and posteriorly into a pair of cephalic lobes covering anterior part of shell. Eyes visible through skin, periocular area unpigmented. Pair of Hancock organs located laterally on either side of head, underneath cephalic shield; wrinkled or perfoliate with lamellae; surface with cilia. Parapodial lobes small, folded up laterally. Foot shorter than shell, comple- mented by posterior pallial lobe modified into an accessory foot protruding slightly behind shell. Animal can retract completely into shell. Operculum absent in adults.
Reproductive system: ( Figs 13 View Figure 13 , 14 View Figure 14 ) Monaulic and separated into two parts: male copulatory system in cephalic region and hermaphroditic gonoduct in posterior part of body; both parts interconnected through external ciliated seminal groove that lies dorsolaterally on right side of body. Male copulatory system: invaginable penis with smooth tip, formed by long duct, sometimes coiled, continuous with prostate that is tightly coiled into a ball, ending as a narrow terminal blind caecum. Penis, prostate and caecum enclosed in thick muscular sheath, contiguous with long tubular vestibular area that opens in male genital aperture into external seminal groove positioned in front of right Hancock’s organ. Retractor muscle attached to proximal area of penial sheath and to body wall close to male genital aperture.
Hermaphroditic gonoduct formed of two parts, the anterior and posterior gonoducts (sensu deMaintenon & Mikkelsen, 2001; see Malaquias & Reid, 2008, for detailed account and discussion of terminology). Posterior gonoduct consists of the hermaphroditic gonad and its immediate coiled duct (ampulla hermaphroditica) up to the sphincter. Anterior gonoduct extends from sphincter to common genital opening on right side of animal; shape elongate or slightly helicoid; formed by part of ampullar duct, female gland mass (encompassing albumen gland, fertilization chamber with an embedded coiled duct here designated as the membrane gland, and three mucous glands), vestibular area, common genital duct, seminal receptacle, and gametolytic duct and gland.
The fertilization chamber occurs centrally in the albumen–membrane gland complex, and is a crossroads where several ducts end and others begin; its surface shows either a complex structure of ducts or simply a few loops formed by wider ducts. The ampullar duct (post-ampullar duct sensu Robles, 1975) connects with the area dorso-laterally on the left side, and seems to open in the area of the fertilization chamber. The membrane gland (capsule gland sensu deMaintenon & Mikkelsen, 2001) is a coiled duct embedded in the fertilization chamber that runs either dorsally along the common genital duct, or within it, towards the common genital aperture. Here it makes a half-loop just behind the vestibular area, continuing backwards firstly in a dorsal position, adjacent to the right lobe of the mucous gland, then disappears underneath the fertilization chamber and reappears dorsally behind it on the right side of the albumen gland, where it merges with the left lobe of the mucous glands. The left lobe continues posteriorly to the tip where it is continuous with the posterior right lobe of the mucous gland that extends the full length to the genital aperture.
The common genital duct is a muscular structure continuous with the fertilization chamber, which leads to the vestibular area behind the common genital aperture.
The seminal receptacle (spermatocyst sensu Marcus, 1957; exosperm storage pouch sensu deMaintenon & Mikkelsen, 2001) is a small outpocket located ventrally and contained within the walls of the common genital duct. The gametolytic gland (spermatheca sensu Marcus, 1957; bursa copulatrix sensu Robles, 1975) is positioned adjacent to the pericardium in the pallial cavity roof, and connects ventrally through a duct into the common genital duct and seminal receptacle.
Jaws: ( Fig. 6 View Figure 6 ) Embedded laterally in buccal mass close to ciliated oral veil; crescentic or elongate, symmetrical, composed of long compacted hexagonal columns, producing scale-like sculpture at surface.
Radula: ( Figs 8–10 View Figure 8 View Figure 9 View Figure 10 ) Radular formula 1.2.1.2.1 (first 3–8 rows with rachidian teeth only). Rachidian tooth broad, denticulate, central cusp usually smaller, 4–9 cusps on either side. Large gap between rachidian and laterals, nearly half-width of rachidian. Inner lateral tooth claw-shaped, usually symmetrical, major central cusp and 3–4 lateral cusps on either side. Outer lateral tooth with smooth vertical internal edge; denticulate convex outer edge with 4–10 robust cusps, innermost usually most developed; base denticulate. Second outer lateral tooth plate-like, fragile, not denticulate.
Gizzard: ( Fig. 11 View Figure 11 ) Located between oesophagus and stomach, with three uncalcified, corneous (chitinous sensu Rudman, 1971), smooth, brownish plates, identical in shape, usually with longitudinal keel. Gizzard spines arranged in anterior and posterior rings at entrance and exit of gizzard; spines are corneous with basal papillae. Anterior spines sparse (about ten), larger, usually laterally flattened. Posterior spines needle-like, numerous, densely arranged in 2–4 rings.
Central nervous system: ( Fig. 1B View Figure 1 ) Euthyneuran. Circumoesophageal nerve ring prepharyngeal. Cerebral, pleural and pedal ganglia separated, equidistant, connected by short connectives. Pedal ganglia larger than others, connected to each other by long commissure that runs ventrally to buccal mass. Pair of cerebral ganglia connected by long commissure dorsal to buccal mass. Right pleural ganglion smaller than left, contiguous with right parietal ganglion of similar size. Left parietal ganglia very small, almost imperceptible, located at half length in left nerve cord. Larger supraoesophageal ganglion arising from nerve cord on posterior right side. Smaller suboesophageal ganglion in left nerve cord, contiguous with larger visceral ganglion. Genital ganglion on posterior left side, located behind visceral ganglion where both nerve cords merge.
ATLANTIC SPECIES THE BULLA STRIATA GROUP
Considerable confusion surrounds what Pilsbry (1895) first designated as the ‘Group of B. striata ’ in the Atlantic Ocean. He observed that ‘The littoral Bullas of this area form a very difficult assemblage requiring a great mass of material for its elucidation. The creatures themselves are mostly not thoroughly differentiated into “species”...’. Fourteen specific and seven varietal names have been applied to the members of this group, which is a reflection of their considerable variation in shell shape and colour.
The name B. striata was introduced by Bruguière (1792) for shells from the Mediterranean, and eastern and western Atlantic. He provided an accurate description and illustration of shells, although the origin of his material was not mentioned and the geographical range was based on earlier works.
Röding (1798) introduced the name B. umbilicata in a sale catalogue. This name was neglected for more than 100 years, and this catalogue was only considered valid for nomenclatural purposes after 1926 (ICZN, 1958: opinion 96). During the 20th century several authors (e.g. Rehder, 1962; Abbott, 1974; Redfern, 2001) resurrected the name for western Atlantic specimens, but the International Commission of Zoological Nomenclature suppressed it (ICZN, 1959: opinion 549), giving precedence to Bulla umbilicata Montagu, 1803 , today included in the genus Cylichnina .
Controversy also surrounds the identity of B. amygdalus , introduced by Dillwyn (1817) using one of Solander’s names (for an account of Solander manuscripts and their molluscan names see Wilkins, 1955). The molluscan volumes of Solander’s manuscripts are closely connected with Joseph Banks’ shell collection ( Wilkins, 1955). In volume 9, slip 140, the name B. amygdalus is mentioned, referring to shells from Jamaica, with a cross-reference to a lot in the Banks collection (now in NHM). This lot consists of seven specimens belonging to four different species of Bulla , of which two specimens belong to the B. striata group. These specimens could be the ones described by Solander as B. amygdalus . However, the crossreference in the manuscript is a pencil number apparently added later, and because not all these cross-references match with labels in the Banks collection it cannot be certain if this case with B. amygdalus refers to the specimens seen by Solander or is merely a fortunate coincidence. Unfortunately, Dillwyn (1817) did not refer to Solander’s specimens, or to the geographical information provided in his manuscript, and considered B. amygdalus to occur on both sides of the Atlantic. Later Adams (1850) mistakenly attributed the authorship of B. amygdalus to Lister (1685 –92). Both Adams (1850) and Pilsbry (1895) regarded this as an exclusively western Atlantic species, and since then the name has been employed sporadically for material from both sides of the Atlantic ( Nordsieck, 1972; Nordsieck & García- Talavera, 1979; Macedo et al., 1999). Poppe & Goto (1991) pointed out the continuing confusion between B. amygdalus and B. striata , considering that the former might be a variety of B. striata .
Later, delle Chiaje (1827) introduced the new name B. columnae for shells from Sicily. Despite the ambiguous description, the illustration is undoubtedly a specimen of the B. striata group. However, the name has hardly been used since; Pallary (1900) used it for specimens from Algeria.
A name that has been used with some consistency among authors is B. adansonii , introduced by Philippi (1847) for specimens from ‘Senegambia’. Philippi recognized minor differences in the West African shells, which he considered sufficient to create a new species. The fact that this name is associated with a geographical area with a distinctive fauna has contributed to its use for West African specimens. Nevertheless, similarities with other forms were also noted. For example Pilsbry (1895) wrote: ‘It is indeed difficult to distinguish some West African specimens from the Antillean B. amygdala ; but as the geographical ranges of the two are now so widely sundered, I consider it best to make the distinction between them here. The only alternative to this course would be to “lump” the whole striata group.’
Philippi (1847) named an additional species, B. media , from the Caribbean Sea. Apart from a few references during the second half of the 19th century (see synonymy of B. occidentalis ), this name has been neglected.
Adams (1850) introduced the name B. occidentalis for shells from St Vincent, Lesser Antilles. This name has been widely applied to the western Atlantic form; for example, Pilsbry (1895) considered it the most abundant of the West Indian Bulla and illustrated several shell variants.
A single unlocalized shell bearing the label ‘ B. zigzag Tryon Mss (nov. spec.)’ is housed in the type collection of ANSP. This shell belongs to the B. striata group, but the name was apparently never published.
›
Six additional specific names and another seven varietal names (see synonymies of B. striata and B. occidentalis ) have been proposed either for eastern or western Atlantic forms, in all cases based on small differences of shell shape and colour. The most prolific author was Menke (1853), who described 14 new species of Bulla , of which five refer unequivocally to the B. striata group.
The taxonomy of the B. striata group has been much disputed, and in the recent literature there is no agreement on the number of species. For example Marcus (1957), García-Talavera (1983), Abbott & Dance (1982) and Vermeij & Rosenberg (1993) all consider a single amphi-Atlantic species, whereas others (e.g. Nordsieck & García-Talavera, 1979; Bernard, 1984; Poppe & Goto, 1991; Macedo et al., 1999; Redfern, 2001) accept the occurrence of several species on each side of the Atlantic Ocean.
The study of material from the entire distribution range of the B. striata group, including shells and anatomy, together with a molecular phylogenetic hypothesis, has revealed the existence of two species, B. striata and B. occidentalis , in the eastern and western Atlantic, respectively. Nevertheless, only the molecular characters and geographical distribution clearly separate these species, whereas the high intraspecific and interpopulational variability of shells, and similarity of anatomy on both sides of the Atlantic, would likely have led to recognition of only a single species. Marcus (1957) had already pointed out that the small anatomical differences between specimens from both sides of the Atlantic appeared insufficient for specific separation.
BULLA STRIATA BRUGUIÈRE, 1792 View in CoL
( FIGS 2A View Figure 2 , 3A–L, 5A, G View Figure 5 , 6A, B View Figure 6 , 7 View Figure 7 , 9A–C View Figure 9 , 10A–D View Figure 10 , 11A–G View Figure 11 , 13A–F View Figure 13 , 14A–D View Figure 14 , 15 View Figure 15 )
Bulla striata Bruguière, 1792: 372 View in CoL , pl. 358, fig. 2a, b (Mediterranean Sea, West Africa and Caribbean Sea; neotype here designated BMNH 20050687, H = 28.5 mm, Fuzeta, Ria Formosa, Algarve, Portugal, Fig. 3A herein). Lamarck, 1822: 33 (in part, includes B. occidentalis View in CoL ). d’Orbigny, 1841: 122 (in part, includes B. occidentalis View in CoL ). Smith, 1890: 296 (in part, includes B. occidentalis View in CoL ). Pilsbry, 1895: 332, pl. 37, figs 42–46 (in part, includes B. occidentalis View in CoL ). Mabille, 1896: 116. Si, 1931: 29. Nobre, 1932: 57. Pasteur-Humbert, 1962: 175. Nordsieck, 1972: 27, pl. 4, fig. 13 (in part, includes B. occidentalis View in CoL ). Marcus, 1976: 125 (in part, includes B. occidentalis View in CoL ). Marcus, 1977: 4 (in part, includes B. occidentalis View in CoL ). Eisenberg, 1981: 153, pl. 135 (in part, includes B. occidentalis View in CoL ). Abbott & Dance, 1982: 279, figured (in part, includes B. occidentalis View in CoL ). von Cosel, 1982b: 21. von Cosel, 1982c: 61. Templado, 1982: 249. García-Talavera, 1983: 34 (in part, includes B. occidentalis View in CoL ). Luque, 1983: 55. Barash & Danin, 1989: 250. Poppe & Goto, 1991: 195, pl. 37, fig. 25. Barash & Danin, 1992: 186, fig. 198. Fernandes & Rolán, 1993: 41. Rios, 1994: 197 (in part, includes B. occidentalis View in CoL ). Murillo, 1996: 36. Macedo et al., 1999: 255, figured. Rolán & Ryall, 1999: 58. Guerreiro & Reiner, 2000: 211, figured. Malaquias & Morenito, 2000: 119. Costa et al., 2002: 128. Rolán, 2005: 203, figs 934, 935. Malaquias & Reid, 2008.
Bullaria striata – Nordsieck & García-Talavera, 1979: 173, pl. 43, fig. 25.
Bulla amygdalus Solander View in CoL in Dillwyn, 1817: 480 ( Jamaica, Barbados, Senegal, Nigeria; in part, includes B. occidentalis View in CoL ; possible types seen, 2 sh, BMNH Banks coll., H = 21.6, 27.6 mm; nomen dubium).
Bulla amygdala View in CoL – Menke, 1854b: 44. Nordsieck, 1972: 28, pl. 4, fig. 14 (in part, includes B. occidentalis View in CoL ). Poppe & Goto, 1991: 195, pl. 37, figs 26, 27. Macedo et al., 1999: 255, figured.
Cylindrobulla amygdala – Nordsieck & Garcia- Talavera, 1979: 174, pl. 43, fig. 28.
Bulla columnae View in CoL delle Chiaje, 1827: 24, pl. 46, figs 17– 11 (? Italy; types not seen) .
Bulla adansonii Philippi, 1847: 121 View in CoL (Senegambia [West Africa]; type untraceable). Adams, 1850: 576, pl. 123, fig. 69. Pilsbry, 1895: 333, pls 38, 48, figs 21, 61.
Bulla adansonii var. minor Dunker, 1853: 4 View in CoL , pl. 4, figs 11, 12.
Bulla adansoni View in CoL – Menke, 1854b: 44. Sowerby, 1868: sp. 13, pl. 5. Mabille, 1896: 118. Dautzenberg, 1910: 12. Nordsieck, 1972: 28, pl. IV, fig. 15. Bernard, 1984: 108, pl. 53, fig. 215. Rolán & Ryall, 1999: 58. Ardovini & Cossignani, 2004: 242.
Bullaria adansoni – Fischer-Piette, 1942: 128. Nicklès, 1950: 136, fig. 279. Marche-Marchad, 1958: 39.
Cylindrobulla adansoni – Nordsieck & García- Talavera, 1979: 173, pl. 43, fig. 26.
Bulla (Bullea) dactylis Menke, 1853: 137 View in CoL (Mediterranean Sea, Gibraltar, Atlantic to Angola; types probably lost, see Dance, 1986). Nordsieck, 1972: 28.
Bulla (Bullea) perdicinia Menke, 1853: 140 View in CoL (Atlantic, Guinea and Sierra Leone; types probably lost, see Dance, 1986). Pilsbry, 1895: 335.
Bulla (Bullea) omphalodes Menke, 1853: 137 View in CoL (Mediterranean Sea, southern Italy; types probably lost, see Dance, 1986).
Bulla striata View in CoL var. minor-solida Monterosato, 1884: 141.
Bulla striata var. solida Monterosato, 1884: 141 View in CoL .
Bulla striata var. stricta Monterosato, 1884: 141 View in CoL .
Bulla roperiana Pilsbry, 1895: 336 View in CoL , pl. 48, figs 19–20 (Balearic Is; types seen, 2 syntypes, ANSP 65373, H = 19.6, 20.2 mm). Clench & Turner, 1962: 130.
Bulla occidentalis View in CoL – Locard, 1897: 51 (not B. occidentalis Adams, 1850 View in CoL ). Nordsieck, 1972: 28, pl. IV, fig. 16 (in part, includes B. occidentalis View in CoL ).
Cylindrobulla occidentalis – Nordsieck & García- Talavera, 1979: 174, pl. 43, fig. 27.
Bulla media – Bergh, 1900: 217–220, pl. 17, figs 16– 25, pl. 18, figs 1–8 (figs 6–8 as ‘ B. media ( var. mediterr .)’; not Philippi, 1847).
Bulla striata var. minor Pallary, 1938: 15 View in CoL .
Retusa mariateresae Parenzan, 1970: 220 , pl. 47, fig. 905 ( Italy; types not seen).
Taxonomic history: See Discussion of Bulla striata View in CoL group above. Bruguière (1792) introduced the name B. striata View in CoL and applied it to shells from the Mediterranean, and eastern and western Atlantic. The types of Bruguière are lost (P. Bouchet, pers. comm.); however, his illustration ( Bruguière, 1792: pl. 358, figs 2a, b), unequivocally belongs to the B. striata View in CoL group. Although the name B. striata View in CoL has been applied to specimens from both sides of the Atlantic, it has been consistently and widely used in the eastern Atlantic and Mediterranean, where other names have been employed only sporadically. The situation in the western Atlantic is different and names such as B. occidentalis , B. amygdalus View in CoL and B. umbilicata View in CoL have commonly been used, besides B. striata View in CoL . To promote taxonomic stability and to maintain prevailing usage a neotype is here designated for B. striata View in CoL (ICZN, 1999: art. 75).
The name B. striata was also used by Quoy & Gaimard (1833) for specimens of B. quoyii from New Zealand and by Vayssière (1906) for unidentifiable specimens from Djibouti. The varietal names used by Monterosato (1884) and Pallary (1900) are regarded as infrasubspecific, and therefore unavailable.
The lack of understanding of this group led Parenzan (1970) to describe a new species of the genus Retusa from Italy based on juvenile shells of Bulla . Although Parenzan’s types could not be traced, his figures are unequivocal, and Thompson, Jarman & Zenetos (1985) have already pointed out this synonymy.
Diagnosis: Shell: small thick, slender, with anterior fine spiral grooves, brownish. Jaws: semicircular. Radula: with rachidian cusps decreasing in size outwards; outer lateral higher than inner; base with sharp and developed denticle. Gizzard plates: with broad dark-brown marginal band. Male genital system: vestibular area half overall length; blind caecum long, wide. Hermaphroditic gonoduct: vestibular area short; membrane gland external, laterally constricted; seminal receptacle large, globose; fertilization chamber with simple superficial structure. Range: eastern Atlantic from southern Portugal to Angola, including the Canary Islands, Cape Verde Islands, São Tomé and St Helena; entire Mediterranean Sea. COI sequences: GenBank DQ974660 View Materials (BMNH20030290); GenBank DQ986567 View Materials (BMNH20030783).
Material examined: Portugal: Ria Formosa, 12 dis, BMNH 20050338 , H = 25.0– 32.5 mm, vi.–vii.2000 / 2001, intertidal, on Zostera noltii Hornemann ; Ria Formosa, 1 sh, BMNH 20050687 , H = 28.5 mm, vi.2000, intertidal, on Zostera noltii . Spain: Barcelona, 5 sh, BMNH 20050333 , H ~ 22.1 mm; Playa de los Genoveses, Cabo de Gata , Almeria, 11 sh, MNCN 15.05 About MNCN / 41930, H ~ 24 mm; Cabo de Palos , Murcia, 21 sh, MNHC 15.05/33997, H ~ 22 mm; Cádiz, 7 sh, MNCN 15.05 About MNCN / 34006, H ~ 32 mm. Balearic Islands: 2 sh, BMNH 19970984 , H = 16.3, 20.6 mm. France: Marseille , 2 sh, BMNH 1929.10.24.141–2, H = 22.7 mm. Italy: Naples, 2 dis, BMNH 20030775 , H = 23.5, 24.6 mm, vi.2003; Palermo, 1 sh BMNH 20050326 , H = 23.3 mm. Malta: 2 sh, BMNH 20050324 , H ~ 30.2 mm, 1962; 1 sh, BMNH 20050323 , H = 34.0 mm, 1933; 3 sh, BMNH 1933611-3 , H = 29.2–32 mm; 2 sh, BMNH 20050716 , H = 32.5, 32.1 mm. Greece: Lefkas, 1 sh, BMNH 20050329 , H = 18.4 mm; Falirón, Atenas, 2 sh, MNCN 15.05 About MNCN /24719, H ~ 12.7 mm. Syria: near Tartous, 6 sh, BMNH 20050331 , H ~ 12.2 mm. Lebanon: 11 sh, BMNH 20050325 , small juveniles. Israel: Nahr Rubin, 1 sh, BMNH 192131816 , H = 10.0 mm. Egypt: Alexandria, 12 sh, BMNH 20050330 , H = 14.7– 22.8 mm. Tunisia: Djerba, 22 sh, BMNH 20050328 , H ~ 23.5 mm. Algeria: 2 sh, BMNH 1875927 , H = 19.0, 21.0 mm; Oran, 4 sh, BMNH 1911.10.26.5098–5101, H ~ 26.4 mm. Canary Islands: Barranco Hondo, Tenerife, 1 spc, BMNH 20030771 , H = 31.3 mm, ix.1994, muddy bottom, low tide 0.5 m depth; Tenerife, 1 sh, BMNH 20050327 , H = 34.5 mm. Cape Verde Islands: Sal-Rei I., Boavista I., 1 spc, BMNH 20030293 , H = 17 mm, viii.2000, live on sand, 3 m depth; 3 sh, BMNH 20030294 , H = 13.0–15.0 mm, iv.2003, dead, on sand, 1 m depth; Baía das Gatas, 2 sh, BMNH 20020733 , H ~ 9.5 mm; Tarrafal, São Tiago I., 5 sh BMNH 20050332 , H = 7.2–12.7 mm, ix.2003, dead, on sand, 3 m depth; Calhau, São Vicente I., 3 sh, BMNH 20050336 . Western Sahara: Rio do Oro , 2 sh, MNCN 15.05 About MNCN /33985, H = 20.5, 21.4. Senegal: Siné- Saloum Delta, 6 dis, BMNH 20030781 , H = 14.6– 16.4 mm, late viii.2003, low tide, on sandy-mud, 0–0.2 m deep; 3 sh, BMNH 20030781 , H = 14.5– 18.0 mm; Dakar, 1 sh, BMNH 20050334 , H = 25.4 mm. Gambia: mouth of River Gambia, 3 sh, BMNH 20050337 , H ~ 22.1 mm. Gabon: 4 sh, AMNH 20813 About AMNH /20811, H ~ 28 mm. Equatorial Guinea: Bioko , 2 sh, BMNH 1842.6.10.58-59, H = 16.8, 20.0 mm. Angola: Luanda, 3 sh, BMNH 20050335 , H = 26.2– 32.3 mm.
Shell: ( Figs 3A–L, 5A, G View Figure 5 ) Maximum H = 34.0 mm, thick, shape variable from slender- to square-ovate, wider anteriorly. North-eastern Atlantic specimens ( Portugal and Canary Islands) more quadrangular/ pyriform; Mediterranean shells usually more slender; specimens from Cape Verde Islands smaller (adult H = 10–15 mm). Left side convex; right side with posterior part slightly concave in adults; both edges convex in juveniles; 9–16 anterior spiral grooves; 2–8 posterior spiral grooves (absent in West Africa, except for Cape Verde Islands); juveniles lacking posterior spiral grooves, anterior grooves usually faint. Faint growth lines all over shell; major growth interruptions occasionally present behind lip. Parietal callus thin, occasionally covering part of spire aperture. Rear part of lip slightly folded inwards in adults. Spire aperture wide; spire 5–6 whorls, with raised striae. Protoconch diameter 100–125 Mm. Background colour varies from dark yellowish brown to whitish, mottled with dark blotches, either axially or spirally aligned, or scattered over shell; three faint dark spiral bands occasionally present anteriorly, posteriorly and behind midpoint. Aperture pale to dark with shell pattern visible; lip dark to pale brown; columella white. Periostracum brownish orange.
Animal: ( Fig. 2A View Figure 2 ) Brown with dark blotches and bright white dots scattered over cephalic shield, cephalic and parapodial lobes. Paler lozenge-shaped area between the eyes and rear part of cephalic shield, surrounded by darker pigmentation, and with an axial white line across it. Cephalic shield with anterior edge darker. Foot with abundant fine dark brown pigment spots, rear edge paler with white dots.
Jaws: ( Fig. 6A–B View Figure 6 ) Semicircular; hexagonal to quadrangular scales with smooth or slightly denticulate edges. Height 0.7–1.5 mm, width 0.5–0.8 mm (H = 23.5, 32.5 mm).
Radula: ( Figs 7 View Figure 7 , 9A–C View Figure 9 , 10A–D View Figure 10 ) In total, 23–26 rows (H = 15.6–28 mm), first three rows with rachidian teeth only. Rachidian tooth 235–425 Mm wide (H = 15.6, H = 32.5 mm); 9–18 cusps (usually 11–14); cusps triangular, sharp, usually decreasing in size outwards, with one (sometimes two) central cusp smaller. Inner lateral tooth with large central cusp twice as long an wide as others, symmetrically flanked by 3–4 cusps of decreasing size; all cusps sharp. Outer lateral tooth with 5–8 cusps; innermost largest; outermost smallest; base with sharp denticle, of similar length to outermost cusp. Outer lateral tooth higher than inner lateral.
Gizzard plates and spines: ( Fig. 11A–G View Figure 11 ) Plates 2.5– 4 mm by 0.5–1.5 mm wide (H = 32.5– 14.60 mm); light yellowish brown with broad dark brown margin; quadrangular, left edge either slightly concave or convex, right concave; prominent axial keel on left side, keel extremities occasionally grooved. Anterior gizzard spines laterally flattened or conical, slightly curved, at least twice size of posterior spines. Posterior spines needle-like, usually curved; small spines sometimes present behind plates on right side.
Male genital system: ( Fig. 13A–F View Figure 13 ) Length 8–15 mm (H = 15.6, 30.21 mm). Vestibular area about half overall length of system (excluding blind caecum), as wide as penial sheath. Prostate rounded to oval. Blind caecum long and wide.
Hermaphroditic gonoduct: ( Fig. 14A–D View Figure 14 ) Length 10– 25 mm (H = 15.6, 30.1 mm). Membrane gland external, smooth or wrinkled; seminal receptacle large, globose, wider than gametolytic duct. Gametolytic duct longer than common genital duct. Gametolytic gland of similar size to fertilization chamber. Fertilization chamber with simple network of ducts visible at surface. Vestibular area small.
Ecology: This species occurs in sheltered areas among seagrass ( Zostera spp. ) or green algae ( Ulva and Enteromorpha spp. ), on mud and sandy mud, usually on tidal flats. Templado (1982) reported occurrence in the western Mediterranean from 0 to 6 m depth,
·
among the seagrass Cymodocea nodosa and green alga Caulerpa prolifera . It is usually active at night, but can also be found crawling during daylight at low tide. In Portugal it is abundant between May and July, and was found to be common in Senegal during September.
Geographical distribution: ( Fig. 15 View Figure 15 ) Eastern Atlantic and Mediterranean, widely distributed in tropical and temperate areas. In the Atlantic it ranges from southern Portugal to Morocco, Mauritania, Bijagós Islands, southwards to Angola, including the islands of the Canaries, Cape Verde, São Tomé and Principe, and St Helena. Occurs all over the Mediterranean on both African and European coastlines from the Iberian Peninsula to Cyprus, Syria, Israel and Egypt.
Remarks: The length of adults varies among geographical localities. Specimens from Cape Verde Islands are always smaller, suggesting a case of insular dwarfism. The main shell feature that distinguishes juveniles and adults is the presence of a concave right edge in adults, due to the inward folding of the lip. This feature is absent in juveniles, in which both edges remain convex. No correlation was found between shell length and number of anterior and posterior spiral grooves.
The radula of this species is extremely variable. Specimens from Ria Formosa, Portugal, show remarkable differences in the rachidian teeth within individuals. Often the shape of cusps varies erratically along each tooth, and the number of cusps among successive teeth. Either one or two central smaller cusps can be present, and bifid cusps are common. This might be a feature of this population, because individuals from a locality in Senegal show uniform numbers and shapes of cusps. However, bifid cusps were also observed by Vayssière (1885) in specimens from Marseille, France.
The extreme variability of tooth form found in the Ria Formosa population is unusual in gastropods and suggests a rare case of intra-individual fluctuating asymmetry ( Palmer & Stobeck, 1986; Taylor & Lewis, 1995). The number of cusps was not correlated with shell size. Differences in the shape of the lateral teeth were also found within and among populations, with specimens from Senegal having narrow inner laterals than those in northern Atlantic and Mediterranean specimens. Moreover in both inner and outer laterals the number and sharpness of cusps can vary among rows, and bifid cusps can be present. The use of radular characters for systematic studies should take this variability into account.
Variation was also found in the shape of gizzard plates. In specimens from Senegal the plates are more slender and the extremities of the axial keel slightly folded to the left.
Vayssière (1885), Si (1931) and Pruvot-Fol (1954) have provided accounts of the reproductive structures of this species, based on specimens from France. However, their descriptions and illustrations are difficult to understand and not very informative. The most variable feature in the male genital system is the shape of the prostate, from oval to round, or irregular. This could be due either to the amount of sperm stored inside the prostatic duct and blind caecum, or to compression and distortion during fixation.
Regarding the female glands some specimens show a globose, smooth and inflated seminal receptaculum, whereas in others it is shrunken and constricted. This appears to be an artefact resulting from the amount of sperm inside this pouch. For example, the bifurcation of the distal connection to the gametolytic duct is particularly evident when more sperm is stored in this part of the duct, and when the amount is large it can even look as if the receptacle is made of two distinct pouches, instead of a single one. A detailed account of the morphology and function of the female glands of B. striata is given by Malaquias & Reid (2008).
The geographical distribution of this species suggests a long larval stage that enables it to establish populations not only along the entire coast of the Mediterranean Sea and West Africa to Angola, but also on offshore islands as far as St Helena. Several authors have mentioned this species from the archipelagos of the Azores ( Malaquias, 2001) and Madeira ( Malaquias et al., 2002, based on incorrect synonym), but these records should be regarded as doubtful, because no recent surveys have found this species (e.g. Mikkelsen, 1995; Wirtz, 1998; Ávila, 2000; Malaquias et al., 2002) and no specimens have been located in museums. Nobre (1932) refers to this species at Setúbal, near Lisbon. Nevertheless, this species is no longer present in this area and the northern limit is the Algarve in southern Portugal. In addition Vayssière (1906) cited its presence in Djibouti, based on resemblances with Mediterranean specimens from France, and comparing it with sympatric B. arabica (as B. ampulla ). The data provided by the author are vague and thus it is difficult to ascertain the true identity of his material, but it is unlikely to have been B. striata .
Bulla striata View in CoL can only be confused with B. occidentalis from the western Atlantic. Their morphology is so similar that Marcus (1955), comparing specimens from Brazil and the Mediterranean Sea, concluded that the differences were not enough to justify specific separation. The only difference that can be pointed out is the usual lack of posterior spiral grooves in adult shells of B. occidentalis , whereas in B. striata View in CoL they are only absent in juveniles. Nevertheless, this is a weak character because shells of B. occidentalis also sometimes show posterior spiral grooves, as also noted by Marcus (1955). The anatomy of both species is very similar and no single diagnostic character was found. DNA sequences (see Fig. 40 View Figure 40 ) were the only characters that clearly separate these two species, with minimum genetic distances between the two species (uncorrected p -distances) of 12.7% for the COI gene (see Discussion). A synopsis of the morphological variability of shell and anatomical characters, and a comparison with other species, is given in Table 1.
BULLA OCCIDENTALIS ADAMS, 1850 View in CoL
( FIGS 2B–C View Figure 2 , 3M–X, 5B, I–J View Figure 5 , 6C–D View Figure 6 , 8 View Figure 8 , 9D–H View Figure 9 , 10E–H View Figure 10 , 11H View Figure 11 , 12 View Figure 12 , 13G–L View Figure 13 , 14E–H View Figure 14 , 16 View Figure 16 )
Bulla striata Bruguière, 1792: 372 View in CoL , pl. 358, fig. 2a, b (in part, includes B. striata View in CoL ). Lamarck, 1822: 33 (in part, includes B. striata View in CoL ). d’Orbigny, 1841: 122 (in part, includes B. striata View in CoL ). Pilsbry, 1895: 332, pl. 37, figs 42–46 (in part, includes B. striata View in CoL ). Johnson, 1934: 148. Perry & Schwengel, 1955: 192, pl. 39, fig. 281. Marcus, 1957: 390. Nordsieck, 1972: 27 (in part, includes B. striata View in CoL ). Abbott, 1974: 319, pl. 10, fig. 4000. Marcus, 1976: 125 (in part, includes B. striata View in CoL ). Andrews, 1977: 162, figured. Marcus, 1977: 4 (in part, includes B. striata View in CoL ). Eisenberg, 1981: 153, pl. 135 (in part, includes B. striata View in CoL ). Abbott & Dance, 1982: 279, figured (in part, includes B. striata View in CoL ). García-Talavera, 1983: 34 (in part, includes B. striata View in CoL ). Vokes & Vokes, 1983: 33, pl. 22, fig. 5. Sterrer, 1992: 142. Andrews, 1994: 75, figured. Diaz & Puyana, 1994: 244, fig. 983 (in part, includes B. striata View in CoL ). Rios, 1994: 197, pl. 65, fig. 929 (in part, includes B. striata View in CoL ). Mikkelsen, 1996: 436.
Bulla umbilicata Röding, 1798: 15 View in CoL (invalid name, ICZN, 1959: opinion 549; types untraceable). Redfern, 2001, pl. 70, fig. 659.
Bulla amygdalus Solander View in CoL , in Dillwyn, 1817: 480 (in part, includes B. striata View in CoL ; possible types seen, 2 sh, BMNH Banks coll., H = 21.6, 27.6 mm; nomen dubium). Adams, 1850: 575, pl. 122, fig. 72. Sowerby, 1868: sp. 7, pl. 3, figs a, b.
Bulla amygdala View in CoL – Pilsbry, 1895: 329, pl. 38, figs 49– 50, 62–65, pl. 39, fig. 79. Nordsieck, 1972: 28, pl. 4, fig. 14 (in part, includes B. striata View in CoL ). Dall & Simpson, 1901: 363. Perry & Schwengel, 1955: 192, pl. 39, fig. 282.
Bulla media Philippi, 1847: 121 (Mare Antillarum; types untraceable; nomen dubium). Adams, 1850: 576, pl. 123, fig. 70. Sowerby, 1868: sp. 11, pl. 4, figs a, b. Mabille, 1896: 118. Bergh, 1900: 217, pl. 17, figs 16–25, pl. 18, figs 1–5 (in part, includes B. striata View in CoL ; see pl. 28, figs 6–8; in part, includes Bulla spp. from Pacific Ocean).
Bulla occidentalis Adams, 1850: 577 View in CoL , pl. 123, figs 72, 73 (St. Vincent’s, West Indies [Lesser Antilles]; lectotype here designated, BMNH 197649, H = 14.2 mm, Fig. 3S herein). Menke, 1854b: 45. Sowerby, 1868: sp. 14, pl. 5, figs a–d. Dall, 1889: 55. Pilsbry, 1895: 331, pl. 38, figs 51–53, 55–60, pl. 39, figs 77, 78. Mabille, 1896: 114. Peile, 1926: 85. Johnson, 1934: 148. Perry & Schwengel, 1955: 192, pl. 39, fig. 280. Rios, 1970: 131. Vilas & Vilas, 1970: 91. Nordsieck, 1972: 28, pl. IV, fig. 16 (in part, includes B. striata View in CoL ). Marcus, 1976: 4. Eisenberg, 1981: 152, pl. 134. Damerval & Damerval, 1990: 190, pl. 35, fig. 1. Abbott & Dance, 1991: 279, figured: 170. Redfern, 2001: 158, pls 70, 115, fig. 658A, B.
Bullaria occidentalis – Peile, 1926: 85.
Bulla rubiginosa Gould, 1852: 221 View in CoL , pl. 15, fig. 226 (not introduced in Gould, 1849 as cited by Gould, 1852; Rio de Janeiro, Brazil; types not seen, probably lost; see Johnson, 1964). Pilsbry, 1895: 330, pl. 39, fig. 76. Mabille, 1896: 119.
Bulla (Bullea) nux Menke, 1853: 140 View in CoL (Atlantic, Cuba; types probably lost, see Dance, 1986). Pilsbry, 1895: 350.
Bulla (Bullea) sulcata Menke, 1853: 138 View in CoL (São Pedro dos Indios, Brazil; types probably lost, see Dance, 1986).
Taxonomic history: See Discussion of Bulla striata View in CoL group above. The present species has a more variable shell than its eastern Atlantic sister species B. striata View in CoL , and this variability has resulted in a profusion of available names based on vague descriptions. The confusion surrounding B. amygdala Solander View in CoL , in Dillwyn, 1817 has been discussed earlier; this name is regarded as a nomen dubium because it cannot be applied with certainty to specimens from either the eastern or the western Atlantic. Philippi (1847) gave a detailed diagnosis of B. media , a type locality of Mare Antillarum and, although types cannot be traced, he quoted recognizable figures by Gualtieri (1742) and Lister (1685 –92). This is potentially the earliest available name, although it has not been used since 1900. The next available name is B. occidentalis , introduced by Adams (1850) for shells from St Vincent, West Indies, and represented by a type specimen (lectotype here designated). Adams (1850) described the shells of B. occidentalis as ‘... variegated with red-brown spots and waved lines... ’. In fact only one among the six beach worn syntypes bears this peculiar pattern, whereas the others show a more typical B. striata View in CoL -group pattern. This name has been in continuous usage for western Atlantic specimens since its first description. The few references to the eastern Atlantic derive from Locard (1897) who used this name for specimens from the Canary Islands, and was subsequently quoted in two other works ( Nordsieck, 1972; Nordsieck & García- Talavera, 1979). We recommend reversal of precedence in this case, in order to maintain prevailing usage of the name B. occidentalis , but this will require a ruling from the ICZN. Pending an application to the Commission, we maintain usage of the junior synonym (ICZN, 1999: art. 23.9.3).
Diagnosis: Shell: small, thick, slender, anterior fine spiral grooves present, brownish. Jaws: semicircular. Radula: rachidian cusps decreasing in size outwards; outer lateral higher than inner; base with sharp and developed denticle. Gizzard plates: left edge straight, conspicuous marginal dark brown band. Male genital system: vestibular area half overall length (excluding blind caecum); blind caecum long and wide. Hermaphroditic gonoduct: short vestibular area, membrane gland external, smooth; seminal receptacle large, globose; fertilization chamber with simple superficial structure. Range: Bahamas, Florida to southern Brazil. COI sequences: GenBank DQ986542 View Materials – 986543 (BMNH 20050351/4; BMNH20030779/1); GenBank DQ986547 View Materials –986548 (BMNH20030340; BMNH20030349).
Material examined: Bermuda: 4 sh, BMNH 1911.12.21.431-4, H ~ 18.2 mm. Bahama Islands: Eleuthera I., 14 sh, BMNH 20050342 , juveniles. Florida: Miami , 1 dis, BMNH 20030045 , H = 28.8 mm, i.1988, shallow seagrass beds; NW Fort Pierce Inlet , 3 dis, FLMNH 303042 View Materials , H = 15.1– 16.2 mm, iv.2003, 0.1 m deep, silt with mangroves; Florida Keys, 2 dis, BMNH 20030777 , H = 15.0, 19.28 mm; Sebastian Inlet, 9 sh, BMNH 20050341 , H ~ 28.2 mm. Texas: 1 sh, BMNH 19970937 , H = 24.1 mm; Aransas Bay , 4 sh, MNCN 15.05 About MNCN /33993, H ~ 22.2 mm . Mexico: Bahia de Punta Soliman , Quintana Roo, 4 sh, BMNH 20050646 , H = 0.6–11 mm, v.2005, washed ashore . Cuba: 5 sh, BMNH 1854.10 .4.6 (d’Orbigny collection), H = 10–37.7 mm; 5 sh, BMNH 1854.10 .4.7 (d’Orbigny collection), H = 13.4–28.4 mm; Ensenada de la Broa , 2 dis, DZUO 039802 , H = 24.6, 29.40 mm, iii.1998; Cape of Santo António , 1 dis, BMNH 20050351 , H = 10.2 mm, xii.2004, 2– 3 m deep on Thalassia sp. ; Cienfuegos, 1 dis, BMNH 20030848 , H = 25.2 mm, viii.2003; São Tiago, 4 sh, BMNH 1950817173–76 , H ~ 19.96 mm; La Habana, 18 sh, BMNH 20050649 , H = 16.6– 26.5 mm, ix.1999, washed ashore . Jamaica: 3 sh, BMNH 20060655 , H = 26.4–28.7 mm; 5 sh, BMNH 1904.10.25.72–76, H = 20.0– 30.6 mm; 5 sh, BMNH 20050343 , H = 24.3–25.0 mm; 3 sh, BMNH 20050719 , H = 26.6–28.0 mm . Virgin Islands: Bessup Bay , St Thomas, 2 sh, AMNH 127163 About AMNH , H = 38.0 mm; Christiensted Harbor, St Croix , 2 sh, AMNH 190684 About AMNH , H = 40.0, 45.0 mm . Barbados: 1 sh, BMNH 19723 , H = 30.0 mm. St Vincent : 4 sh, BMNH 196968 , 3 sh, BMNH 197649 . Martinique: 4 sh, BMNH 1854.10 .4.5 (d’Orbigny collection), H = 9–24.2 mm; St Martin : 3 sh, BMNH 20050344 , H ~ 25.8 mm . Antigua: English Harbour , 1 spc, CAS 067267 About CAS , H = 24.0 mm, vii.1918, 2 m depth ; Guiana I., 3 sh, BMNH 1934.2.13.303–5, H ~ 24.6 mm . Guadeloupe: 7 sh, BMNH 20050717 , H = 11.2–15.8 mm. St Lucia: 6 sh, BMNH 20050347 , H = 17.9–24.1 mm; 1 sh, BMNH 19970934 , H = 21.5, 11 sh, BMNH 20050340 , H = 18.6–25.40 mm . Trinidad: Las Cuevas Bay , 10 sh, BMNH 20050346 , H = 13.6–19.3 mm . Tobago: Pigeon Point beach, 8 sh, BMNH 20050348 , H = 19.1–24.9 mm; Nylon Pool, Buccoo Bay , 8 sh, BMNH 20050345 , H = 17.0– 20.2 mm, 8.viii.1973, on shallow sandy bottom . Belize: Lighthouse Reef, 1 sh, BMNH 20050350 , H = 22.2 mm . Panama: Bocas del Toro, 2 sh, BMNH 20050647 , H = 15.0, 20.0 mm, iv.2005, washed ashore; Buena Ventura I., 2 sh, BMNH 20050648 , H = 19.4, 31.4 mm, iv.2005, washed ashore . Colombia: El Rodadero, Santa Marta , Magdalena Province, several shells, RMNH. MOL.CR.1540, 1– 2 m deep, viii.1975 . Venezuela: 2 sh, BMNH 20050349 , H = 25.3, 30.3 mm; Amuay, Paraguana , 1 sh, AMNH 190706 About AMNH , H = 35.0 mm . Surinam: Paramaribo, several shells, RMNH 1911 About RMNH . Brazil: Pernambuco, 1 sh, BMNH 19567126 , H = 26.5 mm; Pará, 1 sh, BMNH 18456646 , H = 31.5 mm; Ilha de Frades, Bahia , 17 sh, BMNH 20050339 , H = 15.6–24.6 mm; Alagoas , Recife , 3 dis, BMNH 20030340 , H = 19.4–20.4 mm, iv.2003, collected at night, 0.5 m, on mud; Pedra da Andorinha , São Paulo, 2 dis, MZUSP No 94, H = 26.1, 27.2 mm, 1962, 1 m deep; Ubatuba, São Paulo, 3 dis, MZUSP 30009 View Materials , H = 33.3–35.1 mm; São Sebastião I., São Paulo, 3 dis, MZUSP 24889 View Materials , H = 27.1–30.9 mm, vii.1984, sheltered lagoon with seagrass .
Shell: ( Figs 3M–X, 5B, I–J View Figure 5 ) Maximum H = 45.0 mm. Thick, shape variable within and among geographical regions, from slender to pyriform, globular or squareovate. Very thick shells found in Jamaica. Left side convex, right side with posterior part slightly concave in adults. Rear part of lip slightly folded inwards in adults. In total, 4–11 conspicuous fine anterior spiral grooves present. Posterior spiral grooves generally absent, usually faint when present (maximum five). Growth lines faint, present all over shell. Columella white, extremely thick in some specimens. Parietal callus thin. Spire aperture wide. Spire with 5–6 whorls (usually six) ornamented with conspicuous laminated striae; sutures distinct or indistinct. Protoconch diameter 100–150 Mm. Background colour variable from whitish, light or dark brown, to yellow brown mottled with brown dots, sometimes merging together creating either large blotches or bands axially or spirally orientated. Three dark spiral bands sometimes present, two at extremities and one between middle and posterior part of shell. Occasionally with a fourth faint band between anterior and middle ones. Shell pattern visible through aperture. Lip whitish to pale brown. Periostracum brownishorange.
Animal: ( Fig. 2B–C View Figure 2 ) Brown with bright white dots scattered over cephalic shield, cephalic and parapodial lobes.
Jaws: ( Fig. 6C–D View Figure 6 ) Semicircular; scales hexagonal to quadrangular, with either smooth or denticulate edges. Conspicuous denticulation was found in specimens from Cuba and Brazil. Height 0.7–1.1 mm, width 0.4–0.6 mm (H = 20.3, 22.7 mm).
Radula: ( Figs 8 View Figure 8 , 9D–H View Figure 9 , 10E–H View Figure 10 ) In total, 18–28 rows (H = 10.2–33.6 mm), first three rows with rachidian teeth only. Rachidian teeth 200–400 Mm wide (H = 16.2, 30.9 mm); 9–15 cusps both in smaller (H ~ 10–20 mm) and larger specimens (H ~ 25– 30 mm); cusps triangular, occasionally slender, all sharp, decreasing in size outwards (few exceptions observed); central of smaller size, occasionally absent. Inner lateral teeth with major central cusp, symmetrically flanked by 3–4 (sometimes five) cusps on either side of decreasing size; central cusp two to three times longer and wider, occasionally curved inwards; cusps usually sharp, sometimes blunt. Outer lateral with 5–8 cusps, innermost cusp longer and wider, usually twice the size of second cusp, either sharp or blunt; cusps decreasing in size downwards, all sharp; base with sharp denticle of similar length to outermost cusp. Outer lateral higher than inner lateral.
Gizzard plates and spines: ( Figs 11H View Figure 11 , 12 View Figure 12 ) Plates 1.3– 4 mm by 0.6–1.8 mm wide (H = 19.20, 35.10 mm); plates light yellowish brown with broad marginal dark brown band; quadrangular, left edge straight or slightly convex, right concave; axial keel on left side, prominent, groove on extremities either present or absent, surface smooth or formed by layers partly overlaping each other. Anterior gizzard spines laterally flattened, slightly curved, at least twice the size of posterior spines. Posterior spines needle-like, usually curved; small spines occasionally present behind plates on right side.
Male genital system: ( Fig. 13G–L View Figure 13 ) Total length 7–20 mm (H = 15.0, 29.4 mm). Vestibular area about half overall length of system (excluding blind caecum), wider than penial sheath. Prostate short, rounded to oval. Blind caecum long and wide.
Hermaphroditic gonoduct: ( Fig. 14E–H View Figure 14 ) Total length 11–18 mm (H = 16.2, 26.9 mm). Membrane gland external and smooth. Seminal receptacle large and globose, wider than gametolytic duct. Gametolytic duct either longer or shorter than common genital duct. Gametolytic gland of similar size or slightly larger than fertilization chamber. Fertilization chamber with simple network of ducts visible at surface. Vestibular area short.
Ecology: This species occurs in shallow areas with muddy bottoms and seagrass down to 3 m deep. Specimens from Florida (Fort Pierce inlet) were found crawling during the day (P. Mikkelsen, pers. comm.), whereas Humfrey (1975) noted that in Jamaica this species was buried a few centimetres within the sediment during the day and was active at night. In Florida this species was found to spawn during March and eggs were found attached to the seagrass Halodule wrightii Aschers (P. Mikkelsen, pers. comm.). Marcus (1957) mentioned that this species was common around São Paulo, Brazil, in November.
Geographical distribution: ( Fig. 16 View Figure 16 ) This is a western Atlantic species occurring in temperate and tropical areas from Bermuda to southern Uruguay. It occurs in both coasts of Florida, Texas, and Mexico, and there are scattered records on the mainland coast of Central America, in Belize, Costa Rica and Panama; in the Caribbean it is present in the Bahamas, Greater and Lesser Antilles from Cuba, Jamaica and Puerto Rico to Trinidad. It is found on the coast of South America in Colombia, Venezuela and Surinam, and along the coast of Brazil to Uruguay.
Remarks: The shell and anatomical data provided by Bergh (1900; as B. media ) and Marcus (1955, 1957; both as B. striata ) agree with the variability found in the studied material.
This species has a more variable shell than its eastern Atlantic sister species B. striata . It usually lacks posterior spiral striae, and can reach larger dimensions.
The width of the rachidian tooth is correlated with shell size but, as in B. striata , the number of rachidian cusps is not. Variation was found in the rachidian teeth of single individuals within and among rows, as described in B. striata , and in some cases bifid and trifid cusps were present. Inner lateral teeth can show great variation from narrow with minute lateral cusps and a large central cusp, to broad with either the same or different numbers of cusps on either side; tooth shape can change along the radula, and teratologies can be found with entire longitudinal columns of inner laterals of aberrant form. Cases of bifid and trifid cusps are rare in lateral teeth.
Variation in gizzard plates is mostly ontogenetic, with smaller specimens (H <15 mm) often having concentric rings on the surface of the plates, whereas in larger specimens the surface is worn smooth due to use.
A high degree of variation was found in the shape of the male genital system with the prostate ranging from rounded to oval, or even undifferentiated and of similar size to the penial sheath. The blind caecum varies from half to twice the length of the prostate. As pointed out for B. striata , this is likely to result from differences in the amount of stored sperm or from preservation artefacts; the differences are not correlated with ontogenetic stages or geographical origin. In some cases the disparity among specimens collected at the same site exceeds the differences found between the extremes of the geographical range.
In the female glands variation was found mostly in the length of the gametolytic duct. Particularly in specimens from Brazil this duct is extremely variable; some individuals show a duct as short as the common genital duct, whereas in others the duct reaches beyond the fertilization chamber. Variation in shape of the seminal receptacle was also observed as described in B. striata .
This species is similar to B. striata and the main differences are described above (see Remarks on B. striata ). A synopsis of the morphological variability of shell and anatomical characters, and a comparison with other species, is given in Table 1.
BULLA MABILLEI LOCARD, 1897 View in CoL
( FIGS 2D View Figure 2 , 4A–D, 5C View Figure 5 , 6E View Figure 6 , 15 View Figure 15 , 18A–B View Figure 18 , 19A–B View Figure 19 , 20A–B View Figure 20 , 21A–B View Figure 21 , 22A–B View Figure 22 , 23A–B View Figure 23 , 24A–B View Figure 24 )
Bulla ampulla View in CoL – d’Orbigny, 1839: 45 (not Linnaeus, 1758). Guerreiro & Reiner, 2000: 209, figured (not Linnaeus, 1758).
Bulla cruentata Adams, 1850: 577 View in CoL , pl. 126, fig. 75 ( Moluccas [in error]; types seen, 3 syntypes BMNH 197652, H = 45.5, 45.6, 53.1 mm). Mabille, 1896: 116.
Bulla solida A. Adams View in CoL in Sowerby, 1868: sp. 10, pl. 4, figs a, b (no locality; type seen, holotype BMNH 1976046, H = 34.5 mm, figured in Willan, 1978: 60, figs 1, 2; not Gmelin, 1791). Mabille, 1896: 116 (not Gmelin, 1791).
Bulla punctata A. Adams View in CoL in Sowerby, 1868: sp. 15, pl. 5, figs a, b (no locality; types seen, 2 syntypes BMNH 197647, H = 38.2, 39.2 mm; not Schröter, 1804). Watson, 1897: 276. Nobre, 1937: 15.
Bulla angasi Pilsbry, 1895: 347 View in CoL , pl. 36, figs 32, 33 (replacement name for B. solida A. Adams View in CoL in Sowerby, 1868).
Bulla mabillei Locard, 1897: 50 View in CoL , pl. 2, figs 1, 2 ( São Vicente I., Cape Verde, 16°96′N 20°23′W; types seen, 2 syntypes MNHN; lectotype here designated H = 50.9 mm, figured in Valdés & Héros, 1998: 709, pl. 7, fig. B). Odhner, 1932: 25. Marche-Marchad, 1956: 59. Marche-Marchad, 1958: 39. Marcus & Marcus, 1966: 155. Nordsieck, 1972: 28, pl. 4, fig. 17. Talavera, 1978: 126. Cosel, 1982a: 31. Cosel, 1982b: 21. Cosel, 1982c: 61. Bernard, 1984: 108, pl. 54, fig. 216. Sanchez & Batet, 1991: 218. Wirtz, 1995: 184, figured. Malaquias & Calado, 1997: 154. Malaquias et al., 2002: 73, figs 2–4. Ardovini & Cossignani, 2004: 242. Cervera et al., 2004. Rolán, 2005: 389, figs 936, 937.
Cylindrobulla mabillei – Nordsieck & García- Talavera, 1979: 174, pl. 43, fig. 29.
Bulla amygdala – Malaquias, 2000: 92 (not Dillwyn, 1817 = B. striata ).
Taxonomic history: This species has a particularly complex nomenclature because names have been proposed based on specimens with incorrect or unknown localities. Nevertheless, the large bright reddish shell of this eastern Atlantic species is distinctive among worldwide species of Bulla , so that its recognition is straightforward.
Adams (1850) introduced the name B. cruentata for large reddish shells said to originate from the Moluccas, using material made available by H. Cuming. Very few shells bearing this name are housed in museums worldwide (specimens were traced in four museums: AMNH, MNCN, NHM and ZMB); all lots are approximately contemporaneous and labelled simply ‘Moluccas’, suggesting that all were split from a single original lot or that the locality was added to unlocalized shells. Apart from the original description and a few subsequent works based on the same material (e.g. Sowerby, 1868; Pilsbry, 1895), there are no further references to B. cruentata and it has not been collected during field work recently undertaken in the area. It is therefore likely that this species was wrongly localized and was based on specimens from the eastern Atlantic. No author to our knowledge has used B. cruentata as a valid name since Mabille (1896).
Adams (1850) described and illustrated B. punctata from Panama. However, in the index to the work (p. 607) he added a footnote that the name B. punctata was a misspelling of B. punctulata , so his name B. punctata is not available (it is in any case a junior primary homonym). Nevertheless A. Adams did in fact label specimens presented to him by H. Cuming as B. punctata , and this name was later validly introduced by Sowerby (1868) based on those specimens bearing Adams’ manuscript name. These specimens have no locality, but are large and reddish, and this was enough to lead two authors to use the name for this eastern Atlantic Bulla species ( Watson, 1897; Nobre, 1937).
Bulla solida View in CoL was another manuscript name of A. Adams that was validated by Sowerby (1868) and applied to another reddish shell from unknown locality in the H. Cuming collection. Pilsbry (1895) noted that this name was preoccupied by B. solida Gmelin, 1791 View in CoL , and replaced it with B. angasi View in CoL . Based on a quote by Angas (1867), Pilsbry attributed the origin of this species to eastern Australia. However Angas’ (1867) quotation on B. solida View in CoL is very unclear, referring to a manuscript name by Gmelin (which cannot be traced and of which no specimen is available, as also noted by Willan, 1978), while at the same time apparently giving a description of Sowerby’s B. solida View in CoL , which he was likely to have examined during his collaboration with A. Adams at the British Museum ( Hedley, 1913; Iredale, 1959). Despite these contradictions, Willan (1978) considered B. angasi View in CoL the valid name for an Australian species and designated the shell of B. solida A. Adams View in CoL in Sowerby, 1868, as the lectotype of B. angasi Pilsbry, 1895 View in CoL . In fact a replacement name applies to the same type as the name it replaces, so this designation is unnecessary. This particular shell has a peculiar coloration of large red blotches resulting from the merging of several dots, but apart from this bears all the features of the reddish eastern Atlantic Bulla View in CoL . The name B. solida A. Adams View in CoL in Sowerby, 1868 is a junior homonym, and is therefore invalid. The replacement name B. angasi View in CoL is potentially available, though lacking a type locality. However, since its introduction by Pilsbry (1895) it has never been used for the present species (see Synonymy of B. vernicosa View in CoL ).
The first author explicitly to name this species from the eastern Atlantic was Locard (1897), based on specimens collected in the Cape Verde archipelago. Interestingly, he acknowledged the similarity between his B. mabillei and B. cruentata View in CoL , refering to the former as an intermediate between B. ampulla View in CoL and B. cruentata View in CoL . The name B. mabillei has since become familiar for this Atlantic species and has been widely used by numerous authors (see Synonymy above). Pending formal application to ICZN (1999: art. 23.9.3) for a ruling on protection of this name (in relation to its available senior synonyms B. cruentata View in CoL and B. angasi View in CoL ), stability is best served by maintaining prevailing usage.
Without explanation Nordsieck & García-Talavera (1979) transferred B. mabillei to Cylindrobulla (a genus of sacoglossan gastropods). Malaquias (2000) considered B. mabillei a synonym of B. amygdala , which is clearly a name in the B. striata group. d’Orbigny (1839) had earlier misidentified specimens from the Canary Islands as B. ampulla .
Diagnosis: Shell: large, thick, globose, reddish. Jaws: elongated. Rachidian cusps of similar size (except smaller marginal and central); inner and outer laterals of similar height; outer lateral cusps of same size, base with short denticle. Gizzard plates: light brown with fine darker rings. Male genital system: vestibular area less than half overall length (excluding blind caecum); blind caecum long and thin; prostate and penial duct coiled. Hermaphroditic gonoduct: vestibular area long; first part of membrane gland internal; seminal receptacle small; fertilization chamber with complex superficial structure. Range: insular species from Madeira to São Tomé and Princípe. COI sequences: GenBank DQ986533 View Materials –986534 (BMNH 20060099, BMNH20060097).
Material examined: Madeira Archipelago: 2 sh, BMNH 1911.10.26.5113-5114, H = 21.1, 40.2 mm; 2 sh, BMNH 20050355 , H = 35.1–49.2 mm; southern Madeira , 2 sh, MMF( HN) 30485, H = 19.0, 51.2 mm; Porto Santo I., 3 sh, BMNH 20050352 , H = 25.0– 37.4 mm. Canary Islands: Gran Canaria, 1 sh, BMNH 1996359 , H = 36.5 mm; Lanzarote, 1 sh, BMNH 1993889 , H = 44.9 mm; Las Teresitas, Tenerife, 1 dis, BMNH 20050711 , H = 29.7 mm, xi.1995, 8 m sandymud with gravel; Garachico, Tenerife, 3 dis, BMNH 20020457 , H = 20.4–26.9 mm, ix.2002, 5– 10 m, sandymud with rocks and algae; Barranco Hondo, Tenerife, 1 dis, BMNH 20030774 , H = 48.4 mm, ix.2003, 0.5 m in mud; Tenerife (north coast), 10 sh, BMNH 20050354 , H = 13.2–25.4 mm. Cape Verde Islands: 1 sh, BMNH 1996358 , H = 58.9 mm; 2 sh, BMNH 1905.7.13.10-11, H = 57.8, 61.4 mm; Calhau, São Vicente I., 6 sh, BMNH 20060525 , H = 26.7–40.0 mm. São Tomé and Princípe: 4 sh, BMNH 20050353 , H = 31.3–34.2 mm; 2 sh, BMNH 18981223-4 , H = 34.1, 43.50 mm .
Shell: ( Figs 4A–D, 5C View Figure 5 ) Maximum H = 61.4 mm. Large, thick, oval to ovate-quadrangular. Faint regular growth lines all over shell; major growth interruptions occasionally present behind lip. Spire aperture wide; spire of 6–7 whorls, with conspicuous raised striae; sutures indistinct. Protoconch diameter 140–160 Mm. Parietal callus usually thin and whitish. Background colour pale to bright red, with dark red blotches shadowed with white, merging to form zigzag axial bands or even large blotches. Aperture white, occasionally shell pattern visible through the aperture. Columella reddish white. Lip dark red. Periostracum not distinguishable.
Animal: ( Fig. 2D View Figure 2 ) Uniform reddish brown with white blotches scattered all over body. Periocular area unpigmented. Colour photographs of the animal can be found in Sánchez & Batet (1991) and Wirtz (1995).
Jaws: ( Fig. 6E View Figure 6 ) Elongate; hexagonal scales with smooth edges. Height = 1.8 mm, width = 0.6 mm (H = 29.7 mm).
Radula: ( Figs 18A–B View Figure 18 , 19A–B View Figure 19 , 20A–B View Figure 20 ) In total, 24–36 rows (H = 20.4–28.4 mm), first four rows with rachidian teeth only. Rachidian teeth 582 Mm (H = 20.4 mm) to 700–1300 Mm wide (H = 26.9, 48.4 mm); 11 cusps; cusps sharp, triangular with quadrangular base; central and marginal cusps smaller, other cusps of similar size. One large specimen (H = 48.4 mm) with 12–14 cusps, of irregular shapes, all same size, only occasionally with smaller central and marginal cusp. Inner lateral teeth wide, with major central cusp symmetrically flanked on either side by three (seldom four) smaller cusps of decreasing size; central cusp twice as wide; all cusps sharp. Outer lateral with 4–6 sharp cusps, outermost smaller; base with short denticle. Inner and outer laterals of same height.
Gizzard plates and spines: ( Figs 21A–B View Figure 21 , 22A–B View Figure 22 ) Plates 3.2–4 mm by 1–1.5 mm wide (H = 20.4, 29.7 mm); light brown, with few dark brown rings, central area yellowish; quadrangular, left and right edges concave; prominent axial keel on left side, slightly grooved at extremities; surface smooth. Anterior gizzard spines triangular, laterally flattened, at least twice the size of posterior spines. Posterior spines needle-like, usually curved; small spines sometimes present behind plates on right side.
Male genital system: ( Fig. 23A–B View Figure 23 ) Total length 13–28 mm (H = 26.9, 48.4 mm). Vestibular area about one-third overall length (excluding blind caecum); narrower than penial sheath. Prostate oval, only slightly wider than penial sheath. Blind caecum long and thin. Prostate and penial duct coiled.
Hermaphroditic gonoduct: ( Fig. 24A–B View Figure 24 ) Total length 28–43 mm (H = 29.7, 48.4 mm). First part of membrane gland internal; second part external and smooth. Seminal receptacle small, usually globose, of similar width to gametolytic duct. Gametolytic duct longer than common genital duct. Gametolytic gland larger than fertilization chamber. Fertilization chamber with complex network of ducts visible at surface. Vestibular area long.
Ecology: This is a subtidal species that lives from shallow waters down to 70 m depth, on sandy mud with rocks and pebbles covered with algae.
Geographical distribution: ( Fig. 15 View Figure 15 ) This is an eastern Atlantic species restricted to the oceanic islands of the archipelagos of Madeira, Selvagens, Canaries, Cape Verde, and São Tomé and Princípe.
Remarks: Anatomical data on this species was briefly given by Malaquias et al. (2002), based on a single specimen from the Canary Islands and therefore intraspecific variability was not evaluated. This is a uniform species in shell shape, colour and anatomy. Nevertheless small shells (H = 30–34 mm) occasionally can be thick and solid (BMNH 20050353; from São Tomé). Variation of the radula was found in the rachidian teeth of a large specimen (H = 48.4 mm) as described above.
This species can only be confused with B. solida , with which it shares a very similar shell shape and anatomy. However, the reddish coloration of the shell of B. mabillei is unique among the Atlantic Bulla . Also the penial duct of B. mabillei is coiled throughout its length, whereas that of B. solida is uncoiled. Nevertheless this character is weak, because intermediates were found. A molecular comparison between these two species was not possible as we were unable to obtain specimens of B. solida fixed for DNA extraction. Because of their close similarity, the two are likely to be sister taxa. A synopsis of the morphological variability of shell and anatomical characters, and a comparison with other species, is given in Table 1.
The few references to the West African mainland coast must be regarded as doubtful (e.g. Gabon, Bernard, 1984; Angola, Rolán & Ryall, 1999); these specimens were probably collected offshore by trawlers.
BULLA SOLIDA GMELIN, 1791 View in CoL
( FIGS 4E–H, 5D View Figure 5 , 6F–G View Figure 6 , 16 View Figure 16 , 18C–D View Figure 18 , 19C–D View Figure 19 , 20C–D View Figure 20 , 21C–D View Figure 21 , 22C–D View Figure 22 , 23C–D View Figure 23 , 24C–D View Figure 24 )
Bulla solida Gmelin, 1791: 3434 View in CoL (no locality; neotype here selected, H = 40.4 mm BMNH 20050356, Los Tanques, Paraguana, Venezuela, Fig. 4E herein). Pilsbry, 1895: 335, pl. 37, figs 36–38, pl. 38, fig. 54, pl. 43, figs 1–2. Johnson, 1934: 148. Krebs, 1864: 93. Morris, 1973: 264, pl. 72. Bandel, 1976: 89. Marcus, 1976: 121, figs 1–7. Marcus, 1977: 4. Diaz & Puyana, 1994: 244, fig. 984. Mikkelsen, 1996: 436. (Not B. solida A. Adams View in CoL in Sowerby, 1868.)
Bullea solida – Mörch, 1875: 173.
Bulla ampulla View in CoL – d’Orbigny, 1841: 121 (not Linnaeus, 1758).
Taxonomic history: Gmelin (1791) described this species from an illustration in Knorr (1772: pl. 21, fig. 2), based on a specimen from Martinus Houttuyn’s collection (see Boeseman & de Ligny, 2004: 70). Both Knorr’s illustration and Gmelin’s description are poor, but they probably refer to this western Atlantic species. The similarity of shape to B. ampulla and the coloration of the shell and lip mentioned by Gmelin (1791) agree with this species. Unfortunately neither Knorr nor Gmelin provided data on geographical origin. The Houttuyn collection was sold and dispersed ( Boeseman & de Ligny, 2004), and type material could not be traced. To remove possible ambiguity, a neotype is designated.
This uncommon, but distinctive, species has seldom been referred to in the literature, and apart from the misidentification by d’Orbigny (1841) has always been named B. solida .
Diagnosis: Shell: large, thick, globose, pinkish brown. Jaws: elongated. Radula: rachidian cusps of similar size (except marginal and central); inner and outer laterals of similar height; outer lateral cusps of same size; base with short denticle. Gizzard plates: reddish brown with narrow submarginal dark brown band. Male genital system: vestibular area less than half overall length (excluding blind caecum); blind caecum long, thin; prostate coiled. Hermaphroditic gonoduct: vestibular area long; first part of membrane gland internal; seminal receptacle small; fertilization chamber with complex superficial structure. Range: western Atlantic from Florida to Venezuela.
Material examined: Florida: Indian River , 3 dis, HBOM 62 View Materials : 281, H = 34.9–40.0 mm, vii.1974, 25 m; 1 dis, HBOM 65 View Materials : 282, H = 35.1 mm, February 1974, 14 m; Fort Lauderdale , 1 sh, AMNH 3093 About AMNH , H = 42.5 mm, 15–20 m. Honduras: Punta Patuca / Castilla , 5 sh, AMNH 230317 About AMNH , H = 44.5–50.8 mm, 27–37 m. Panama: 1 sh, BMNH 20050358 , H = 45.4 mm. Venezuela: Los Tanques, Paraguana, 1 sh, BMNH 20050356 , H = 40.4 mm; 2 sh, BMNH 20050357 , H = 26.9, 41.2 mm; 3 sh, BMNH 20050359 , H = 29.2–43.8 mm. Guadeloupe: 1 sh, BMNH 1854.10 .4.3 (d’Orbigny collection), H = 44.4 mm. Martinique: 1 sh, BMNH 1854.10 .4.4 (d’Orbigny collection), H = 29.6 mm. Cuba: 2 sh, BMNH 1854.10 .4.2 (d’Orbigny collection), H = 35.2, 37.7 .
Shell: ( Figs 4E–H, 5D View Figure 5 ) Maximum H = 50.8 mm. Large, thick, oval to ovate-quadrangular. Faint regular growth lines present; major growth interruptions occasionally present behind lip. Spire aperture wide; spire 6–7 whorls, with conspicuous raised striae; sutures indistinct. Protoconch diameter about 100 Mm. Parietal callus thin. Background colour pinkish brown, with dark brown blotches shadowed with white, sometimes merging to form axial or spiral bands. Aperture white, shell pattern often visible through aperture. Columella white. Lip pinkish. Periostracum not distinguishable.
Animal: No data available.
Jaws: ( Figure 6F–G View Figure 6 ) Elongate; triangular scales with smooth edges. Height 2.0– 2.4 mm, width 0.4–0.5 mm (H = 35.1, 40.0 mm).
Radula: ( Figs 18C–D View Figure 18 , 19C–D View Figure 19 , 20C–D View Figure 20 ) In total, 28–31 rows (H = 40.0– 35.1 mm), first 4–6 rows with rachidian teeth only. Rachidian teeth 582 Mm (H = 20.4 mm) to 618–667 Mm wide (H = 35.1, 40.0 mm); 11–13 cusps; cusps sharp, triangular with quadrangular base; central and outwards 1–2 marginal cusps smaller, other cusps of similar size. Inner lateral teeth wide, with major central cusp symmetrically flanked on either side by three (seldom four) smaller cusps of decreasing size; central cusp twice as wide; all cusps sharp, occasionally blunt. Outer lateral with five sharp cusps, outermost smaller; base with short denticle. Inner and outer laterals of same height.
Gizzard plates and spines: ( Figs 21C–D View Figure 21 , 22C–D View Figure 22 ) Plates 6 mm by 2–2.5 mm wide (H = 34.9, 38.4 mm); reddish brown with a narrow submarginal dark brown band, central area yellowish, fine regular concentric dark brown rings; quadrangular, both edges concave; prominent axial keel on left side, slightly grooved at extremities; surface smooth, with fine regular concentric rings. Anterior gizzard spines conical, sharp, at least twice the size of posterior spines. Posterior spines needle-like, usually curved; small spines present behind plates on right side.
Male genital system: ( Fig. 23C–D View Figure 23 ) Length 22–24 mm (H = 35.1, 38.4 mm). Vestibular area less than onethird overall length (excluding blind caecum), as wide as penial sheath. Prostate oval, only slightly wider than penial sheath. Blind caecum long and thin.
Hermaphroditic gonoduct: ( Fig. 24C–D View Figure 24 ) Length 32 mm (H = 38.4 mm). First part of membrane gland internal; second part external and smooth. Seminal receptacle small, either globose or cylindrical, of similar width to gametolytic duct. Gametolytic duct longer than common genital duct. Gametolytic gland larger than fertilization chamber. Fertilization chamber with complex network of ducts visible at surface. Vestibular area long.
Ecology: Little is known about the ecology of this species. Based on information from labels of museum specimens it lives subtidally down to 40 m deep. Bandel (1976) mentioned the presence of egg-masses attached to plants and rocks near the surface in brackish lagoons in Santa Marta, Colombia, where the species is reported to be common ( Diaz & Puyana, 1994).
Geographical distribution: ( Fig. 16 View Figure 16 ) It is restricted to the Caribbean Sea with a northern extension to the Florida Peninsula. It occurs on Cuba, on the mainland of Central America in Panama and Honduras, and in northern South America in Venezuela and Colombia.
Remarks: This seems to be an uncommon and overlooked species. It is hardly represented in Museum collections either as shells or wet material, and has seldom been referred to in the literature. The number of available specimens constrained the study of the variability of the shell and anatomical characters. Nonetheless, it was noticeable that shells are all very similar, and that the number and shape of rachidian cusps can vary in the same individual. Anatomical features of the digestive and reproductive systems of this species have been previously depicted by Marcus (1976) based on specimens from Colombia and largely agree with our findings.
Diaz & Puyana (1994) reported the occurrence of B. solida along the coast of Brazil, but we found no evidence for its presence south of Venezuela.
The similarities between this species and B. mabillei were discussed in the Remarks on the latter species. A synopsis of the morphological variability of shell and anatomical characters, and a comparison with other species, is given in Table 1.
EASTERN PACIFIC SPECIES
BULLA GOULDIANA PILSBRY, 1895 View in CoL
( FIGS 2F View Figure 2 , 4I–L, 5E View Figure 5 , 6H–I View Figure 6 , 17 View Figure 17 , 18E View Figure 18 –F, 19E–F View Figure 19 , 20E–F View Figure 20 , 21E–G View Figure 21 , 22E–F View Figure 22 , 23E–G View Figure 23 , 24E–G View Figure 24 )
Bulla nebulosa Gould View in CoL in Adams, 1850: 578, pl. 123, figs 79, 80 (types untraceble; neotype here selected, H = 48.2 mm, BMNH 20050360, Guaymas, Mexico, Fig. 4I herein; not Schröter, 1804). Menke, 1850: 162; 1854b: 44. Carpenter, 1857: 173. Sowerby, 1868: sp. 6, pl. 2, figs a–c. Mabille, 1896: 115. Bergh, 1900: 214, pl. 22, figs 24–29. Johnson, 1964: 115.
Bulla gouldiana Pilsbry, 1895: 340 View in CoL , pls 36, 48, figs 22–24, 15–17 (replacement name for B. nebulosa View in CoL ). Marcus, 1961: 4–5, pl. 1, figs 3–4. Johnson, 1964: 84. Marcus & Marcus, 1967: 143, 148. Abbott, 1974: 319, 001. Robles, 1975: 279, fig. 1. Eisenberg, 1981: 152. Stebbins, 1987: 95. Farfan & Ramirez, 1988. Mikkelsen, 1996: 436. Behrens & Hermosillo, 2005: 27, fig. 10. Gosliner & Williams, 2007: 785. Mclean, 2007: 751, 752, pl. 373, fig. j.
Bullaria gouldiana – Baker & Hanna, 1927: 127.
Bulla (Bulla) gouldiana View in CoL – Keen, 1971: 794, 235.
Bulla adamsi – Bergh, 1900: 216, pl. 22, fig. 30.
Taxonomic history: Adams (1850) described B. nebulosa View in CoL probably using some of A. A. Gould’s material in the collection of H. Cuming, and he attributed the authority of the species to Gould. Nevertheless it is clear that Gould himself never formally published the name ( Johnson, 1964). However, the name B. nebulosa View in CoL was preoccupied by Schröter (1804), who used it probably for an Australian species ( Pilsbry, 1895), although the description by Schröter is vague and type specimens are untraceable. Pilsbry (1895) proposed the replacement name B. gouldiana View in CoL , referring to Sowerby’s (1868) descriptions and figures of B. nebulosa View in CoL . Several shells from the H. Cuming collection in BMNH are possibly the type series used by A. Adams, and three of them (BMNH 20050360, 20050365) appear to have been figured by Sowerby (1868: sp. 6, figs a–c). To stablize the concept of this species a neotype is here designated, a shell that seems to be one illustrated by Sowerby (1868: sp. 6, pl. 2, fig. a) (ICZN, 1999: art. 75). Bergh (1900) identified specimens from the Gulf of California as B. adamsi , a nomen dubium introduced by Menke (1850) (see Taxonomic history of B. punctulata View in CoL ). Otherwise, the name B. gouldiana View in CoL has been in use throughout the 20th century.
Diagnosis: Shell: large, thin, globose, pale violet to brown. Jaws: elongated. Radula: rachidian cusps of similar size (except smaller marginal and central); inner and outer laterals of similar height; outer lateral cusps of same size; base with short denticle. Gizzard plates: pale reddish brown with a dark brown edge. Fine concentric darker rings present all over surface. Male genital system: vestibular area less than half overall length (excluding blind caecum); blind caecum short; prostate and penial duct coiled. Hermaphroditic gonoduct: vestibular area short; first part of membrane gland internal; seminal receptacle small; fertilization chamber with complex superficial structure. Range: from Santa Barbara, California, to Mazatlan, Mexico; Gulf of California. COI sequence: GenBank DQ986532 View Materials (BMNH20060112).
Material examined: California: San Luis Obispo , 1 sh, CAS 072860 About CAS , H = 46.1 mm, x.1985 ; San Diego , 2 sh, BMNH 1855.3 .14.37, H = 26.1–33.0 mm; San Diego, 5 sh, BMNH 20050362 , H = 36.2–48.1 mm . Mexico: 1 sh ( AMS c43474), H = 19 mm; Gulf of California , 6 sh, BMNH 20050365 , H = 38.2–45.6 mm; Guaymas , Sonora, 1 sh, BMNH 20050360 , H = 48.2 mm; El Cochore, Guaymas area , Sonora, 3 dis, CAS 067260 About CAS , H = 35.4–40.6 mm, iii.1958 ; Guaymas , Sonora, 2 sh, BMNH 20050361 , H = 44.4–48.4 mm; 2 km north of Boca del Tule to Arena Blanca , Baja California, 2 dis, CAS 101586 About CAS , H = 28.35, 28.7 mm, iv.1976 ; Estero de Punta Banda, south of Ensenada , Baja California, 4 sh, BMNH 20050364 , H = 35.3–45.2 mm; Estero de Punta Banda, south of Ensenada , Baja California, 6 sh, BMNH 20050363 , H = 20–36.1 mm ; Isla Conchas, San Ignacio Lagoon , Baja California, 2 sh, AMNH 157996 About AMNH , H = 30.0 mm, vii.1959 , tidal flat; Bahia de Magdalena , Baja California, 1 dis, BMNH 20050366 , H = 25.4 mm, dredged on Ulva spp. ; El Mostero , Baja California, 3 spc, BMNH 20050720 H = 12.4– 15.5 mm, vii.1994 .
Shell: ( Figs 4I–L, 5E View Figure 5 ) Maximum H = 48.4 mm. Large, thin, fragile, oval to ovate-quadrangular, wider anteriorly. Faint regular growth lines present; major growth interruptions occasionally present behind lip. Parietal callus thin, occasionally thick, partially covering spire aperture. Rear part of lip slightly folded inwards. Spire aperture wide; spire of six whorls, with raised striae; suture distinct or indistinct. Protoconch diameter 120–145 Mm. Background colour light brown to pale violet, with dark brown blotches shadowed with white, spread randomly over shell or merging to form irregular zigzag axial bands. Aperture showing shell pattern. Columella whitish brown. Lip pinkish brown. Periostracum thick, orange to dark brown.
Animal: ( Fig. 2F View Figure 2 ) Uniform yellowish orange, with large white blotches scattered all over body, particularly dense on the cephalic shield and lobes. Periocular area unpigmented.
Jaws: ( Fig. 6H–I View Figure 6 ): Elongate to boomerang-shaped. Triangular spatulate scales with smooth edges. Height 1.9–2.6 mm, width 0.3–0.8 mm (H = 25.4, 28.7 mm).
Radula: ( Figs 18E–F View Figure 18 , 19E–F View Figure 19 , 20E–F View Figure 20 ) In total, 21–29 rows (H = 28.7–33.9 mm), first 4–8 rows with rachidian teeth only. Rachidian teeth 667–784 Mm wide (H = 28.4, 33.9 mm); 10–14 cusps (usually 11); cusps triangular and sharp; central and last 1–2 marginal cusps smaller, others of similar size, occasionally decreasing outwards. Inner lateral teeth wide, with major central cusp symmetrically flanked on either side by 3–4 cusps of decreasing size; central cusp twice as wide; all cusps sharp. Outer lateral with five sharp cusps; outermost smaller; base with short denticle. Inner and outer laterals of same height.
Gizzard plates and spines: ( Figs 21E–G View Figure 21 , 22E–F View Figure 22 ) Plates 4.5–7 mm long by 1.5–2 mm wide (H = 25.4, 40.6 mm); pale reddish brown with a dark brown edge. Fine concentric darker rings present all over surface; quadrangular, both edges concave; prominent axial keel on left side, slightly grooved at extremities; surface smooth, with fine regular concentric rings. Anterior gizzard spines laterally flattened, at least twice the size of posterior spines. Posterior spines needle-like; slightly curved.
Male genital system: ( Fig. 23E–G View Figure 23 ) Length 29–32 mm (H = 28.35, 40.6 mm). Vestibular area about onequarter of overall length (excluding blind caecum); wider than penial sheath. Prostate cylindrical. Blind caecum short and thin. Prostate and penial duct coiled.
Hermaphroditic gonoduct: ( Fig. 24E–G View Figure 24 ) Length 33– 40 mm (H = 25.4, 33.9 mm). First part of membrane gland internal; second part external and smooth. Seminal receptacle small, globose, of similar width to gametolytic duct. Gametolytic duct longer than common genital duct. Gametolytic gland larger than fertilization chamber. Fertilization chamber with complex network of ducts visible at surface. Vestibular area short.
Ecology: This species lives in shallow water from the intertidal to 10 m deep in protected bays or estuaries, on sand and mud flats, sometimes with filamentous green and brown algae ( Marcus, 1961; Behrens, 1980; Behrens & Hermosillo, 2005; Neubig, 2006). It is active at night ( Marcus, 1961), but can also be seen crawling at dusk ( Neubig, 2006) and is more common during winter ( Marcus & Marcus, 1967). The life span is about 1 year ( MacGinitie & MacGinitie, 1949). In southern California (La Jolla) reproduction occurs between May and July ( Neubig, 2006).
Geographical distribution: ( Fig. 17 View Figure 17 ) It extends as far north as San Luis Obispo and Santa Barbara, California , southwards to the mouth of the Gulf of California, throughout the Gulf of California and to Mazatlan in Mexico .
Remarks: The anatomical descriptions by Pilsbry (1895), Bergh (1900), Marcus (1961) and Robles (1975) agree with the studied material and an account of the functioning of the hermaphroditic gonoduct can be found in the last of these.
The penial duct is in general coiled but can show straight parts. In the hermaphroditic gonoduct there is variation in the shape of the seminal receptacle. This structure is very small, sometimes almost imperceptible, usually globose, and an additional second pouch (as described in B. striata ) can be present.
The variation in shell shape (quadrangular to oval) is neither ontogenetic nor geographical, because both forms can be found in adults and juveniles, and in the same localities.
At first sight the more slender quadrangular shell form resembles that of B. punctulata , particularly because this latter species can have a similar colour pattern to B. gouldiana . These attributes can therefore lead to misidentification as B. punctulata . The references to this species south of Mazatlan (as far as Ecuador) are probably the result of this confusion. Despite the similarities between shells of B. gouldiana and B. punctulata , the former is always thin, light and fragile, whereas the latter is solid. However, the most striking differences are in the anatomy of the reproductive systems. Bulla gouldiana has a small blind caecum and coiled penial duct, whereas B. punctulata shows a developed blind caecum and uncoiled penial duct. Also, the female glands of the former species have a small seminal receptacle, whereas the later bears a larger receptacle. A synopsis of the morphological variability of shell and anatomical characters, and a comparison with other species, is given in Table 1.
BULLA PUNCTULATA ADAMS, 1850 View in CoL
( FIGS 2E View Figure 2 , 4M–S, 5F, H View Figure 5 , 6J–K View Figure 6 , 17 View Figure 17 , 18G–H View Figure 18 , 19G–H View Figure 19 , 20G–H View Figure 20 , 21H–J View Figure 21 , 22G–H View Figure 22 , 23H–I View Figure 23 , 24H–I View Figure 24 )
Bulla panamensis Philippi, 1848: 141 ( Panama; types lost; nomen dubium). Menke, 1850: 162. Pilsbry, 1895: 343.
Bulla quoyii Gray View in CoL in Adams, 1850: 576, pl. 123, fig. 71 (Galapagos Islands; types seen, 2 syntypes, BMNH 20060656, H = 12.1, 13.2 mm; not Gray, 1843). Pilsbry, 1895: 342, pl. 34, fig. 9.
Bulla quoyi – Sowerby, 1868: sp. 19, pl. 6 (not Gray, 1843). Mabille, 1896: 115 (not Gray, 1843).
Bulla rufolabris Adams, 1850: 577 View in CoL , pl. 123, fig. 76 (Galapagos Islands; types seen, 3 syntypes, BMNH 196975, H = 30.4, 29.7, 28.2 mm). Menke, 1854b: 44. Sowerby, 1868: sp. 17, pl. 6, figs a, b. 44. Mabille, 1896: 115.
Bulla (Bulla) rufolabris View in CoL – Keen, 1971: 794, 237.
Bulla punctata Adams, 1850: 577–578 View in CoL (misspelling of B. punctulata View in CoL , see footnote in Adams, 1850: 604, 607; not Schröter, 1804; not A. Adams in Sowerby, 1868 = B. mabillei ). Mabille, 1896: 115.
Bulla punctulata Adams, 1850: 577–578 View in CoL , 604, 607, pl. 123, fig. 77 ( Panama; lectotype here designated H = 33.2 mm, BMNH 196974/1, Fig. 4R herein; figured in Higo, Callomon & Goto, 2001: 141; paralectotype H = 27.2 mm, BMNH 196974/2, seen). Menke, 1854b: 44. Sowerby, 1868: sp. 8, pl. 3, figs a, b. Pilsbry, 1895: 341, pl. 36, figs 29–30, pl. 37, fig. 39. Stearns, 1893: 376. Bartsch & Rehder, 1939: 15. Sphon & Mulliner, 1972: 149. Morris, 1973: 204, pl. 62 (in part, includes B. gouldiana View in CoL ). Abbott, 1974: 319. Eisenberg, 1981: 152, pl. 134 (in part, includes B. gouldiana View in CoL ). Montoya, 1983: 343. Emerson, 1995: 18. Alamo & Valdivieso, 1997: 82, fig. 182. Hickman & Finet, 1999: 116, fig. 161. Paredes, Cardoso & Tarazona, 2004: 213. Valdés & Camacho-García, 2004: 488, fig. 7B. Behrens & Hermosillo, 2005: 28, fig. 11 (as B. punctuolata ).
Bullaria punctulata – Baker & Hanna, 1927: 127.
Bulla (Bulla) punctulata View in CoL – Keen, 1971: 794, 236.
Bulla aspersa Adams, 1850: 578 View in CoL , pl. 123, fig. 78 (Payta, Peru; holotype seen, BMNH 196969, H = 34.5 mm). Menke, 1854b: 44 (as B. adspersa ). Sowerby, 1868: sp. 18, pl. 6, figs a, b. Pilsbry, 1895: 341, pl. 36, figs 25–28. Mabille (1896): 115 (in part, includes B. ampulla View in CoL ).
Bulla adamsi Menke, 1850: 162 (Mazatlan, Mexico; nomen dubium). Carpenter, 1857: 172. Stearns, 1894: 158.
Bullaria quoyana – Dall, 1919: 300 (new name for B. quoyii Gray View in CoL in Adams, 1850; in part, includes B. gouldiana View in CoL ).
Taxonomic history: Several names have been given to this species mainly based on differences shown by the shells. The first was by Philippi (1848), referring to specimens from Panama. He did not explicitly mention on which coast of Panama the shells were collected and the description was vague. He emphasized the lack of spiral grooves (a feature present in the western Atlantic B. occidentalis ), so he could have beeen referring either to the Caribbean B. solida or to the eastern Pacific species. Menke (1850) and Pilsbry (1895) considered it to be the latter, but this cannot be confirmed because the types are lost, and we therefore consier this to be a nomen dubium.
Adams (1850) described and illustrated four species based on shells from Central and South America. He named shells from the Galapagos Islands ‘ B. quoyii Gray , MS’, presumably based on named material in the H. Cuming collection, and evidently being unaware that the name B. quoyii had previously been validly published, for an Australasian species, by Gray (1843). The original illustration depicts a juvenile shell, similar to the mainland shells of B. punctulata . A. Adams also attributed shells from the Galapagos Islands to a second species, which he named B. rufolabris . Although the shells from the Galapagos Islands show some unique features, they nevertheless appear to be conspecific with those occurring on the mainland (see Remarks below). The two other species, B. punctulata from Panama and B. aspersa from Peru, have similar shells. The name B. aspersa , apart from references by Sowerby (1868) and Pilsbry (1895), has been neglected, whereas the name B. punctulata has become widely used for the present species (see Synonymy above). Of these four species introduced by Adams (1850), B. punctulata is the third in the sequence of pagination but, acting as first revisers, and to promote taxonomic stability, we recommend its continued use (ICZN, 1999: art. 24).
Recently Valdés & Camacho-García (2004) listed B. punctata Adams, 1850 in the synonymy of B. punctulata , but erroneously indicated (by their BMNH registration numbers) the types of B. punctata A. Adams in Sowerby (1868), which are in fact specimens of B. mabillei (see Taxonomic history of that species). Surprisingly, they mentioned that these large rounded red shells of B. mabillei ‘ have a similar shell morphology ’ to the small, slender, brownish B. punctulata that they illustrated. The name B. punctulata has also been used incorrectly for B. orientalis from the Indo-West Pacific with shells that are very similar in shape and coloration (e.g. Angas, 1867; Fukuda, 1995; Hori, 2000).
The identity of B. adamsi Menke, 1850 is unclear. Menke introduced this name for shells from Mazatlan, Mexico, but at the same time identified this species as that figured by Adams (1850: pl. 123, figs 64–66) under the name B. australis Quoy & Gaimard, 1833 , based on shells from Tahiti that clearly do not match the features of any of the eastern Pacific species. Menke’s collection was sold and dispersed, and types are untraceable ( Dance, 1986). Carpenter (1857) and Pilsbry (1895) had already acknowledged this problem, and the latter suggested that B. adamsi should ‘ be dropped from the list of West American Bullas ’. However, some authors have still employed this name for specimens from the Indo- West Pacific (e.g. Dautzenberg, 1929; Eisenberg, 1981).
Diagnosis: Shell: small, thick, squat to pyriform, brown. Jaws: elongated. Radula: rachidian cusps of similar size (except marginal and central); inner and outer laterals of similar height; outer lateral cusps of same size; base with short denticle. Gizzard plates: yellowish with narrow marginal light brown band. Male genital system: vestibular area less than half overall length (excluding blind caecum); blind caecum long, thin; prostate coiled. Hermaphroditic gonoduct: vestibular area short. First part of membrane gland internal; seminal receptacle large; fertilization chamber with complex superficial structure. Range: Mazatlan, Mexico, southwards to Peru. COI sequence: GenBank DQ986556 View Materials (BMNH20030291).
Material examined: Mexico: Mazatlan, Sinaloa, 2 sh, BMNH 185910287 , H = 26.9, 33.5 mm; Anson Bay, I. Pedro Blanco (near Mazatlan ), 1 sh, BMNH 1848828– 26 , H = 19.50 mm; Puerto Vallarta , Jalisco, 1 sh, BMNH 20050645 , H = 20.6 mm, v.2005, washed ashore . Costa Rica: Cabo Blanco (San Miguel Laboratory), Nicoya Peninsula, 1 spc, BMNH 20030291 , H = 24.0 mm, 5 v.2003, intertidal rock pool with sand; Cabo Blanco (San Miguel), Puntarenas, 2 dis, INBio 01482898 View Materials , H = 16.0, 19.6 mm, 2 m depth; Junquillal, 200 m NW Playa Guaria, Parque Natural Santa Rosa , Guanacaste, 2 dis, INBio 03458490 View Materials , H = 13.8, 14.9 mm, iv.2002, 8 m depth . Panama: 4 sh, MNCN 15.05 About MNCN /33996, H = 27.6–38.0 mm; Coiba, 2 sh, BMNH 20050718 , H = 20.0, 23.1 mm; 2 sh, BMNH 1969741-2 , H = 27.2, 33.2 mm; Gulf of Panama, 4 spc, BMNH 20050721 , H = 10.7–12.5 mm . Ecuador: Punta de Santa Helena, 3 sh, MNCN 15.05 About MNCN /33995, H = 36.2– 38.4 mm. Manta-San Mateo Beach, 7 sh, BMNH 20050368 , H = 11.5–22.8 mm; La Libertad, 1 sh, AMS C.945568, H = 17.0 mm. Galapagos Islands: Academy Bay, Barranco , Isla de Santa Cruz, 3 dis, CAS 067270 About CAS , H = 13.8–18.1 mm, ii.1964; small islet, Academy Bay , 1 sh, AMNH 139496 About AMNH , H = 20.0 mm ; Isla San Cristobal, 18 sh, MNCN 15.05 About MNCN /17894, H ~ 23 mm ; Isla de Santa Fé, 10 sh, MNCN 15.05 About MNCN /17893, H ~ 26.4 mm . Peru: 4 sh, AMS C.88798, H = 28.0–31.0 mm; 2 sh, MNCN 15.05 About MNCN /33987, H = 34.7, 34.9 mm; north Peru, 5 sh, BMNH 20050367 , H = 16.4–22.0 mm .
Shell: ( Figs 4M–S, 5F, H View Figure 5 ) Maximum H = 38.4 mm. Small, thick, squat, ovate-quadrangular to pyriform. Growth lines present, fine. Spiral aperture wide; spire with 5–6 whorls with raised striae; sutures distinct. Protoconch diameter 125–150 Mm. Parietal callus thin. Background colour brownish to pinkish, with dark and white blotches scattered over shell, occasionally merging to form either zigzag axial or two spiral bands, one posteriorly and another between middle and anterior area of shell. Worn dead shells show a more conspicuous dotted pattern. Aperture grey, shell pattern visible through aperture. Columella white to pink; parietal callus white. Lip brown to pink. Periostracum brown, translucent.
Animal: ( Fig. 2E View Figure 2 ) Light brown to pale pink, with fine bright yellowish white spots spread over body. Periocular area unpigmented (for colour figures, see Hickman & Finet, 1999; Valdés & Camacho-García, 2004).
Jaws: ( Fig. 6J–K View Figure 6 ) Elongate; triangular spatulate scales with smooth edges. Height 1.0– 1.1 mm, width 0.3–0.4 mm (H = 17.6, 18.1 mm).
Radula: ( Figs 18G–H View Figure 18 , 19G–H View Figure 19 , 20G–H View Figure 20 ) In total, 22–28 rows (H = 14.9--17.6 mm), first 4–5 rows with rachidian teeth only. Rachidian teeth 364–423 Mm wide (H = 14.9–19.6 mm); 9–12 cusps; cusps triangular and sharp; central and marginal cusps smaller, other cusps of similar size, slightly decreasing outwards. Inner lateral teeth wide, with major central cusp symmetrically flanked on either side by three cusps of decreasing size; central cusp twice as wide; all cusps sharp. Outer lateral with five sharp cusps, outermost smaller; base with short denticle. Inner and outer laterals of same height.
Gizzard plates and spines: ( Figs 21H–J View Figure 21 , 22G–H View Figure 22 ) Plates 2.5–3.2 mm long by 0.8–0.9 mm wide (H = 16.0, 19.6 mm); wide marginal light brown band, central area yellowish; quadrangular, both edges concave; prominent axial keel on left side, slightly grooved at extremities; surface smooth, with fine regular concentric rings. Anterior gizzard spines conical, laterally flattened, either sharp or with rounded tips, about five times larger than posterior spines. Posterior spines needle-like, usually curved.
Male genital system: ( Fig. 23H–I View Figure 23 ) Length 12–17 mm (H = 18.1–19.6 mm). Vestibular area about one-third to half overall length of entire system (excluding blind caecum), wider than penial sheath. Prostate oval, conspicuously wider than penial region. Blind caecum long and thin. Prostate duct coiled.
Hermaphroditic gonoduct: ( Fig. 24H–I View Figure 24 ) Length 13–14 mm (H = 16.5, 19.6 mm). First part of membrane gland internal; second part external and smooth. Seminal receptacle large and globose, wider than gametolytic duct. Gametolytic duct longer than common genital duct. Gametolytic gland larger than fertilization chamber. Fertilization chamber with complex network of ducts visible at surface. Vestibular area short.
Ecology: This species occurs from the intertidal area down to 25 m deep, on sand and mud ( Keen, 1971; Hickman & Finet, 1999), and can be found in sandbottomed rock pools on intertidal platforms. It is primarily nocturnal ( Behrens & Hermosillo, 2005).
Geographical distribution: ( Fig. 17 View Figure 17 ) The northern limit of this species is Mazatlan, Mexico, and the southern limit northern Peru. It is likely to occur all along the coastline between these extremes where it is known from Costa Rica, Panama and Ecuador, and offshore on the Revillagigedo Islands (Socorro I), Cocos Islands and the Galapagos.
Remarks: Adult shells of this species can be very different in size and colour pattern, but apart from the Galapagos population there was no correlation between morphotype and geographical area. The colour pattern can resemble that of B. gouldiana (see Remarks on that species), although it is often more similar to B. occidentalis . Shell shape is usually quadrangular. However, the population from the Galapagos shows special features, with a smaller mean length of 20 mm, violet-pink shells, pyriform shape, conspicuous regular growth lines, usually spiral bands, and a pink or reddish lip. It would be interesting to test their conspecificity with molecular and anatomical data.
The radula is uniform among individuals and only seldom were the rachidian cusps of irregular shape and size.
There are several references to B. punctulata in Baja California (e.g. Baker & Hanna, 1927; Valdés & Camacho-García, 2004), which are probably due to similarities between this species and a particular morphotype of B. gouldiana (see Remarks on that species). Study of shells and anatomy failed to detect this species north of Mazatlan, Mexico. A synopsis of the morphological variability of shell and anatomical characters, and a comparison with other species, is given in Table 1.
Valdés & Camacho-García (2004) in their study of the Pacific cephalaspideans from Costa Rica seem to have mixed up shells from the Atlantic and Pacific sides. In fact the shell depicted as B. punctulata (p. 489, fig. 10E) probably belongs to B. occidentalis . Bulla punctulata does not have an elongate shape (whereas this is typical of B. occidentalis ), and importantly spiral striae on the base of the shell are present only in B. occidentalis , B. striata and B. quoyii (from Australasia).
INDO- WEST PACIFIC SPECIES
BULLA AMPULLA LINNAEUS, 1758 View in CoL
( FIGS 25A View Figure 25 , 26A–J, 28A–B View Figure 28 , 29A–B View Figure 29 , 30A–C View Figure 30 , 31A–C View Figure 31 , 32A–B View Figure 32 , 33A–D View Figure 33 , 35A–H View Figure 35 , 37A–C View Figure 37 , 38 View Figure 38 )
Bulla ampulla Linnaeus, 1758: 727 View in CoL View Cited Treatment (in part, includes B. solida ; Mauritius, Jamaica, Barbados, here restricted to Mauritius; probable types seen, LSL, lectotype here designated, H = 35.6 mm, 3 paralectotypes H = 25.5, 32.4, 54.8 mm; Fig. 26D herein; a fourth paralectotype is B. solida ). da Costa, 1776: 174, pl. 3, figs 4, 5. Bruguière, 1792: 371, pl. 358, fig. 3a, b. de Montfort, 1810: 330. Lamarck, 1822: 33. d’Orbigny, 1841: 478, pl. 45, fig. 12. Deshayes & Milne-Edwards, 1836: 668. Adams, 1850: 575, pl. 22, fig. 59–69. Hanley, 1855: 204. Sowerby, 1868: pl. 1, sp. 3a, b. Angas, 1877: 189. Watson, 1886: 637 (in part, includes B. arabica View in CoL ). Whitelegge, 1889: 275. Pilsbry, 1895: 343, pls 34, 48, figs 1–3, 4. Mabille, 1896: 111. Bergh, 1900: 210, pl. 28, figs 9– 25. Iredale, 1910: 71. Shirley, 1911: 102. Hedley, 1918: M104. Dautzenberg, 1929: 335. Habe, 1950: 21, pl. 3, figs 5, 15. Risbec, 1951: 143, fig. 9. Dodge, 1955: 23. Macnae, 1962: 185. Cernohorsky, 1972: 207, pl. 59, fig. 1, 1a. Kirtisinghe, 1978: 57, pl. 30, fig. 12. Lin & Qi, 1985: 115. Springsteen & Leobrera, 1986: 284, pl. 81, fig. 10. Gosliner, 1987: 45, fig. 21. Short & Potter, 1987: 116, fig. 6. Drivas & Jay, 1988: 130, pl. 50, fig. 12. Dharma, 1992: 76, pl. 16, fig. 20. Guangyu, 1997: 93, pl. 26, fig. 2. Apte, 1998: 85, pl. 11, fig. 12. Burn & Thompson, 1998: 957, fig. 16.36C. Steyn & Lussi, 1998: 176, 727. Higo, Callomon & Goto, 1999, G4895. Hori, 2000: 758, fig. 377. Dharma, 2005: 206, pl. 78, fig. 13a, b. Ekawa & Sakashita, 2005. Thach, 2005: 230, pl. 71, figs 7, 12.
Vesica ampulla – Swainson, 1840: 360.
Bullaria ampulla – Thiele, 1931 (1992): 384 (638), fig. 483.
Bulla ampulla striata Röding, 1798: 15 View in CoL (nomen dubium, not Bruguière, 1792; cited as B. ampullastriata in synonymy by Cernohorsky, 1972: 207).
Bulla ovum vanelli Röding, 1798: 15 (nomen dubium; cited as B. ovumvanelli in synonymy by Cernohorsky, 1972: 207).
Bulla fasciata Röding, 1798: 15 (nomen dubium).
Bulla bulla rubra Röding, 1798: 15 (nomen dubium).
Bulla flammea Röding, 1798: 16 (nomen dubium).
Bulla villosa Martyn, 1784 View in CoL : unnumbered, fig. 95 [right hand side] (invalid name, ICZN, 1957: opinion 456; China; types untraceable, Cabinet of Mr Hurlock ).
Bulla columellaris Menke, 1854a: 26 View in CoL ( Moluccas [ Indonesia]; types untraceable). Menke, 1854b: 43. Iredale, 1914: 667 (attributed to Menke, 1853). Hedley, 1916: 72.
Bulla columellaris var. bifasciata Menke, 1854a: 26 View in CoL ( Moluccas [ Indonesia]; types untraceable). Mabille, 1896: 112. Shirley, 1911: 102.
Bulla ampulla var. bifasciata View in CoL – Pilsbry, 1895, 344, pl. 34, fig. 5. Guangyu, 1997: 95, pl. 26, fig. 1.
Bulla columellaris var. vermiculata Menke, 1854a: 26 View in CoL (Moluccas [ Indonesia]; types untraceable).
Bulla trifasciata Sowerby, 1868 View in CoL : pl. 1, sp. 1 ( Philippine Islands; types seen, 2 syntypes BMNH 197651, H = 43.4, 43.5 mm; Fig. 26A–B, herein). Brazier, 1878: 83. Pilsbry, 1895, 344, pl. 34, fig. 4. Mabille, 1896: 112. Guangyu, 1997: 95, pl. 26, fig. 5.
Bulla delorti Mabille, 1896: 116 View in CoL ( Japan; 4 syntypes seen, MNHN; syntype figured by Valdés & Héros, 1998: 708, fig. 7A).
Bulla ampulla var. mozambicensis Bergh, 1900: 210 View in CoL , pl. 28, figs 22–24 (types untraceable; Mozambique).
·
Bulla adamsi – Dautzenberg, 1929: 334 (not Menke, 1850). Michel, 1985: 87, fig. 239 (not Menke, 1850).
Quibulla selina Iredale, 1929: 349 View in CoL , pl. 38, fig. 1 ( Sydney , Australia; types seen, holotype, H = 53 mm; four paratypes H = 47–53 mm, AMS C.57855).
Bulla cruentata View in CoL – Drivas & Jay, 1988: 130, pl. 50, fig. 13 (not Adams, 1850 = B. mabillei ).
Taxonomic history: This is the type species of the genus. The name B. ampulla View in CoL has been broadly applied to all Indo-Pacific specimens bearing a large and rounded shell, but it is here shown for the first time that such specimens include two species ( B. ampulla View in CoL and B. arabica View in CoL ). In the original description, Linnaeus (1758) gave as localities not only Mauritius in the Indian Ocean, but also Jamaica and Barbados in the western Atlantic (based on data from several earlier authors, e.g. Buonanni, 1684; Lister, 1685 –92; Gualtieri, 1742), and this led authors until the mid- 19th century to attribute a broader geographical range to the species (e.g. Bruguière, 1792; Lamarck, 1822; Deshayes & Milne-Edwards, 1836). From the second half of the 19th century the name B. ampulla View in CoL was generally used only for Indo-West Pacific specimens (e.g. Pilsbry, 1895; Bergh, 1900; Habe, 1950; Hori, 2000). The type lot of five shells in LSL consists of two species: four B. ampulla View in CoL and one B. solida (internally marked in pencil ‘ampulla’, probably by J. E. Smith, according to an annotation by S. P. Dance). Bulla solida View in CoL is restricted to the tropical western Atlantic. The problems of subsequent additions to the Linnean Collection, which hamper its use and the designation of types, have been discussed by Dance (1967). Therefore, to fix unambiguously the concept of this species, a lectotype is here designated (ICZN, 1999: article 74).
Martyn (1784) introduced the name Bulla villosa , but his work does not comply with the requirements of binominal nomenclature and is not valid for nomenclatural purposes (ICZN, 1957: opinion 456), except for nine species from New Zealand, which do not include B. villosa (ICZN, 1957: opinion 479).
The names introduced by Röding (1798) lack descriptions and no types can be traced. Röding referred to illustrations in Martini (1769) and Gmelin (1791), but those works potentially include two different species ( B. ampulla and B. arabica ), so it is impossible to determine the species intended by Röding.
Menke (1854a) introduced the name B. columellaris for specimens from the Moluccas, Indonesia. He described two varieties that match the main colour patterns present in B. ampulla from this region: shells with two conspicuous dark spiral bands ( var. bifasciata ) and those without bands ( var. vermiculata ). Sowerby (1868) named B. trifasciata based on two shells from the Philippines bearing three striking dark spiral bands. Mabille (1896) named shells from Japan B. delortii and the type material in MNHN is conspecific with B. ampulla . Iredale (1929) named Quibulla selina from Sydney Harbour; inspection of the types in AMS revealed these to belong to the bandless variety of B. ampulla .
Diagnosis: Shell: large, rounded, thick, pale to brownish or greenish, with axial zigzag stripes or dark spiral bands. Jaws: elongate to semicircular. Radula: central and marginal rachidian cusps of smaller size; outer lateral of similar height to inner, base with sharp denticle. Gizzard plates: broad dark brown marginal band. Male genital system: vestibular area one-quarter of overall length; blind caecum small, narrow. Hermaphroditic gonoduct: vestibular area short; first part of membrane gland internal; seminal receptacle cylindrical; fertilization chamber with complex network of ducts. Range: tropical Indo-West Pacific; Indian Ocean from South Africa to India and Andaman Sea, including Madagascar, Seychelles and Mascarene Islands; western Pacific from south-east Asia to southern Japan, Indonesia, Philippines, northern Australia, New Guinea and Fiji. COI sequences: GenBank DQ986520 View Materials –986523 (BMNH20060116/3; BMNH20060119; BMNH2006 01120; BMNH20060106/1); GenBank DQ986528 View Materials – 986529 (BMNH20030672/4; BMNH20050164/12).
Material examined: South Africa: Salt Rock, Umhlali, 3 dis, NM W2407, H = 38.3–38.7, in tide-pool; Knysna Lagoon, 7 sh, BMNH 20060526, H ~ 45 mm; Jeffreys Bay, 1 sh, BMNH 20060527, H = 24.3 mm; Algoa Bay, 3 sh, BMNH 1868.6.4.77, H = 21.6–25.7 mm; Port ·
Alfred, 2 sh, AMNH 147936 About AMNH , H = 55.0 mm. Mozambique: Fernão Veloso, Nacala Bay , 2 dis, BMNH 20060528 , H = 44.8, 47.8 mm; Bazaruto I., 1 dis, NM L6783 , H = 39.4 mm; 1 dis, BMNH 20060663 , H = 39.7 mm. Tanzania: Dar es Salaam, 2 sh, BMNH 20060529 , H = 45.7 mm; Dar es Salaam, 7 sh, BMNH 20060530 , H ~ 30.9 mm; Mnanzi Moja , Zanzibar, 2 sh, AMNH 135355 About AMNH , H = 42.0, 50.0 mm. Kenya: Mombasa, 9 sh, BMNH 20060532 , H ~ 37.7 mm; Shimoni , 1 dis, BMNH 20060531 , H = 40.4 mm; Shimoni , 1 sh, BMNH 20060533 , H = 35.9 mm. Madagascar: Anjahana, NW I. Ste Marie, 2 sh, BMNH 20030638 ; Libanona Beach , Taolagnaro , 1 dis, BMNH 20030672 /1, H = 47.0 mm, 3 m deep near shore, 17.x.2003; 8 sh, BMNH 20030635 , H ~ 43.5 mm. Mauritius: Tamarin , 1 dis, BMNH 20060580 , H = 35.1 mm, 24 October 1985. Ile Mayotte: 1 sh, BMNH 20060534 , H = 39.0 mm. Reunion: Cap Boucan Canot , 1 dis, NM K5607 , H = 22.6 mm. Seychelles: 1 sh, BMNH 19970929 , H = 37.0 mm; Mahé, 2 sh, BMNH 20060535 , H = 44.5 mm; Port Victoria , Mahé, 1 sh, BMNH 20060536 , H = 30.6 mm; Passe Hoasean , Aldabra , 1 sh, BMNH 20060537 , H = 27.8 mm. India: Tuticorin , Gulf of Mannar , 3 dis, BMNH 20050164 , H = 41.6–43.9 mm, iii.2003, 15 m depth in sandy mud; 1 sh, BMNH 20050164 /11, H = 38 mm; 2 sh, BMNH 20050165 , H = 52.5 mm; Port Blair , Andaman Islands , 6 sh, BMNH 1925.2.3.544–549, H ~ 34.4 mm; Nicobar Islands, 3 sh, BMNH 20060538 , H = 21.6 mm. Sri Lanka: Trincomalee , 2 sh, AMS C.107527. Burma, 1 sh, BMNH, 20060539, H = 37.0 mm. Thailand: Praphad beach, Kampuan , Ranong Province, 1 sh, BMNH 20070015 , H = 27.5 mm, intertidal sando, xi.2000. Malaysia: Semporna , Sabah, 1 sh, BMNH 20060545 , H = 35.4 mm. Vietnam: Cua Be , Nha Trang , 2 dis, BMNH 20010449 , H = 49.0 mm; Nha Trang , 2 dis, BMNH 20060106 , H = 45.0, 50.0 mm; Hon Kuoi , Nha Trang , 1 sh, BMNH 20041003 , H = 53.6 mm; 1 sh, BMNH 20041005 , H = 49.2 mm. China: SE Ping Chau I., Mirs Bay , Hong Kong, 2 sh, AMS C.103243; Chan Wan Bay , Hong Kong, 1 sh, AMS C.103553; Shai Han Bay , Lantau I., Hong Kong, 1 sh, AMS C.94490. Taiwan: Kaohsiung, 1 sh, BMNH 20050601 , H = 56.2 mm. Japan: Kagoshima, Hakamagoshi , 3 dis, BMNH 20060116 , H = 37.0– 42.6 mm, vi.2005. Philippines: 1 dis, BMNH 1867.5 .22.30, H = 39.9 mm; 2 sh, BMNH 20060540 , H = 32.5 mm; 4 sh, BMNH 20060541 , H ~ 46.4 mm; 2 sh, BMNH 20060542 , H = 23.9 mm; 3 sh, BMNH 20060543 , H = 62.3 mm; Zamboanga, Mindanao , 4 sh, MNCN 15.05 About MNCN /30304, H ~ 50 mm; Isla Basilan , 1 sh, MNCN 15.05 About MNCN /30307, H = 54 mm. Guam: 1 sh, RMNH. Indonesia: near mounth of Mahakam river , Kalimantan, 8 sh, AMS C.100818; southern Bali, 1 sh, RMNH; Bali, 3 sh, AMS C.60896; Moluccas, 3 sh, BMNH 20060544 , H = 33.0 mm; Ambon, 20 sh, RMNH; Ambon, 2 sh, RMNH. Papua New Guinea: Port Moresby, 1 sh, BMNH 20060546 , H = 48.9 mm; 2 sh, AMS C.58314; Poreporera , Port Moresby, 1 sh, AMS C.58038; Hula , 2 sh, AMS C.58022; Hisiu , 2 sh, AMS C.76484; Haidaio , 1 sh, AMS C.58278; Arona , 6 sh, AMS C.58264; Kapukapu , 4 sh, AMS C.58251; New Britain, 1 dis, ZMB 38888, H = 33.6 mm. Solomon Islands: 2 sh, BMNH 1920.8.20.9-10, H = 36.9, 38.2 mm; 1 sh, BMNH 20060547 , H = 38.5 mm; 3 sh, AMS 85086, H ~ 36 mm. New Caledonia: Illot Tangadiou , Koumac (20°33.5′S, 164°13′E), 5 sh, MNHN, Paris, H ~ 49.0 mm, October 1993; Récif de l’Infeernet , Koumac (20°37.1′S- 164°14.7′E), 1 sh, MNHN, Paris, H = 51.2 mm, 1–5 m deep, October 1993; Nouméa (dredged outer lagoon), 1 dis, BMNH 20060548 , H = 58.7 mm; 1 sh, BMNH 19990828 , H = 46 mm; Baie des Citrons , Noumea, 2 sh, BMNH 20060549 , H = 35.7 mm; Ovemo, Magenta, Baie des Isoles , Nouméa, 3 dis, BMNH 20060550 , H = 38.0– 40.7 mm; NW Ile Ste Marie, Nouméa, 4 sh, AMS C.86252; N. Duvéa I., 4 sh, AMS C.116809. Fiji: Darvasamu, Tailevu, 1 sh, BMNH 20060551 , H = 50.2 mm; Suva, Viti Levu, 1 sh, BMNH 20060552 , H = 50.0 mm; Nadi Bay , Viti Levu, 10 sh, AMS C.68302; Ovalau, 1 sh, BMNH 1871.3 .29.16, H = 44.6 mm. Tahiti: 7 sh, BMNH 20060553 , H ~ 52.2 mm; 2 sh, BMNH 20060554 , H = 41.3 mm. Australia: Bundegi Reef, North-west Cape, WA, 2 dis, WAM S19141 View Materials , H = 37.0, 38.4 mm; Wallabi Group, Houtman Abrolhos Islands , WA, 1 dis, WAM S19151 View Materials , H = 45.5 mm; WA, Exmouth, south of NW Cape, Yardie Creek , WA, 1 dis, AMS C.444877, H = 44.6 mm; N end of Bay N of Cloates, WA, 2 dis, WAM S19144 View Materials , H = 41.8, 44.0 mm; Montebello I., WA, 1 sh, AMS C.49704; Little Lagoon, Shark Bay , WA, 2 sh, AMS C.122229; Notch Point, Dirk Hartog I., Shark Bay , 1 dis, WAM S19152 View Materials , H = 52.5 mm; Ninga- loo, S. of North-west Cape, WA, 2 sh, AMS C100494 ; Turtle Beach, W. of North-west Cape, WA, 2 sh, AMS C95194; Shoal Flats, WA, 1 dis, WAM S19127 View Materials , H = 53.4 mm; Croker Island , Cobourg Peninsula, NT, 3 sh, AMS C.77615, H ~ 35 mm; Smith Point, NT, 1 sh, AMS C.104021; Telegraph Stn area , Cape York, Qld, 4 sh, AMS C.94590; Long Island , Torres Strait, Qld, 2 sh, AMS C.107534; Stradbroke I. (27°35′S- 153°28′E), southern Qld , 1 sh, AMSc.013062, H = 5 1 mm, outer beach, ix.1902; Crescent Head Beach (31°11.5′S, 152°58.8′E), NSW, 2 sh (fragments), AMS C.88735, H = 49, 51 mm, viii.1959; Port Stephens , NSW, 1 sh, AMS C.88781, H = 44 mm, collected 1877; Sydney Harbour (33°51′S, 151°14′E), NWS, 4 sh, AMS C.88732, H = 42–56 mm, ‘ Triton’ dredge 1928–29; Sydney Harbour (33°51′S, 151°14′E), NSW, 4 sh, AMS C.88736, H = 46–53 mm, ‘ Triton’ dredge 1928–29; Sydney Harbour (33°51′S, 151°14′E), NSW, 1 sh, AMS C.107532, H = 48 mm; Sydney Harbour (33°51′S, 151°14′E), NSW, 1 sh, AMS C.94556, H = 38 mm, ‘ Triton’ dredge 1928–29; Sydney Harbour (33°51′S, 151°14′E), NSW, 1 sh, AMS C.88737, H = 53 mm, ‘ Triton’ dredge 1928–29; Sydney Harbour (33°51′S, 151°14′E), NSW, 4 sh, AMS C.88732, H = 42–56 mm, ‘Triton’ dredge 1928– 29 GoogleMaps .
Shell: ( Figs 26A–J, 28A–B View Figure 28 ) Maximum H = 62.3 mm, thick (thin in shells up to H = 30 mm), shape ovatequadrangular to rounded. Fine growth lines all over shell; major growth interruptions occasionally present behind lip. Parietal callus usually thin. Posterior part of lip can be slightly folded inwards in adults. Spire aperture wide; spire 5–7 whorls, with raised striae. Protoconch diameter 135–170 Mm. Background varies from white to pale yellowish brown, finely mottled with brown dots scattered over shell or dark blotches, often forming zigzag axial stripes, alternatively with two or three narrow dark spiral bands; darkest shells brownish green with two or three darker spiral bands. Aperture white, shell pattern sometimes visible; lip white to pale brown; columella white. Periostracum translucent.
Animal: ( Fig. 25A View Figure 25 ) Pale brown with few white dots scattered over surface. Central area of cephalic shield dark brown; occasionally specimens show a dark band across the cephalic shield between the eyes and the anterior edge. Periocular area unpigmented. Foot pale yellowish brown; white pigment absent (description based on specimens from southern India).
Jaws: ( Fig. 29A–B View Figure 29 ) Elongate to semicircular shape. Triangular spatulate scales with smooth rounded edges. Height 1.3–3.0 mm, width 0.4–0.9 mm (H = 33.6–47.0 mm).
Radula: ( Figs 30A–C View Figure 30 , 31A–C View Figure 31 , 32A–B View Figure 32 ) In total, 24–31 rows (H = 36.0–50.0 mm), first four or six rows with rachidian teeth only. Rachidian tooth 645–1000 Mm wide (H = 36.0, 53.4 mm); 9–13 (usually 11) cusps; cusps triangular, sharp, usually last marginal cusp of smaller size, central cusp minute. Inner lateral tooth with large central cusp, longer and twice as wide as others, symmetrically flanked by 3–4 cusps of decreasing or similar size; cusps sharp. Outer lateral tooth with 5–6 cusps; innermost largest, others of similar size; outermost smallest; cusps sometimes bifid or trifid, base with sharp denticle, smaller than outermost cusp.
Gizzard plates and spines: ( Fig. 33A–D View Figure 33 ) Plates 5.4– 8.1 mm long by 2.1–2.9 mm wide (H = 33.6–58.7 mm); yellowish brown or reddish, with broad dark brown margin; quadrangular to oval, left edge either slightly concave, rectilinear or convex, right concave; top flat or angled; surface smooth, occasionally with fine regular concentric rings; axial keel on left side, prominent or absent, keel extremities reduced or expanded, occasionally grooved. Anterior gizzard spines conical or laterally flattened, usually with large globular base, sharp, twice width of posterior spines. Posterior spines needle-like or triangular, small but variable in size, large globular base.
Male genital system: ( Fig. 35A–H View Figure 35 ) Length 15–45 mm (H = 22.6, 50.0 mm). Vestibular area about onequarter overall length of system (excluding blind caecum), usually narrower than penial sheath; penial sheath thick, muscular. Prostate cylindrical, inflated, sometimes globular. Blind caecum small, narrow.
Hermaphroditic gonoduct: ( Fig. 37A–C View Figure 37 ) Length 36–56 mm (H = 33.6, 49.0 mm). First part of membrane gland internal; second part external, smooth. Seminal receptacle small, cylindrical. Gametolytic duct longer than common genital duct; apical area globular. Gametolytic gland four times size of fertilization chamber. Fertilization chamber with complex network of ducts visible at surface. Vestibular area small.
Ecology: This species occurs in sheltered areas in habitats of sand or mud and seaweeds from the intertidal down to 15 m deep. They are active at night and breed during July when egg masses and mating were observed in the mid-intertidal zone on sandy flats with seagrass in sheltered bays in northern Mozambique (D. G. Reid, pers. comm.). An image of a living specimen from South Africa can be found in Gosliner (1987).
Geographical distribution: ( Fig. 38 View Figure 38 ) This is mainly a tropical species that extends its distribution into the warm temperate areas of South Africa and southern Japan. Occurs along the coast of East Africa from South Africa, Mozambique to Kenya; in Madagascar and the islands of Mauritius, Seychelles and Reunion; to southern India and Sri Lanka, Burma and Andaman Islands; western Malaysia, Vietnam, China, Taiwan; southern Japan; Indonesian islands of Borneo, Java, Bali, Sulawesi and Moluccas; Philippines and Guam; western and northern Australia from Shark Bay to Cape York, New Guinea and the Coral Sea, to the Solomon Islands, New Caledonia and Fiji.
Remarks: The main differences between B. ampulla and its sister species B. arabica lie in the dimensions, coloration and shape of the shell, shape of the gizzard plates and male genital system (see Remarks on B. arabica ). Moreover, the molecular phylogeny supports their separation and sister relationship.
Variability of the shells and anatomy of B. ampulla has been found across its distribution. Shells from eastern Africa and India are more oval and are usually marked with zigzag axial stripes, whereas those from continental south-east Asia tend to be paler and finely mottled in brown; north-west Australian shells are whitish, but in New Caledonia they again show the pattern of zigzag stripes. A distinct greenish form with two to three conspicuous dark spiral bands co-occurs with the more conventional colour forms in southern Japan, Indonesia, New Guinea and Solomon Islands. In addition, western Pacific shells are in general more rounded. Up to 30 mm in length the shells are usually thin and the animals are immature. Above this length shells tend to be thick and solid. However, thin shells were found in adult specimens from South Africa.
The variation found in the shape of the jaws could be caused by artefacts of fixation. Direct fixation without previous relaxation of the animal results in strong contraction of the body, which might cause internal structures to distort. Both elongate and semicircular shapes have been depicted by Bergh (1900) and Ekawa & Sakashita (2005) for specimens from eastern Africa and Japan, respectively.
Larger specimens can show gizzard plates with a flat top and small keels, whereas plates of juveniles or young adults have an angled top with a prominent keel. The action of the plates during digestion when they crush and mince food eventually causes them to become worn. These extreme shapes correspond to those previously described in this species by Bergh (1900; East Africa) and Ekawa & Sakashita (2005; Japan).
In the male genital system, the sheath enveloping the prostate and penial duct is thick and muscular, particularly in the prostate region. However, specimens from New Caledonia had a thin and translucent sheath in the penial region. Additionally, some western Pacific specimens showed a distinct texture in the apical region of the prostate sheath. This might be linked with fixation or amount of sperm storage in the ducts. Occasionally, the apical region of the penis can protrude and either the end of the vestibular area or the beginning of the penial region expands to become wider than the remaining structure. Variability was also detected in the coiling of the prostate and penial ducts. The prostate can either be coiled in a ball near the distal part of the system or be represented by a straight duct contiguous with the blind caecum and with coiling limited to a lower cylindrical portion; the former situation is much more common. Usually the penis is a straight duct that occasionally coils along its length. The apical tip of the penis is wider and wrinkled.
The differences found in the fertilization chamber of the hermaphroditic gonoduct, where some specimens showed a network of narrow ducts with tissue filling the space in between and other wider ducts that are inflated and pressed against each other, are possibly the result of the amount of sperm or ova inside the ducts and not a distinct anatomical feature. The same applies to the visible part of the membrane gland that can vary from inconspicuous to a wide white duct.
The only previous representation of the reproductive system of this species was by Risbec (1951) based on a specimen from New Caledonia, but both the male and the hermaphroditic gonoducts were incomplete.
The features of the radula of B. ampulla match those described by previous authors ( Pilsbry, 1895; Bergh, 1900; Thiele, 1931; Habe, 1950; Ekawa & Sakashita, 2005).
Despite the variability found in the various characters no relation could be established with a shell morphotype or geographical region. Nevertheless, molecular data suggest a potential segregation between eastern Africa and India plus south-east Asia. Specimens from the central regions of the western Pacific and northern Australia were not available for sequencing. The banded shell morphotype is quite distinctive and only one specimen was obtained for dissection. When more specimens representing the entire geographical range and all morphotypes become available for dissection and molecular phylogenetic analysis, the systematics of B. ampulla might require reappraisal. A synopsis of the morphological variability of shell and anatomical characters, and a comparison with other species, is given in Table 2.
The records of this species in New South Wales and southern Queensland are based on material dredged from subtidal deposits ( Iredale, 1929) or washed ashore, and these shells are probably subfossil (I. Loch, pers. comm.). Populations of B. ampulla are otherwise unknown from the entire coast of eastern Australia .
BULLA ARABICA SP. NOV.
( FIGS 25B View Figure 25 , 26K–N, 28C View Figure 28 , 29C View Figure 29 , 30D View Figure 30 , 31D View Figure 31 , 32C View Figure 32 , 33E–F View Figure 33 , 35I–K View Figure 35 , 37F–G View Figure 37 , 38 View Figure 38 )
Bulla ampulla View in CoL – Cooke, 1886: 130. Watson, 1886: 637 (in part, includes B. ampulla View in CoL ). Melvill & Standen, 1901: 456. Vayssière, 1906: 18, pl. 2, figs 27–33. Eales, 1938: 81, fig. 2 (in part, includes B. ampulla View in CoL ). Bosch & Bosch, 1982: 137. Sharabati, 1984: pl. 34, fig. 8. Barash & Danin, 1992: 186, fig. 199. Coulombel, 1994: 112. Bosch et al., 1995: 179, fig. 823. Zenetos et al., 2003: 172. Mienis, 2004: 33. (All not Linnaeus, 1758.)
Bulla ampulla View in CoL var. M. rubri Bergh, 1900: 210 , pl. 28, figs 18–20.
Bullaria ampulla – Green, 1994: 71, figured (not Linnaeus, 1758).
Types: Ras al-Khaimah, United Arab Emirates (25°49′30″N, 55°58′30″E); holotype GoogleMaps , BMNH 20060103 /1, H = 39.3 mm; Fig. 26L; 4 paratypes , BMNH 20060103 , H = 30.5–38.8 mm .
Etymology: From its geographical occurrence in the Arabian Peninsula.
Taxonomic history: This new species has hitherto been included under B. ampulla . The uncertainty over the names of Röding (1798) has already been discussed in the Taxonomic history of B. ampulla . Bergh (1900) introduced the varietal name ‘var. M. rubri ’ for a single examined specimen from the Red Sea, but this appears to refer only to its origin and not to a comparative diagnosis, so we consider this to be infrasubspecific and therefore unavailable (ICZN, 1999: art. 45.6).
Diagnosis: Shell : large thick, ovate-quadrangular, brown mottled with darker blotches. Jaws : semicircular. Radula : rachidian cusps similar in size (central and marginal smaller); outer lateral of similar high to inner; base with sharp and developed denticle. Gizzard plates: broad dark brown marginal band, base wider than plate. Male genital system: vestibular area one-quarter of overall length; blind caecum small and globular. Hermaphroditic gonoduct: vestibular area short; first part of membrane gland internal; seminal receptacle cylindrical; fertilization chamber with complex superficial structure. Range : Red Sea , Persian Gulf and Gulf of Oman. COI sequences: GenBank DQ986516 View Materials ( BMNH 20060102 ); GenBank DQ986517 View Materials ( BMNH 20060103 ) .
Material examined: Red Sea, 1 dis, ZMB 789 View Materials , H = 25.5 mm. Egypt: Nuweiba, Sinai, Gulf of Aqaba , 1 dis, BMNH 20060555 , H = 26.4 mm; Gulf of Suez, 1 sh, BMNH 20060556 , H = 38.7 mm. Sudan: 2 sh, BMNH 20060557 , H = 27.5 mm; 2 sh, BMNH 20060558 , H = 37.2 mm; Port Sudan Harbour , 3 sh, BMNH 20060559 , H = 21.6 mm. Saudi Arabia: Bal Half Mukalla , 3 sh, BMNH 20060560 , H = 26.4 mm. Yemen: Aden, 12 sh, BMNH 20060561 , H ~ 30.1 mm; Aden, 1 sh, BMNH 19021230670 , H = 30.4 mm; Aden, 3 sh, BMNH 20060562 , H = 36.2 mm; Aden, 1 sh, BMNH 20060563 , H = 42.7 mm; Aden, 1 sh, BMNH 20060564 , H = 30.6 mm. Oman: Khasab , 1 dis, BMNH 20060565 , H = 35.9 mm; Muscat, 2 sh, BMNH 20060566 , H = 29.1 mm; Muscat, 5 sh, BMNH 20060567 , H ~ 22.9 mm; Muscat, 2 sh, BMNH 20060568 , H = 36.8 mm. United Arab Emirates: Khor Julfar , Ras al-Khaimah, 2 dis, BMNH 20060101 ; Khor Julfar , Ras al-Khaimah, BMNH 20060102 , H = 42.2, 39.6 mm, 19 iii.2004, low tide on seagrass; Jebel Dhanna, Abu Dhabi, 1 sh, BMNH 20060571 , H = 34.0 mm. Pakistan: Karachi, 1 sh, BMNH 20060569 , H = 34.3 mm; 3 sh, BMNH 20060570 , H = 28.0 mm. Kuwait: 1 sh, BMNH 20060660 , H = 22.7 mm .
Shell: ( Figs 26K–N, 28C View Figure 28 ) Maximum H = 42.2 mm, thick, shape ovate-quadrangular. Growth lines all over shell; major growth interruptions occasionally present behind lip. Parietal callus thin; columella thick. Spire aperture wide; spire 6–7 whorls, with raised striae. Protoconch diameter 130–140 Mm. Background colour whitish to brown, mottled with darker blotches that can merge to form faint spiral bands. Aperture white with shell pattern visible; lip brown; columella white.
Animal: ( Fig. 25B View Figure 25 ) Orange-yellow with bright white dots scattered over cephalic shield, cephalic and parapodial lobes and foot.
Jaws: ( Fig. 29C View Figure 29 ) Semicircular shape. Triangular spatulate scales with smooth rounded edges. Height 1.5–2.4 mm, width 0.5–0.9 mm (H = 26.4–39.6 mm).
Radula: ( Figs 30D View Figure 30 , 31D View Figure 31 , 32C View Figure 32 ) In total, 23–24 rows (H = 26.4–39.6 mm), first three rows with rachidian teeth only. Rachidian tooth 476–784 Mm wide (H = 26.4, 39.6 mm); 11 (occasionally 12) cusps; cusps triangular, sharp, sometimes bifid, usually last marginal cusp of smaller size, central cusp minute. Inner lateral tooth with large central cusp twice as long and wide, symmetrically flanked by 3–4 cusps of decreasing size; cusps generally sharp. Outer lateral tooth with five cusps; innermost largest; outermost smallest; cusps sometimes bifid, base with large sharp denticle.
Gizzard plates and spines: ( Fig. 33E–F View Figure 33 ) Plates 4.0– 6.7 mm long by 1.4–2.4 mm wide (H = 26.4–39.6 mm); dark brown with broad dark brown margin; quadrangular, left edge either slightly concave, straight or convex, right concave; top angled; surface smooth or with fine regular concentric rings; axial keel on left side, keel extremities prominent, grooved; base rounded, larger than plate. Anterior gizzard spines large, conical, sharp, with rounded base. Posterior spines small, needle-like, triangular, with globular base.
Male genital system: ( Fig. 35I–K View Figure 35 ) Length 8–25 mm (H = 26.4, 39.6 mm). Vestibular area about onequarter to one-third overall length of system (excluding blind caecum). Prostate globular to cylindrical, half length of penial sheath. Blind caecum small, usually globular. A penial pouch near the vestibular area sometimes present.
Hermaphroditic gonoduct: ( Fig. 37F–G View Figure 37 ) Length 41– 56 mm (H = 38.3, 49.0 mm). First part of membrane gland internal; second part external, wide and smooth. Seminal receptacle cylindrical. Gametolytic duct longer than common genital duct. Gametolytic gland four times size of fertilization chamber. Fertilization chamber with complex network of narrow ducts visible at surface. Vestibular area small.
Ecology: Little is known about the ecology of this species. In the southern Persian Gulf it has been found in the low intertidal of sheltered lagoons in habitats with seagrass where it was abundant during March and appeared to be most active nocturnally (G. R. Feulner, pers. comm.). In the north of the Red Sea it was observed in shallow waters down to 25 m deep in reefal habitats with seagrass, and in the Gulf of Eilat ( Israel) it breeds in April laying eggs on green algae at a depth of 25 m ( Koretz, 2005).
Geographical distribution: ( Fig. 38 View Figure 38 ) This species is restricted to the Red Sea and Arabian Peninsula. It occurs throughout the Red Sea from the Gulfs of Suez and Aqaba, to Sudan and Djibouti; it is recorded from Yemen, Oman and as far as Karachi in Pakistan, and in the Persian Gulf from Kuwait, Bahrain and the United Arab Emirates.
Remarks: Although similar in external appearance to B. ampulla several features separate these two species. Bulla arabica is smaller (max. H ~ 40 mm) than B. ampulla (max. H ~ 60 mm) and the adult stage is reached at H = 25–35 mm. Additionally, shells are generally more square and paler than the eastern African type of B. ampulla . The coloration of the cephalic shield is orange-yellow dotted with white and without any darker region of brown as observed in specimens of B. ampulla from southern India. Anatomically, the gizzard plates of this species have a large rounded base that exceeds the width of the plates, whereas the base in B. ampulla is no wider than the plate. This feature, previously depicted by Vayssière (1906: pl. 2, fig. 31), is not found in any other species and seems therefore to be a synapomorphy of B. arabica . Also, in the male genital system the penial sheath is thin and translucent in comparison with the usually thick muscular condition in B. ampulla ; in addition, the blind caecum is smaller and generally globular rather than a small narrow cylindrical structure as in B. ampulla . Genetic data ( Fig. 40 View Figure 40 ) also support their separation, with minimum genetic distances between the two species (uncorrected p -distances) of 10.5% for the COI gene (see Discussion).
The drawings by Bergh (1900) of the outer laterals of this species (as B. ampulla var. M. rubri ) agree in general with the variability found in the studied specimens, although he depicts outer laterals with more numerous cusps (6–7). Variability in the number of cusps in the lateral teeth is common in Bulla . A synopsis of the morphological variability of shell and anatomical characters, and a comparison with other species, is given in Table 2.
The apparent absence of this species from the southern coast of the Arabian Peninsula (Arabian Sea coast) may be due to the paucity of suitable environments. Shells of this species (as B. ampulla ) have been found in the eastern Mediterranean on the coasts of Israel, Greece, Cyprus and Turkey ( Barash & Danin, 1982, 1992; Dalanotte & Vardala- Theodorou, 2001; Zenetos et al., 2003; Mienis, 2004), but so far the occurrence of living specimens has not been reported. This species might be an occasional migrant into the Mediterranean, where stable populations have not yet been established.
BULLA ORIENTALIS HABE, 1950 View in CoL
( FIGS 25E View Figure 25 , 26O–R, 28D View Figure 28 , 29D View Figure 29 , 30E View Figure 30 , 31E View Figure 31 , 32D View Figure 32 , 33G–H View Figure 33 , 36A–C View Figure 36 , 37D–E View Figure 37 , 39 View Figure 39 )
Bulla punctulata View in CoL – Angas, 1867: 226. Brazier, 1879: 198. Melvill & Standen, 1899: 155. Hedley, 1907: 486. Hedley, 1918: M104. Cernohorsky, 1972: 207, pl. 59, fig. 5. Salvat & Rives, 1975: 177. Short & Potter, 1987: 116, fig. 4. Fukuda, 1995: 31, fig. 1039. Guangyu, 1997: 93, pl. 8, fig. 1. Higo et al., 1999: G4895. Hori, 2000: 758, pl. 377, fig. 3. Nakano, 2004: 30, figured. Cobb & Willan, 2006: 30, figured. (All not Adams, 1950.)
Bulla punctata View in CoL – Whitelegge, 1889: 275. Melvill & Standen, 1897: 396 (not Adams, 1950).
Bulla adamsi – Dautzenberg, 1929: 334 (not Menke, 1850). Eisenberg, 1981: 152, fig. 16 (not Menke, 1850).
Bulla peasiana View in CoL – Kuroda, 1941: 133, pl. 3, fig. 47 (not Pilsbry, 1895). Kuroda & Habe, 1952: 42 (not Pilsbry, 1895).
Bulla orientalis Habe, 1950: 21 View in CoL , pl. 3, fig. 10 (Karenko, Formosa [ Taiwan]; holotype in Geological Institute Taihoku Imperial University [ Taiwan National University] No. 1431 and paratype in National Science Museum, Tokyo NSMT-Mo 54561; not seen). Kira, 1962: 114, pl. 40, fig. 12. Burn, 1966: 97, figs 5–8.
Taxonomic history: The taxonomic history of this species has been confused by a number of misidentifications. Presumably because of the dotted pattern of the western Pacific specimens, the name B. punctulata Adams, 1850 View in CoL has been often been used for this species (e.g. Angas, 1867; Hedley, 1918; Hori, 2000). However, this name is correctly applied to an eastern Pacific species (see Taxonomic history of B. punctulata View in CoL ). Specimens from East Africa have been misidentified as B. ampulla . It can indeed be difficult to separate juveniles of B. ampulla from adults of the East African form of B. orientalis View in CoL , so that literature references to B. ampulla from Africa cannot with certainty exclude B. orientalis View in CoL . In Japan the name B. peasiana View in CoL , correctly applied to a Hawaiian species, has sometimes been used ( Kuroda, 1941; Kuroda & Habe, 1952).
The name B. adamsi is controversial because Menke (1850) introduced it for an eastern Pacific species while referring to an illustration of a Tahitian shell in Adams (1850) (see Taxonomic history of B. punctulata ).
The only available name for this species is therefore B. orientalis Habe, 1950 .
Diagnosis: Shell: small, thick or thin, slender or quadrangular, reddish brown with spiral bands or scattered with dark dots. Jaws: semicircular. Radula: with rachidian cusps of decreasing size (central and marginal smaller); outer lateral of similar size to inner; base with sharp denticle. Gizzard plates: with broad brown marginal band. Male genital system: vestibular area one-quarter of overall length; prostate rounded; blind caecum large and narrow. Hermaphroditic gonoduct: vestibular area short; first part of membrane gland internal; seminal receptacle large and cylindrical; fertilization chamber with complex structure. Range: Indo-West Pacific; from South Africa to southern Japan; north-east Australia, New Guinea to Fiji. COI sequences: GenBank DQ986535 View Materials – 986536 (BMNH20040859; BMNH20030672/2).
Material examined: South Africa: Algoa Bay , 50 sh, BMNH 20060572 , H ~ 27.8 mm. Tanzania: Dar es Salaam, 1 sh, BMNH 19920741 , H = 23.5 mm; 2 sh, BMNH 20060574 , H = 31.5 mm. Madagascar: Taolagnaro , 1 dis, BMNH 20030672 /2, H = 27.8 mm, 17.x.2003, 3 m depth near shore. Seychelles: Mahé, 2 sh, BMNH 20060575 , H = 17.7 mm; Mahé, 1 sh, BMNH 20060576 , H = 20.7 mm. Mauritius: 15 sh, BMNH 20060577 , H ~ 28.5 mm; 8 sh, BMNH 20060578 , H ~ 31.2 mm; 4 sh, BMNH 20060579 , H ~ 31.1 mm. Maldives: Gan-Fedu Gap, Gan, Addu Atoll , 2 sh, BMNH 1966513 , H = 24.7 mm. India: Port Blair, Andaman Islands , 1 sh, BMNH 20060581 , H = 26.6 mm. Vietnam: Nha Trang Harbour , 1 sh, AMNH 307092 About AMNH , H = 30 mm. Japan: Kyoda , Okinawa, 1 dis, BMNH 20040859 , H = 22.0 mm, vi.2004, Zostera bed, 2–3 m depth; Ryukyu Islands , 1 sh, BMNH 20060582 , H = 21.6 mm. Australia: Christmas I., 2 sh, AMS C.152467, 1983; Cocos I., 1 sh, AMS C.436599, 1979; Taylor Reef, Qld , 1 sh, AMNH 215098 About AMNH , H = 28 mm; Mackerel Bay, Hook I., Qld , 2 sh, BMNH 20060583 , H = 22.5 mm. Low Isles, Qld, 3 sh, AMS C.72621; Low Isles, Qld, 14 sh, AMS C.76027; North Keppel I., Qld, 1 sh, AMS C.94602; Swain Reefs, Qld, 8 sh, AMS C.436387; Murray I., Torres Strait, Qld, 6 sh, AMS C.29593; Frederick Reef, Coral Sea, Qld, 5 sh, AMS C.329419; Coral Sea, Carola Cay, Marion Reef, Qld, 1 sh, AMS C.329407; Holmes Reef, Coral Sea, Qld, 9 sh, AMS C.329430; Wreck Reef, Coral Sea, Qld, 3 sh, AMS C.88731; Cay A, Holmes Reef, Coral Sea, Qld, 2 sh, AMS C.329413; Low Isles, GBR, Qld, 14 sh, AMS C.88774, x.1928; Langford Reef, Whitsunday Group, GBR, Qld, 2 sh, AMS C.436626, xi.1969; NE Herald Cay, Coral Sea, Qld, 21 sh, AMS C.329401, 16.vi.1997; Tregosse Reefs, Diamond Islets, West Cay , Coral Sea, Qld, 6 sh, AMS C.329418, x.1964; Erscott’s Hole Lagoon (31°32.79′S- 159°03.68E), Lord Howe I., 1 spc, AMS GoogleMaps C.458246, H = 24 mm, ii.1979, sand, 3 m. New Guinea: Port Moresby , 1 sh, AMS C.88196. Solomon Islands: Honiara, Guadalcanal, 2 sh, BMNH 20060661 , H = 20.2 mm. Fiji: 1 sh, RMNH ; Nadi Bay , Viti Levu, 8 sh, AMS C.68305. New Caledonia: Lifou, RMNH ; Noumea, 1 sh, AMS C.83136. New Hebrides: 3 sh, AMS C.88186.
Shell: ( Figs 26O–R, 28D View Figure 28 ) Maximum H = 35.1 mm, thick (shells from western Pacific are thinner), shape variable from quadrangular to more elongate, wider anteriorly. Growth lines all over shell. Parietal callus thin. Spire aperture wide; spire seven whorls, with raised striae. Protoconch diameter 130–190 Mm. Background colour reddish brown mottled with pale yellow dots; pattern of three or four faint darker spiral bands present in (East African shells, or brown dots shadowed in yellow scattered over shell in western Pacific shells. Aperture pale; lip reddish; columella white.
Animal: ( Fig. 25E View Figure 25 ) Skin covered with fine purple spots. Bright white dots absent.
Jaws: ( Fig. 29D View Figure 29 ) Semicircular shape. Triangular spatulate scales with smooth rounded edges. Height 1.4 mm, width 0.6 mm (H = 22.0– 27.8 mm).
Radula: ( Figs 30E View Figure 30 , 31E View Figure 31 , 32D View Figure 32 ) In total, 26–30 rows (H = 22.0, 27.8 mm), first five rows with rachidian teeth only. Rachidian tooth 403–561 Mm wide (H = 22.0, 27.8 mm); 11 cusps; cusps triangular, sharp, decreasing in size outwards, central and marginal cusps smaller, central sometimes absent. Inner lateral tooth with large central cusp longer and twice as wide, symmetrically flanked by three cusps of decreasing size; cusps sharp. Outer lateral tooth with 4–5 cusps; innermost largest; others of similar size; base with sharp denticle.
Gizzard plates and spines: ( Fig. 33G–H View Figure 33 ) Plates 3.1– 4.4 mm long by 1.2–1.6 mm wide (H = 22.0– 27.8 mm); light orange with broad brown margin; quadrangular, left edge slightly concave, right concave; top angled; surface smooth; axial keel on left side, keel extremities prominent, usually grooved. Anterior gizzard spines twice the size of posterior spines, sharp. Posterior spines small, triangular, sharp.
Male genital system: ( Fig. 36A–C View Figure 36 ) Length 10–13 mm (H = 22.0, 27.8 mm). Vestibular area about onequarter to one-third overall length of system (excluding blind caecum), narrower than penial sheath. Prostate globular to oval. Blind caecum large (see Remarks), narrow.
Hermaphroditic gonoduct: ( Fig. 37D–E View Figure 37 ) Length 24– 29 mm (H = 22.0, 27.8 mm). First part of membrane gland internal; second part external and smooth. Seminal receptacle cylindrical, large. Gametolytic duct longer than common genital duct. Gametolytic gland twice the size of fertilization chamber. Fertilization chamber with complex network of ducts. Vestibular area small.
Ecology: This species appears to be uncommon and active at night. Inhabits shallow depths on dead coral and sand ( Ono, 1999) and has been found among seagrass. An image of a live specimen can be found in Hori (2000).
Geographical distribution: ( Fig. 39 View Figure 39 ) Occurs widely throughout the Indo-West Pacific. In the Indian Ocean it occurs in South Africa, Tanzania, Madagascar, Seychelles, Mauritius, Maldives and Andaman Islands. In the western Pacific it is known from Vietnam, Ryukyu Islands in southern Japan; southwards to New Guinea, north-eastern Australia, New Caledonia, Solomon Islands and Fiji.
Remarks: Two shell morphotypes can be recognized in this species. Indian Ocean shells are thicker and squarish with a red-brown pattern and darker faint spiral bands, whereas western Pacific shells are thin and elongate with a distinct dotted pattern.
The only anatomical description of this species was provided by Burn (1966) based on a specimen from Queensland, Australia, of small size (H = 16.5 mm), which could account for the small number of tooth rows (20) in the radula.
The male genital system is different from that of other Indo-West Pacific species in its rounded prostate and long blind caecum. Although the blind caecum is smaller and narrower than that of Atlantic and eastern Pacific species, its length is at least half the diameter of the prostate, making it proportionally larger as compared with other Indo-West Pacific species, in which the length of this structure varies from one-quarter ( B. quoyii ) to one-tenth of the prostate diameter in all other species. This was also observed by Burn (1966) in a comparasion between this species, B. vernicosa (as B. ovulum ) and B. occidentalis (as B. striata , based on Marcus, 1957). The penial duct in this species is straight and uncoiled. A synopsis of the morphological variability of shell and anatomical characters, and a comprasion with other species, is given in Table 2.
This species is uncommon and only two complete specimens were obtained for dissection and molecular phylogenetic analysis. More specimens are necessary to understand fully the morphological and anatomical variability of this species.
BULLA PEASIANA PILSBRY, 1895 View in CoL
( FIGS 25G View Figure 25 , 27A–C, 28E View Figure 28 , 38 View Figure 38 )
Bulla marmorea Pease, 1860: 431 View in CoL (Sandwich Islands [Hawaii]; holotype seen, BMNH 1961209, H = 20.0 mm; figured in Kay, 1965: 44, pl. 1, figs 13, 14; Fig. 27A herein; paratypes MCZ 297878, not seen; not Schröter, 1804). Sowerby, 1868: pl. 6, sp. 16. Mabille, 1896: 116.
Bulla peasiana Pilsbry, 1895: 348 View in CoL , pl. 34, fig. 8 (replacement name for B. marmorea View in CoL ). Kay, 1965: 44, pl. 1, figs 13, 14.
Bulla vernicosa View in CoL – Kay, 1979: 423, figs 134E, H, 137E (not Gould, 1859).
Taxonomic history: This species was originally described by Pease (1860) as B. marmorea View in CoL , based on shells from Hawaii. Finding that the name was preoccupied, Pilsbry (1895) renamed it after Pease. Occasionally the name has been used to refer to B. vernicosa View in CoL in the Kermadec Islands ( Oliver, 1915) and B. orientalis View in CoL from southern Japan ( Kuroda & Habe, 1952).
Diagnosis: Shell: small, thin, elongate, with conspicuous fine growth lines; pale reddish brown. Information on the anatomy is not available. Range: Hawaiian Islands.
Material examined: Hawaiian Islands: 1 sh, BMNH 1961209, H = 20.0 mm; Hilo, 1 sh, AMS C.34515; Hilo, 2 sh, BMNH 20060584, H = 25.7, 26.5 mm; Hilo, 3 sh, BMNH 1910.9.28.118–120, H = 20.4–23.5 mm; 1 sh (photograph), Bishop Museum 241104, H = 29 mm (data provided by C. Pittman); Maui, airport beach, 1 spc. (photograph), H = 13 mm, 2 iv.2003, 6– 14 m on sand (data provided by C. Pittman); Maliko Bay, 1 spc. (photograph), H = 15 mm, 5.vi.1999, under rock (data provided by C. Pittman); Midway Atoll, 1 sh (photograph), Bishop Museum 606306, H = 26 mm (data provided by C. Pittman).
Shell: ( Figs 27A–C, 28E View Figure 28 ) Maximum H = 29.0 mm, thin, elongate. Conspicuous and usually fine growth lines all over shell. Parietal callus thin. Spire aperture wide; spire six whorls, with raised striae. Protoconch diameter 160 Mm. Background colour light reddish, mottled with white dots and dark brown blotches; these blotches occasionally merge to produce one or two faint brown spiral bands posteriorly behind midpoint. Aperture with shell pattern visible; lip pinkish brown; columella white.
Animal: ( Fig. 25G View Figure 25 ) Varies from orange to dull brown ( Kay, 1965), with bright white dots scattered over cephalic shield, cephalic and parapodial lobes.
Jaws: Unknown.
Radula: Unknown.
Gizzard plates and spines: Unknown.
Male genital system: Unknown.
Hermaphroditic gonoduct: Unknown.
Ecology: It occurs in sheltered areas in beds of the algae Halimeda incrassata and in rubble and sand from shallow zones down to 10 m or more (C. Pittman, pers. comm.). At Ala Moana Reef, Oahu, Hawaiian Islands, it is common during January and February when animals appear in large numbers on the dead reef platform among coral and gravel at night, particularly at low tide during full moon ( Kay, 1965).
Geographical distribution: This species is endemic to the Hawaiian Islands where it is known from Midway Atoll at the north-west end of the archipelago southwards to Maui and Hawaii ( Fig. 38 View Figure 38 ).
Remarks: This species is accepted as distinct on the basis of shell characters alone; no complete specimens were obtained for either anatomical dissection or molecular analysis.
Several authors have questioned the validity of the species. Pilsbry (1895) when renaming it pointed out the similarities with B. occidentalis from the western Atlantic, raising the possibility that shells from Hawaii were ballast specimens, but although B. peasiana bears a shell of similar slender shape the outline is more straight, it lacks basal striae, and the overall length is smaller. Kay (1979) in her inventory of Hawaiian shells considered it to be a synonym of B. vernicosa . However, B. peasiana and B. vernicosa occur sympatrically in the Hawaiian Islands (photographs of living animals by C. Pittman have been checked by us). Moreover, consistent differences in shells of similar length separate these two species. Shells of B. vernicosa are thick, whereas those of B. peasiana are thin. The colour pattern in B. peasiana is paler, lacking conspicuous spiral dark bands, and the shells bear conspicuous fine axial growth lines all over the surface; in B. vernicosa there are one to three dark bands and growth lines are faint and almost undistinguishable. The shape in B. peasiana is elongate (like the Atlantic B. striata and B. occidentalis ), whereas shells of B. vernicosa are broader and quadrangular. The colour pattern is visible through the aperture, whereas the aperture in B. vernicosa is opaque white. The maximum known length of B. peasiana is about 25 mm, but B. vernicosa can reach 50 mm.
BULLA QUOYII GRAY, 1843 View in CoL
( FIGS 25C View Figure 25 , 27D–K, 28F View Figure 28 , 29E View Figure 29 , 30F View Figure 30 , 31F View Figure 31 , 32E View Figure 32 , 34A–B View Figure 34 , 36D–E View Figure 36 , 37H–I View Figure 37 , 39 View Figure 39 )
Bulla nebulosa Schröter, 1804: 20 View in CoL (types untraceable; nomen dubium, see Pilsbry, 1895).
Bulla australis Férussac, 1822: 575 View in CoL (Port Jackson [Sydney, Australia], type seen, 1 syntype, MNHN, H = 40.1 mm; figured by Valdés & Héros, 1998: 708, fig. 3H; nomen dubium).
Bulla australis Gray, 1825: 408 View in CoL ( Australia; types seen, 3 syntypes (1 broken), BMNH 20060585, H = 45.9, 53.3 mm; not Férussac, 1822). Gray, 1826: 490. Gray, 1843: 243. Brazier, 1885: 89. Whitelegge, 1889: 275. Pilsbry, 1895: 346, pl. 36, figs 17, 18. Pritchard & Gatliff, 1902: 214. Torr, 1914: 366, pl. 20, fig. 12. Iredale, 1929: 349, pl. 33, fig. 4. (All not Férussac, 1822; not Quoy & Gaimard, 1833.)
Bullaria australis – Hedley, 1916: 72 (not Férussac, 1822; not Quoy & Gaimard, 1833).
Bulla australis Quoy & Gaimard, 1833: 357 View in CoL , pl. 26, figs 38, 39 (Port du Roi Georges [King George Sound, Albany, Australia]; types seen, 11 syntypes, MNHN, H = 35.6–57.2 mm; syntype figured by Valdés & Héros, 1998: 708, fig. 3I). Deshayes & Milne-Edwards, 1836: 673. Menke, 1854b: 44. Brazier, 1878: 83. Watson, 1886: 638. Mabille, 1896: 113. Shirley, 1911: 102. Suter, 1913: 534, pl. 49, fig. 7. (All not Férussac, 1822; not Gray, 1825.)
Bullaria australis – Suter, 1913: 534, pl. 49, fig. 7 (not Férussac, 1822; not Gray, 1825).
Bulla quoyii Gray, 1843: 243 View in CoL ( New Zealand; types seen, 10 syntypes, BMNH 1842.11.18.135–142, H = 9.0–22.5; lectotype here selected, H = 22.5 mm; Fig. 27G herein). Menke, 1854b: 44. Pilsbry, 1895: 348, pl. 39, fig. 71. Willan, 1978: 58, figs 15–24. Powell, 1979: 275, pl. 54, fig. 2 (as B. quoyi ). Burn, 1984: 68. Wells & Bryce, 1988: 136, pl. 48, fig. 533. Burn, 1989: 737, fig. 12.7, b. Burn & Thompson, 1998: 957, fig. 16.36 A, B. Jansen, 2000: 70, fig. 274 (as B. quoyi ). Burn, 2006: 7.
Bullaria australis quoyi – Suter, 1913: 535.
Bullaria quoyi – Hedley, 1916: 72.
Quibulla quoyii View in CoL – Rudman, 1971.
Bulla oblonga Adams, 1850: 577 View in CoL , pl. 123, fig. 74 ( Philippines and Island of Annaa; types seen, 3 syntypes, BMNH 197648, H = 53.6–57.5 mm; one figured herein, Fig. 27D). Menke, 1854b: 44. Angas, 1867: 226. Sowerby, 1868: pl. 3, fig. 4a–c. Pilsbry, 1895: 346, pl. 35, figs 12–14.
Bulla australis var. oblonga View in CoL – Pilsbry, 1895: 346, pl. 35, figs 12–14.
Haminea castanea Adams, 1850: 584 , pl. 124, fig. 106a ( New Zealand; probable type seen, BMNH 1881.5 .20.22, H = 25.4 mm; Fig. 27F herein).
B. substriata Menke, 1853: 136 View in CoL ( Australia; types untraceable).
B. tenuissima Sowerby, 1868 View in CoL : pl. 2, fig. 4a, b (Swan River, Australia; type seen, BMNH 197650, H = 20.4 mm; Fig. 27E herein). Pilsbry, 1895: 347, pl. 39, fig. 72. Wells, 1985: pl. 1, figs 1–7.
Bullaria tenuissima – Hedley, 1916: 72.
Bullaria (Quibulla) tenuissima – Cotton & Godfrey, 1938: 32.
Quibulla tenuissima – Cotton, 1959: 404.
Bulla dubiosa Mabille, 1896: 113 View in CoL (no locality; types untraceable).
Bulla striata View in CoL – Quoy & Gaimard, 1833: 354, pl. 26, figs 8, 9 (not Bruguière, 1792). Pilsbry, 1895: 348 (not Bruguière, 1792).
Bullaria botanica Hedley, 1918 : M104 (replacement name for B. australis Gray, 1825 View in CoL ). May, 1921: 103. Macpherson & Gabriel, 1962: 242, fig. 281.
Quibulla botanica Iredale, 1929: 349 View in CoL , pl. 38, fig. 4 ( Sydney harbour, Australia, type seen, 1 neotype, AMS C.57856, H = 55.0 mm; Fig. 27H herein).
Taxonomic history: Schröter (1804) introduced at least 14 specific names in the genus Bulla View in CoL . According to Pilsbry (1895) these are indeed members of Bulla View in CoL in the modern sense but, because they were published without localities and types cannot be found, they are not identifiable with certainty and we regard them all as nomina dubia. Nevertheless, Pilsbry suggested that B. nebulosa View in CoL might be the present species.
The controversy about the valid name of the present species (e.g. Brazier, 1885; Willan, 1978) stems mainly from the fact that three different authors independently introduced the name B. australis to describe the same species. The name was first used by Férussac (1822) for specimens said to have been collected in Sydney. The original description was brief and, if the author was referring to a specimen of Bulla at all, it seems to have been a juvenile. The single type specimen in MNHN does not match the features of any Indo-Pacific species, but instead is an adult specimen of the eastern Atlantic species B. mabillei . The mismatch between the original description, type locality and type material suggests that the name must be considered a nomen dubium. Gray (1825) briefly described, but did not figure, specimens from Australia under the name B. australis , and the three syntypes in BMNH unequivocally belong to the present species. Nevertheless, as a junior homonym this name is invalid. Although Gray (1825) gave the type locality as Australia, the type lot bears a label mentioning New Zealand; this is in a different handwriting and seems to have been added later. Quoy & Gaimard (1833) used two names for this species: B. australis was introduced for Australian specimens and B. striata used for material collected in New Zealand that the authors considered to be similar to specimens from the Mediterranean Sea. Gray (1843) introduced the name B. quoyii for material from New Zealand, presumably because he recognized the use of B. striata by Quoy & Gaimard (1833) to be a misidentification. For the past century B. quoyii has been the name most commonly used to refer to this taxon (e.g. Willan, 1978; Burn & Thompson, 1998).
Adams (1850) introduced the name B. oblonga based on specimens from the Cuming collection (NHM) with type locality given as the Philippines. The specimens clearly belong to B. quoyii , restricted to Australasia, so the type locality must be incorrect. In addition, A. Adams erroneously considered juvenile shells of B. quoyii from New Zealand to belong to the genus Haminoea (as Haminea ) and described them as H. castanea .
Menke’s (1853) description of B. substriata was brief and no types have been traced; the name is attributed to B. quoyii based on the type locality ( Australia) and presence of spiral striae on the shell.
Sowerby (1868) noted a ‘remarkable’ thinness and transparent texture in specimens collected in Western Australia, and named them B. tenuissima , but inspection of the type specimen in the NHM revealed it to be a juvenile of B. quoyii .
Mabille (1896) gave a poor description of a new species that he named B. dubiosa , and gave neither illustration nor geographical data. The types are untraceable, but his description of the shape (‘ ovale allongée ’) and colour together with the presence of spiral striae at the base of the shell seem to indicate B. quoyii .
The name B. botanica was introduced by Hedley (1918) in a checklist as a replacement name for B. australis Gray, 1825 , but without any further comment. Iredale (1929) designated it as the type species of the genus Quibulla and gave Sydney Harbour as type locality.
Diagnosis: Shell: small or large, thin, slender, with anterior fine spiral grooves, marbled in yellowish brown. Jaws: semicircular. Radula : central and marginal rachidian cusps smaller in size; outer lateral similar in height to inner; base with sharp denticle. Gizzard plates: with brown marginal band. Male genital system: vestibular area one-quarter overall length; blind caecum short and narrow. Hermaphroditic gonoduct: vestibular area short; first part of membrane gland internal; seminal receptacle inconspicuous, inserting laterally; fertilization chamber with complex structure. Range : south-west to southeast Australia, northern Tasmania and northern North Island , New Zealand; Kermadec Islands . COI sequences: GenBank DQ986557 View Materials –986558 ( BMNH20030344 ; BMNH20030292 ) .
Material examined: Australia: Newcastle (32°56′S, 151°46′E), NSW, 1 sh, AMS C.88795; Jervis Bay, NSW, 8 sh, BMNH 20060586, H ~ 38.1 mm; Hare Bay, Jervis Bay (35°0′S, 150°46′E), NSW, 1 sh, AMS C.95856; Sydney, NSW, 2 sh, BMNH 20060587, H = 36.4 mm; Sydney, NSW, 4 sh, BMNH 20060588, H ~ 34.2 mm; Sydney, NSW, 6 sh, BMNH 1891.11.6.423–8, H ~ 21.6 mm; Jiguma Beach, NSW, 1 sh, BMNH 20060589, H = 44.7 mm; Ulladulla (35°21.5S, 150°28.5E), NSW, 1 sh, AMS C.94576; Quarantine Bay, Towfold Bay, NSW, 1 spc, AMS C.328874; Lord Howe I., Australia, 1 sh, AMS C.366497; Lord Howe I., Australia, 4 sh, AMS C.366498; North end of Chinaman Long Beach, Wilson’s Promontory (38°50′S, 146°25′E), Vic., 26 sh, AMS C.163725; Melbourne, Vic., 1 sh, BMNH 20060590, H = 29.4 mm; Elizabeth Reef, Tasman Sea, Vic., 1 sh, AMS C.88788; Port Welshpool, Vic., 1 sh, BMNH 20060593, H = 32.0 mm; Wilson’s Promontory, Vic., 2 sh, BMNH 20060594, H = 43.6 mm; Adelaide, SA, 5 sh, BMNH 20060591, H ~ 37.3 mm; Point Turton, York Peninsula, Adelaide, SA, 4 sh, BMNH 20060592, H = 62.2 mm; Port Lincoln (34°47′S, 135°51′E), SA, 2 sh, AMS C.90592; Kingscote area, Kangaroo I. (35°39.5S, 137°39′E), SA, 4 sh, AMS C.91413; Streaky Bay (32°48′S, 134°12′E), SA, 12 sh, AMS C.436373; Tasmania, 1 sh, BMNH 20060599, H = 21.0 mm; near Devonport, Port Sorell (41°8′S, 146°34′E), 1 sh, AMS C. 17667; Whitemark, Flinders I., Bass Strait, 7 sh, AMS C.170210; Emu Point, Oyster Harbour, Albany, WA, 3 dis, WAM 19095, H = 42.7–45.9 mm, ii.1983, 4 m depth; Bandy Creek Harbour, Esperance, WA, 1 dis, BMNH 20030050, H = 46.5 mm, ii.2003, 5 m depth, among green algae; Collesloe Beach, WA, 1 sh, BMNH 20060595, H ~ 21.9 mm; Perth, WA, 5 sh, BMNH 20060596, H ~ 44.7 mm; Perth, WA, 2 sh, BMNH 20060597, H = 62.5 mm; Trigg I. (north of Perth), WA, 17 sh, BMNH 20060598, H ~ 21.9 mm; Perth, WA, 1 sh, BMNH 197650, H = 10.6 mm; Banbury (33°37′S, 115°28′E), WA, 2 sh, AMS C. 436336; Leeuwin (34°21′S, 115°10′E), WA, 2 sh, AMS C.436329; Pelsart I., Abrolhos Islands, WA, 1 sh, AMS 69320; Peron Peninsula, Shark Bay (25°37′S, 113°22′E), WA, 1 sh, AMS C.436360, H = 44 mm, 5.viii.1943. New Zealand: Tamaki Estuary, Auckland, 2 dis, BMNH 20030345, H = 27.1, 29.6 mm, iii.2003, mud with algae at low tide; Cheltenham, North I., 5 sh, AMS 171655; Auckland, 1 sh, BMNH 20060600, H = 26.0 mm; Auckland Harbour, 6 sh, BMNH 20060601, H ~ 28.6; Long Beach, Bay of Islands, 1 sh, BMNH 20060602, H = 21.3 mm; Tokeran Bay, Northland, 2 sh, AMS C.90491; Torbay Beach, Hauraki Gulf, 2 sh, AMS C.96248; Sunday I., Kermadec Islands, 3 sh, AMS C.68712.
Shell: ( Figs 27D–K, 28F View Figure 28 ) Maximum H = 62.2 mm, thin, shape variable from elongate- to ovatequadrangular, wider anteriorly. Both sides convex; 5–9 anterior spiral grooves, usually faint. Faint growth lines all over shell. Parietal callus thin. Spire aperture narrow; spire 6–7 whorls, with raised striae. Protoconch diameter 190 Mm. Background colour pale brown, mottled with dark brown blotches shadowed with yellowish white that merge to form axial zigzag bands. Aperture white with shell pattern visible in some areas; lip pale brown; columella white. Periostracum greenish to orange.
Animal: ( Fig. 25C View Figure 25 ) Purple or pale brown, with patches of bright white dots scattered over cephalic shield, cephalic and parapodial lobes. Area between the eyes and rear part of cephalic shield darker; anterior edge of cephalic shield darker. Foot pale brown with white dots scattered over surface.
Jaws: ( Fig. 29E View Figure 29 ) Semicircular shape. Triangular spatulate scales with smooth rounded edges. Height 1.7–2.3 mm, width 0.5–0.9 mm (H = 27.1–45.9 mm).
Radula: ( Figs 30F View Figure 30 , 31F View Figure 31 , 32E View Figure 32 ) In total, 26–29 rows (H = 27.1–45.9 mm), first 4–6 rows with rachidian teeth only. Rachidian tooth 504–762 Mm wide (H = 27.1, 45.9 mm); 10–12 cusps; cusps triangular, sharp, last marginal cusp of smaller size, central cusp usually present, smallest. Inner lateral tooth with large central cusp twice as long and wide, symmetrically flanked by three cusps of similar size; cusps sharp. Outer lateral tooth with six cusps; innermost largest; others of similar size, outermost smaller; base with sharp denticle.
Gizzard plates and spines: ( Fig. 34A–B View Figure 34 ) Plates 4.6– 6.3 mm long by 1.4–2.6 mm wide (H = 27.1– 45.9 mm); yellow with brown margin; quadrangular, left edge slightly concave or straight, right concave; top angled; surface smooth; axial keel on left side, keel extremities prominent, usually grooved, right lateral expansions developed. Anterior gizzard spines triangular, sharp. Posterior spines small, conical, sharp.
Male genital system: ( Fig. 36D–E View Figure 36 ) Length 24–37 mm (H = 45.9, 29.6 mm). Vestibular area about onequarter to one-third overall length of system (excluding blind caecum), nearly as wide as penial sheath. Prostate cylindrical like penial sheath or oval. Blind caecum small and narrow.
Hermaphroditic gonoduct: ( Fig. 37H–I View Figure 37 ) Length 35– 43 mm (H = 29.6, 46.5 mm). First part of membrane gland internal; second part external and smooth. Seminal receptacle inconspicous. Gametolytic duct longer than common genital duct, inserting laterally. Gametolytic gland four times the size of fertilization chamber. Fertilization chamber with complex network of ducts. Vestibular area small.
Ecology: Common on intertidal mudflats in sheltered areas, on sandy mud with seagrass Zostera and green algae Ulva and Enteromorpha ( Powell, 1979) and also in shallow subtidal areas. Breeds during November when it can be seen abundantly at night ( Wells & Bryce, 1993; Chuk, 2003; Furneaux, 2003). A photograph of a live specimen is given by Wells & Bryce (1993).
Geographical distribution: ( Fig. 39 View Figure 39 ) This is a temperate species restricted to Australasia, occurring from the Abrolhos Islands on the west coast of Australia, all along the southern coast; in northern Tasmania and on the east coast as far north as Sydney and Newcastle. In New Zealand it is known from the northern parts of North Island. It is present in Lord Howe I and Kermadec Islands.
Remarks: Shells of this species are thin and light and adults can reach lengths over 60 mm, which makes this species the largest in the family. Specimens between 15 and 30 mm are already mature ( Willan, 1977; present data). New Zealand specimens are smaller (max. H = 45.0 mm; Willan, 1977) with squarish shells. The variability of shapes found in this species is ecophenotypic, according to Willan (1977); shells from sheltered localities tend to be large and elongate, whereas shells from exposed shores are smaller and more globose.
Among the Indo-West Pacific species B. quoyii is the only one with spiral striae on the base of the shell, a feature otherwise only present in the Atlantic B. striata and B. occidentalis . These striae can be faint or inconspicous and although their number can vary between nine and 24 ( Willan, 1977), it is usually lower (5–9).
Also unique among the family are the lateral insertion of the gametolytic duct in the common genital duct and the inconspicuous seminal receptacle.
Data on the morphology of the radula and gizzard plates provided by Torr (1914), Willan (1977) and Rudman (1971) agree with the present observations, but the width of the rachidian tooth depicted by Willan (1977) is 3–4 times smaller (164 Mm). Willan (1977) did not provide the length of the dissected specimen, mentioning only that it was an adult. This striking difference is probably the result of an error in the scale bar.
The penial duct can be either coiled lengthwise or straight. The hermaphroditic gonoduct of Australian specimens was found to be about half the size of shell, whereas this structure is at least as long and usually longer than the shell in specimens from New Zealand. A synopsis of the morphological variability of shell and anatomical characters, and a comparison with other species, are given in Table 2.
This is the only temperate Bulla species in the Indian and Pacific oceans. Material from Shark Bay in Western Australia was located in the collections of one museum (AMS), but these were old and worn shells, which had been collected washed ashore. There is no evidence for the present occurrence of this species north of the Abrolhos Islands, and even here it is rare and only confirmed by dead shells.
BULLA VERNICOSA GOULD, 1859 View in CoL
( FIGS 25D, F View Figure 25 , 27L–Q, 28G–H View Figure 28 , 29F View Figure 29 , 30G–H View Figure 30 , 31G–H View Figure 31 , 32F View Figure 32 , 34C–D View Figure 34 , 36F–H View Figure 36 , 37J–K View Figure 37 , 39 View Figure 39 )
Bulla marmorea Schröter, 1804: 20 View in CoL (types untraceable; nomen dubium, see Pilsbry, 1895).
Bulla australis View in CoL – Adams, 1850: 576, pl. 122, figs 64– 66 (not Gray, 1825 = B. quoyii View in CoL ; not Quoy & Gaimard, 1833). Sowerby, 1868: pl. 4, sp. 12a–c (not Gray, 1825; not Quoy & Gaimard, 1833).
Bulla adamsi – Menke, 1854b: 43. Pilsbry, 1895: 345, pl. 35, figs 15–16, 19–20. Melvill & Standen, 1899: 155. Hedley, 1910: 370. Suter, 1913: 534, pl. 49, fig. 6. Oostingh, 1923: 139. Guangyu, 1997: 94, pl. 26, fig. 3. (All not Menke, 1850; see B. punctulata View in CoL .)
Bullaria adamsi – Suter, 1913: 534, pl. 49, fig. 6 (not Menke, 1850).
Bulla vernicosa Gould, 1859: 138 View in CoL (Loo Choo Islands [Ryukyu Islands], Japan; types untraceable; neotype here designated, Ryukyu Islands, Japan, BMNH 20060603, H = 39.0 mm; Fig. 27P herein). Pilsbry, 1895: 349. Habe, 1950: 21, pl. 3, fig. 13. Johnson, 1964: 166. Kuroda, Hhabe & Oyama, 1971: 288, pl. 64, fig. 26. Cernohorsky, 1972: 207, pl. 59, fig. 4. Powell, 1976: 110. Willan, 1978: 63, figs 3–14. Powell, 1979: 275, pl. 54, fig. 1. Lin & Qi, 1985: 114. Springsteen & Leobrera, 1986: 284, pl. 81, fig. 11. Short & Potter, 1987: 116, fig. 5. Dharma, 1992: 76, pl. 16, fig. 21. Fukuda, 1994: 40. Guangyu, 1997: 92, pl. 8, fig. 1. Burn & Thompson, 1998: 958. Higo et al., 1999: G4896. Dharma, 2005: 206, pl. 78, fig. 14. Thach, 2005: 230, pl. 71, fig. 6. Tröndlé & Cosel, 2005: 320. Cobb & Willan, 2006: 32, figured.
Bulla ventricosa – Hori, 2000: 758, pl. 377, fig. 1 (mispelling).
Bulla magdelus – Angas, 1867: 227 (error for B. amygdalus View in CoL ; not Dillwyn, 1817 = B. occidentalis ).
Bulla ovulum Gould View in CoL in Angas, 1867: 227 (published in synonymy, unavailable). Burn, 1966: 96, figs 1–4.
Bulla ovula Gould View in CoL in Sowerby, 1868: pl. 2, sp. 5a, b (no locality; types seen, 3 syntypes, BMNH 197645, H = 26.0, 27.4, 31.3 mm; largest figured in Higo et al., 2001: 141, Fig. 27L herein). Mabille, 1896: 116.
Bulla conspersa Pease, 1869: 72 View in CoL , pl. 8, fig. 9 (Marquesas Islands; lectotype seen, ANSP 5705, H = 24.1 mm; Fig. 27O herein; paralectotypes MCZ 297880, 298464, not seen). Pilsbry, 1895: 349, pl. 39, fig. 73.
Bulla angasi View in CoL – Iredale, 1929: 350, pl. 33, fig. 8. Willan, 1978: 60, figs 1–14. Short & Potter, 1987: 116, fig. 3. Jansen, 2000: 70, fig. 273. Cobb & Willan, 2006: 30, figured. (All not Pilsbry, 1895 = B. mabillei .)
Bulla secunda Mabille, 1896: 112 View in CoL ( Japan; types seen, 6 syntypes, MNHN, H = 32.3– 20.2 mm, syntype figured by Valdés & Héros, 1998: 710, Fig. 7D View Figure 7 ).
Bulla subaustralis Mabille, 1896: 114 View in CoL ( Tahiti and Philippines; types untraceable).
Bullaria peasiana – Oliver, 1915: 542 (not Pilsbry, 1895).
Quibulla scotti Iredale, 1937: 258 View in CoL , pl. 16, fig. 11 ( Elizabeth Reef , Tasman Sea, Australia; holotype seen, AMS C60245, H = 36 mm; Fig. 27N herein).
Bulla difficilis Habe, 1950: 21 View in CoL , pl. 3, fig. 13 (Great Loochoo, Okinawa Group [Okinawa, Ryukyu Islands, Japan]; type in National Science Museum, Tokyo, NSMT-Mo 54781, not seen). Guangyu, 1997: 94, pl. 7, fig. 4. Higo et al., 1999: G4896. Hori, 2000: 758, pl. 377, fig. 4. Thach, 2005: 230, pl. 71, fig. 13.
Bulla (Quibulla) subtropicalis Powell, 1965: 167 View in CoL , pl. 22, figs 8, 9 (Norfolk I., Australia; holotype in Auckland Museum, TM-1245, not seen).
Bulla ampulla View in CoL – Fukuda, 1995: 31, pl. 74, 038 (not Linnaeus, 1758).
Bulla punctulata View in CoL – Thach, 2005: 230, pl. 71, fig. 8 (not A. Adams, 1850).
Taxonomic history: The Bulla species described by Schröter (1804) are considered nomina dubia (see Pilsbry, 1895; and Taxonomic history of B. quoyii View in CoL ).
Misunderstanding of the taxonomic status of the present species has resulted mainly from the fact that the original description of Gould (1859) was brief and without an illustration. Although the type material of this species cannot be traced, the original description and type locality can only refer to the present species. Sowerby (1868) named B. ovula based on a manuscript name of Gould, from unknown locality, although the label that is now with the specimens mentions the Ryukyu Islands in Japan. Sowerby’s illustration and study of the type material showed this to be B. vernicosa . Angas (1867) considered B. ovulum to be a synonym of B. magdelus , a name that he attributed to Lister. This is presumably an error for ‘ amygdalus ’, a name introduced by Dillwyn (1817) for West Indian shells and incorrectly attributed to Lister by Adams (1850) (see Taxonomic history of B. striata group). The description and drawing of B. conspersa ( Pease, 1869) are both inconclusive, but based on the type locality (Marquesas Islands) and examination of the holotype it is clear that the author was referring to B. vernicosa . The problems with the names B. angasi and B. adamsi have been discussed above (see Taxonomic history of B. mabillei and B. punctulata ).
Mabille (1896) named the species twice, as B. secunda and B. subaustralis . The study of the type specimens of the former showed it to be B. vernicosa . The types of the latter were untraceable, but the description coupled with the geographical range indicates B. vernicosa . During the 20th century three additional names have been proposed for this species. Iredale (1937) introduced Quibulla scotti with type locality Lord Howe Islands off eastern Australia. Habe (1950) introduced B. difficilis for a juvenile (12.8 mm) specimen from Japan, which he distinguished from the ‘allied species’ B. peasiana by the thickness and colour of shells. However, the illustration and features provided by the author are no different from juveniles of B. vernicosa . Powell (1965) proposed B. subtropicalis for shells washed up on beaches in the north of North Island, New Zealand, and on Norfolk Island.
Adams (1850) used the name B. australis Quoy & Gaimard, 1833 (= B. quoyii from Australasia) for shells from Tahiti. Occasionally, authors have misidentified the species as B. ampulla or B. punctulata (e.g. Fukuda, 1995; Thach, 2005).
In view of the confusion that has surrounded the identity of Indo-Pacific Bulla species , a type specimen is highly desirable to fix the concept of B. vernicosa . We therefore designate as neotype a specimen from Japan (ICZN, 1999: art. 75).
Diagnosis: Shell: small, thick, globose to quadrangular, reddish brown. Jaws: elongate. Radula: rachidian marginal and central cusps smaller in size; outer lateral as high as inner; base with small, sharp denticle. Gizzard plates: with broad dark brown marginal band. Male genital system: separation between vestibular and penial area inconspicuous; blind caecum minute; apical area of prostate sheath translucent. Hermaphroditic gonoduct: vestibular area large; first part of membrane gland internal; seminal receptacle large and oval; fertilization chamber with complex structure. Range: West Pacific, from Malaysia to Japan, Indonesia and northern Australia; New Guinea to Tahiti; Hawaii. COI sequences: GenBank DQ986569 View Materials -986570 (BMNH20040858; BMNH20050598).
Material examined: Malaysia: Tanjong Rhu, N. Langkawi Islands , 2 sh, AMS C.2413; Semporna , Sabah, 5 sh, BMNH 20060604 , H ~ 25.5 mm. Japan: Ryukyu Islands , 4 sh, BMNH 20060603 , H = 38.7 mm; 2 sh, AMS C.38739. Hawaii: Honolulu, Oahu , 1 dis, BMNH 20030042 , H = 31.0 mm. Philippines: 1 dis, BMNH 1867.5 . 22.47, H = 26.0 mm; Bunacan I., Bohol, 2 sh, AMS C.103221; Panglao I., Bohol, 1 dis, MNHN , Paris , H = 27.7 mm; 1 sh, AMS 162989 ; Mataburg, SW of Manila , 2 sh, AMS C.94820; Batangas, Luzon, 4 sh, AMS C.108261. Indonesia: Mahakan River , Kalimantan, 1 sh, AMS C.99648; Banggai Archipelago , 1 sh, BMNH 20060605 , H = 20.4 mm; Sanur Beach, Bali, 1 sh, AMS C.106366. East Timor: Dollar Beach , E of Dili, 18 sh, BMNH 20040855 , H = 29 mm, 23.vii.2004; Dili, 1 dis, BMNH 20040857 , H = 25.0 mm. Papua New Guinea: Galelum Island , Siassi Group, Morobe Dist., 1 dis, AMS C.444875, H = 31.2 mm; Samarai , 2 sh, BMNH 20060606 , H = 28.3 mm; near Port Moresby, 2 sh, AMS C.58314; Fishermen Island , Port Moresby, 1 sh, AMS C.91580; Rabaul, New Britain, 2 sh, AMS C.74640. Solomon Islands: Tulagi Island , 6 sh, BMNH 1962666 , H ~ 28.1 mm; Honiara, Guadalcanal Island , 3 sh, BMNH 20060607 , H ~ 22.8 mm; Bamama Island , 2 sh, BMNH 20060608 , H = 21.1 mm. New Caledonia: Koumac, 3 dis, MNHN , H = 37.8–38.8 mm; Touho, 1 sh, AMS C.94571; Baie des Citrons, Nouméa , BMNH 20060609 , H = 25.8 mm; 1 sh, BMNH 20060610 , H = 29.3 mm. Vanuatu: 2 sh, BMNH 20060611 , H = 34.8 mm; Erakor Lagoon, Efate Island , 2 sh, AMS C.108801. Samoa: Pago Pago Harbor, Tutuila, 6 sh, AMS C.71693. Tonga: Oholei Beach, 30 sh, AMS C.131753. Fiji: 2 sh, BMNH 1926.12.7.11–12, H = 23.9 mm; Suva, Viti Levu , 1 dis, BMNH 20060612 , H = 20.2 mm; 1 sh, BMNH 20060613 , H = 32.9 mm; Dawasamu, Tailevu , BMNH 20060614 , H ~ 28.7 mm. Tahiti : 2 sh, BMNH 20060615 , H = 49.1 mm. Australia: Clerke Reef, Badwell Island , Rowley Shoals, WA, 1 dis, WAM S19153 View Materials , H = 31.1 mm; Exmouth Gulf, WA, 1 sh, AMS C.90836; Learmouth, Exmouth Gulf, 1 sh, AMS C.95704; Vansittart Bay , WA, 2 sh, AMS C.45264; Beagle Bay , WA, 4 sh, AMS C.88768; Broome, Roebuck Bay, WA, 5 sh, AMS C.77959; 1 sh, AMS C.107537; Lady Nora Island , NW of Dampier, 2 sh, AMS C.141114; Port Hedland , WA, 1 sh, AMS C.94554; Shark Bay Peninsula WA, 2 sh, AMS C.99883; Daly R., NT, 1 sh, AMS C.68856; Smith Point Area , Cobourg Peninsula, NT, 1 sh, AMS C.436385; Casuarina Beach, Lizard Island , Queensland, 1 dis, WAM S19142 View Materials , H = 28.4 mm; Middle Cay, NE of Cairns, Qld, 1 dis, AMS C.444880, H = 28.0 mm; Michaelmas Cay, Qld, 1 dis, AMS C.44872, H = 34.9 mm; Moreton Bay, Qld, 4 sh, 103221; Beaver Cay , Qld , 1 sh, BMNH 20060616 , H = 38.2 mm; Cato I., Coral Sea, 2 sh, AMS C.329410; Observatory Cay, Bellona Reef, Qld, 1 sh, AMS C.329428; Creal Reef Cay, Qld, 2 sh, AMS C.329402; Wreck Reef, Coral Sea, AMS C.88772; Sydney, NSW, 2 sh, BMNH 1887.2.9.2149–52, H = 20.6, 25.6 mm; north side Long Reef, Sydney, NSW, 1 sh, AMS C.458445, 25 September, 1977, in shell hubble; Lord Howe Island . 30 sh, AMS C.88777; 1 sh, AMS C.6907; 20 sh, AMS C.59556; south of Erscott’s Hole (31°33′S, 159°4′E), Lord Howe I., AMS GoogleMaps C.380453, 1 spc, H = 24 mm, 20 i.1985, dead rubble reef on outer edge of lagoon, intertidal; Erscott’s Hole Lagoon (31°32.7′S, 159°3.6′E), Lord Howe Island , AMS GoogleMaps C.458223, 4 spcs, H ~ 22 mm, ii.1979, sand; Emily Bay, Kingston , Norfolk I. (29°4′S, 167°58′E) GoogleMaps , Norfolk Island. 1 spc, AMS C.417230, H = 23 mm, 6.xii.1988, 2 m; Norfolk Island, 7 sh, AMS C.94588; Elizabeth Reef, Tasman Sea, 2 sh, AMS C.329421; Middleton Reef, 5 sh, AMS C.329422. New Zealand: 2 sh, BMNH 1850.6 . 10.53, H = 15.8, 22.2 mm; Kermadec Islands , 5 sh, AMS C.94560.
Shell: ( Figs 27L–Q, 28G–D View Figure 28 ) Maximum H = 49.1 mm, thick, shape globose to quadrangular, wider anteriorly. Parietal callus thin; collumella thick. Spire aperture wide; spire six whorls, with raised striae. Protoconch diameter 160–200 Mm. Background colour reddish brown, mottled with pale yellow dots; 2–3 dark spiral bands present anteriorly, posteriorly and behind midpoint; sometimes faint. Australian shells pale, whitish. Aperture white with shell pattern visible; lip reddish brown; columella white.
Animal: ( Fig. 25D, F View Figure 25 ) Light brown to reddish with patches of bright white dots scattered over cephalic shield, cephalic and parapodial lobes. Buccal bulb pink and visible through skin. Juveniles with red pigmentation scattered all over a whitish skin. Foot pale yellow with bright white dots scattered over surface.
Jaws: ( Fig. 29F View Figure 29 ) Elongate shape. Triangular spatulate scales with smooth rounded edges. Height 1.4– 2.0 mm, width 0.4–0.5 mm (H = 25.0–31.0 mm).
Radula: ( Figs 30G–H View Figure 30 , 31G–H View Figure 31 , 32F View Figure 32 ) In total, 26–29 rows (H = 25.0– 34.9 mm), first four rows with rachidian teeth only. Rachidian tooth 481–857 Mm wide (H = 27.7, 37.8 mm); 11–12 cusps; cusps sharp, triangular with quadrangular base, last marginal cusp of smaller size, central cusp usually present, smallest. Inner lateral tooth with large central cusp longer and twice as wide, symmetrically flanked by 3–4 cusps of decreasing size; cusps sharp. Outer lateral tooth with 5–6 cusps; innermost largest; others of similar size, occasionally bifid, outermost smaller; base with sharp small denticle.
Gizzard plates and spines: ( Fig. 34C–D View Figure 34 ) Plates 3.8– 5.8 mm long by 1.2–3.0 mm wide (H = 25.0– 45.4 mm); yellow brown with brown margin; quadrangular, left edge slightly concave or straight, right concave; top angled; surface smooth or with fine regular concentric rings; axial keel on left side, keel extremities prominent, usually grooved. Anterior gizzard spines triangular, sharp. Posterior spines small, conical, sharp, with globular base.
Male genital system: ( Fig. 36F–H View Figure 36 ) Length 8–17 mm (H = 20.2, 34.9 mm). External border between vestibular area and penial sheath usually not evident; vestibular area about one-third overall length of system (excluding blind caecum). Prostate cylindrical, inflated. Blind caecum minute; apical area of penial sheath translucent.
Hermaphroditic gonoduct: ( Fig. 37J–K View Figure 37 ) Length 27– 50 mm (H = 28.4, 37.7 mm). First part of membrane gland internal; second part external, narrow and smooth. Seminal receptacle large, cylindrical, oval. Gametolytic duct longer than common genital duct; apical area globular, inflated. Gametolytic gland twice size of fertilization chamber. Fertilization chamber with complex network of wide ducts. Vestibular area large.
Ecology: This is a mainly intertidal and shallow subtidal species that can also occur as deep as 20–30 m ( Higo et al., 1999; Warren, 2001) and lives in habitats of soft sediment with seagrass. It is active at night and in Sulawesi, Indonesia, spawns between June and August (M. A. E. Malaquias, pers. observ.).
Geographical distribution: ( Fig. 39 View Figure 39 ) This species is broadly distributed throughout the west Pacific, occurring in Malaysia, Hong Kong, Okinawa and Amami Islands and Sagami Bay in southern Japan, central and northern Philippines, throughout Indonesia (East Borneo, Java, Bali, Lombok, Flores, Timor, Sulawesi, Moluccas), northern Australia from Shark Bay in the west to the Brisbane area in the east, off-shore reefs of the Coral Sea, Norfolk Island, Kermadec Islands, Papua New Guinea, Bismarck and Solomon Islands, New Caledonia, Vanuatu, Fiji, Tahiti, Marquesas Islands and Hawaii.
Remarks: In the west Pacific this species can only be confused with juveniles of B. ampulla , although its squarish shape, wider anteriorly, and thick shell allow easy recognition. Nevertheless in B. vernicosa shells of similar size can show different levels of development. Although the maximum length of this species is 49 mm, the mean size of adults is smaller (25 mm) and specimens become mature above 20 mm. The shell shape and pattern are uniform across the distribution, with the exception of northern Australia where a pale almost whitish form occurs together with the conventional pattern.
Anatomically, this species bears unique features. For example, the blind caecum in the male genital system is minute, sometimes indiscernible, or barely projecting. This was also observed by Burn (1966) in specimens from Queensland, Australia.
The penial duct is usually slightly coiled lengthwise. The seminal receptacle in the hermaphroditic gonoduct can be smooth or wrinkled, and is usually proportionally larger than in other species (as large as the common genital aperture). Nevertheless, this seems to result from a high capacity of storage sperm with subsequent changes in the shape and dimensions of the receptacle rather than being a typical feature of the species. The fertilization chamber is made up of ducts that are wide and pressed against each other.
The radula of B. vernicosa was depicted by Habe (1950; from Japan) and Burn (1966; from Queensland, Australia) and no notable differences are apparent. A synopsis of the morphological variability of shell and anatomical characters, and a comparison with other species, is given in Table 2.
Beach shells of this species are known from the north of North Island, New Zealand ( Powell, 1965), but viable populations do not seem to have established in the region ( Burn & Thompson, 1998).
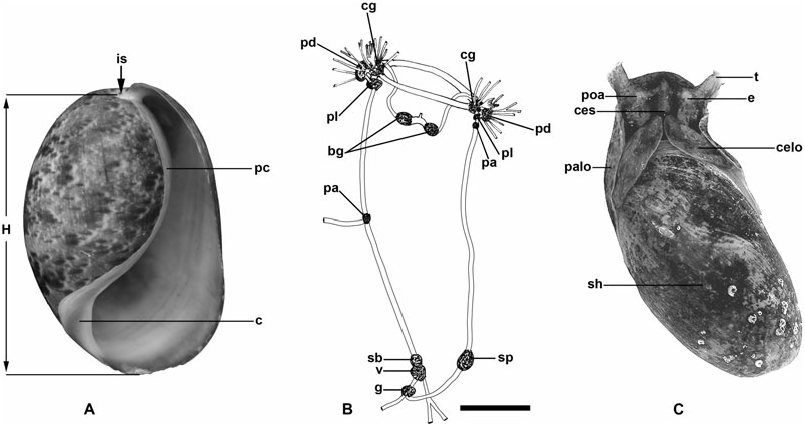






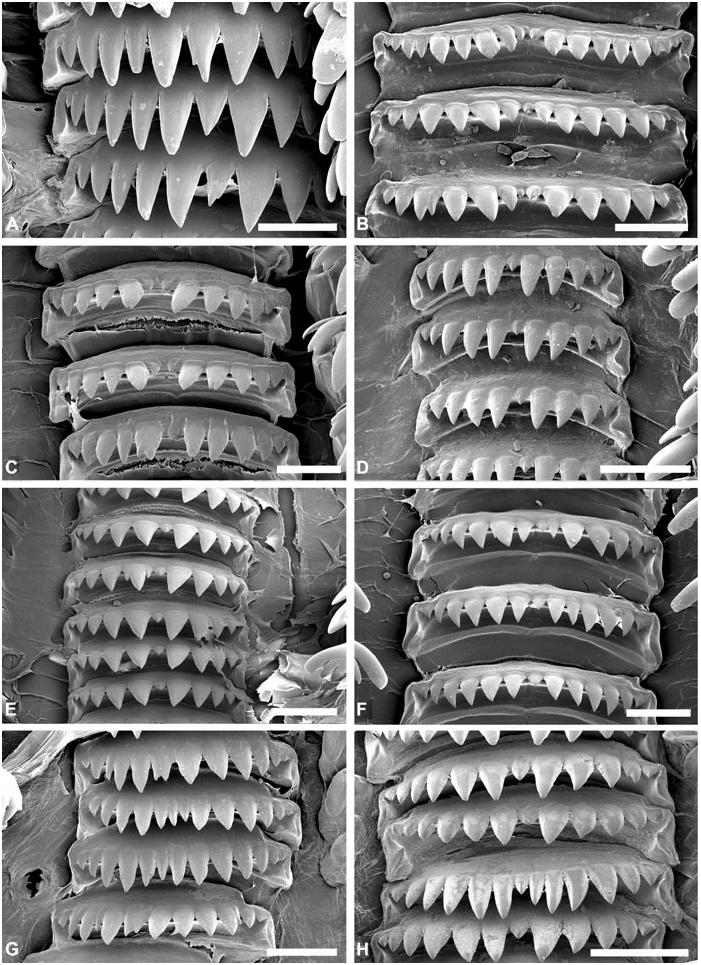
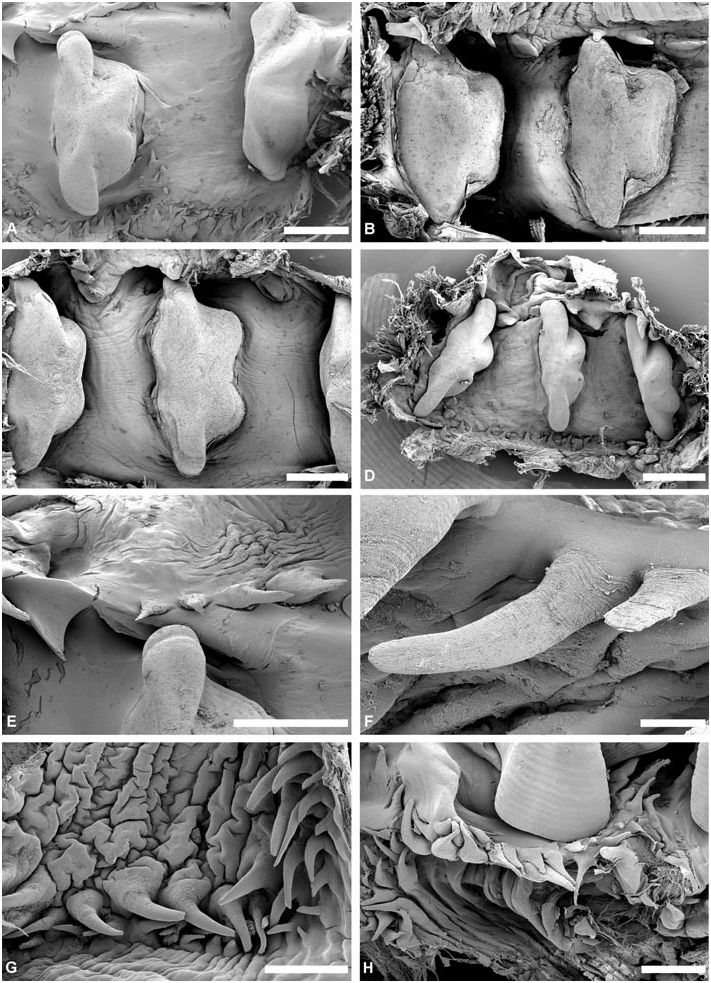
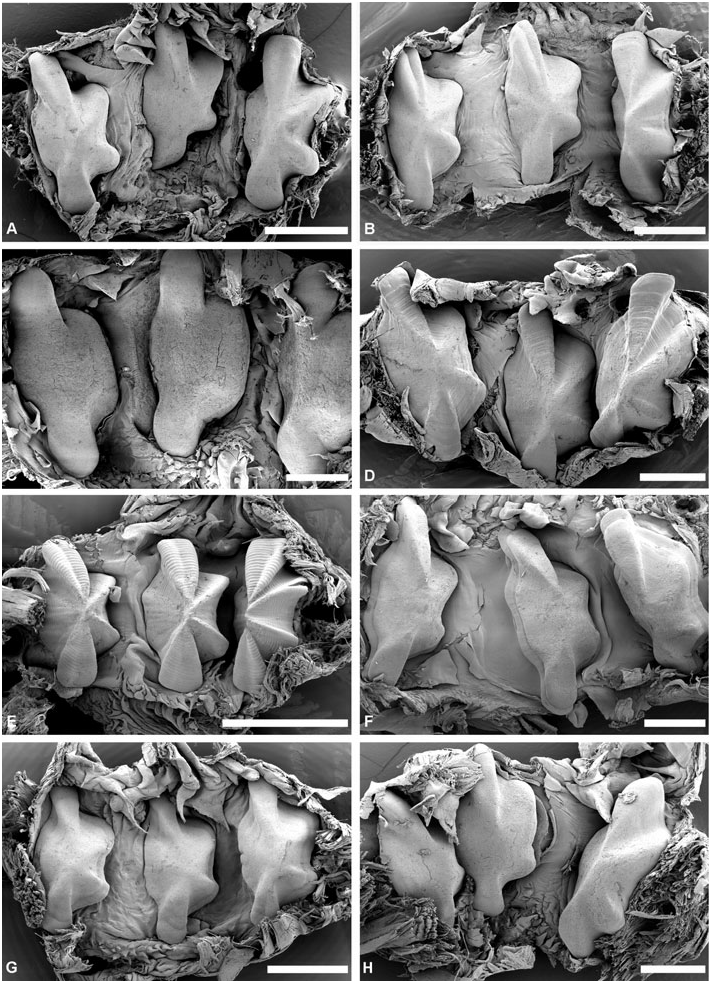



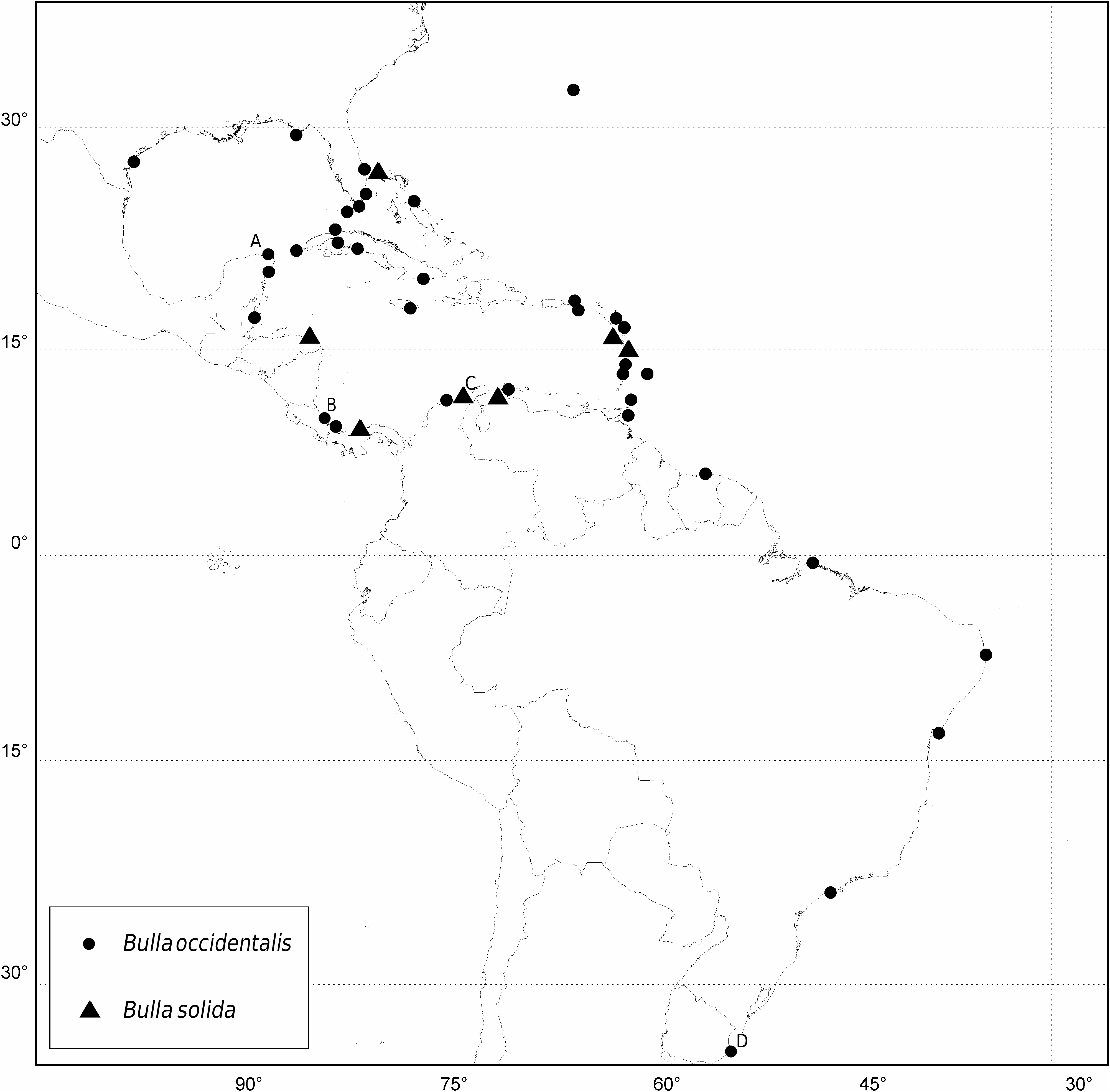


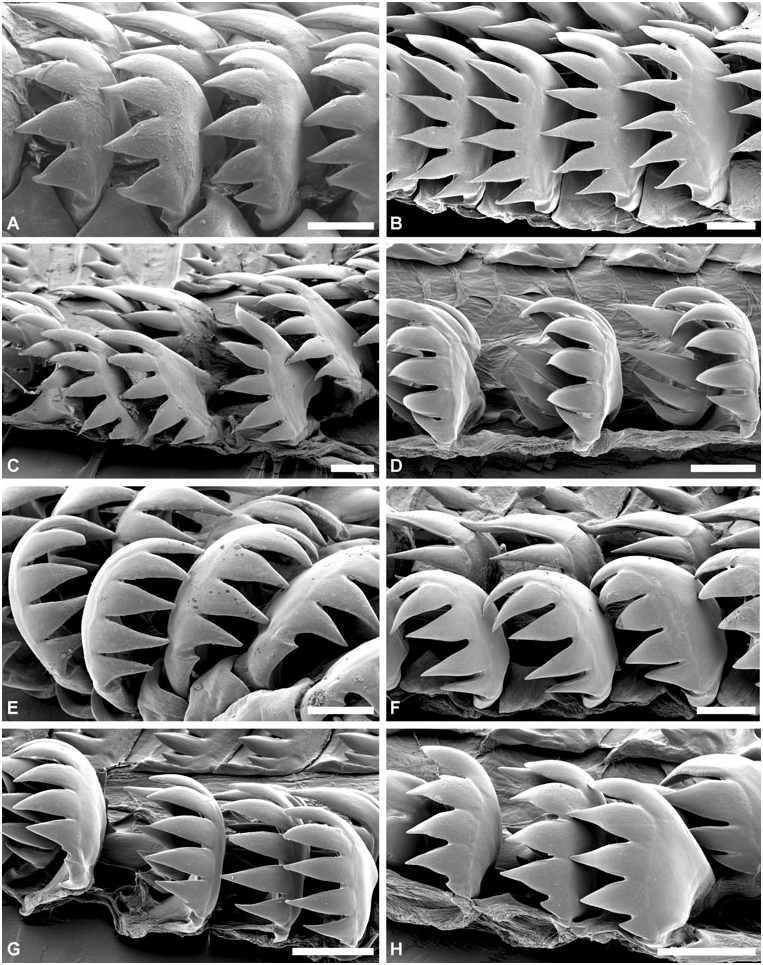
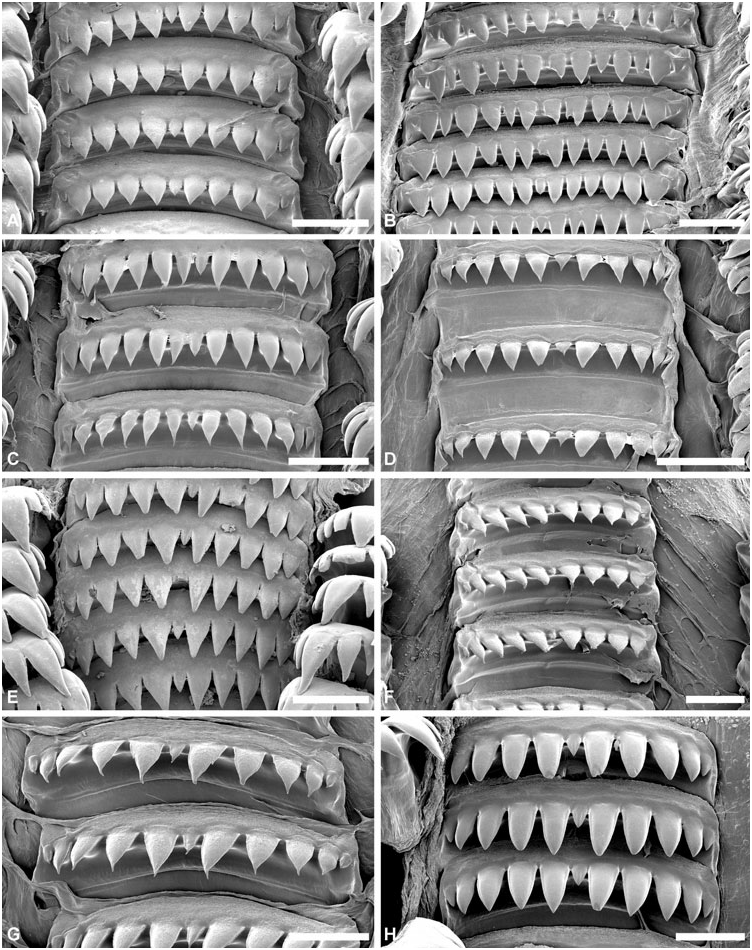
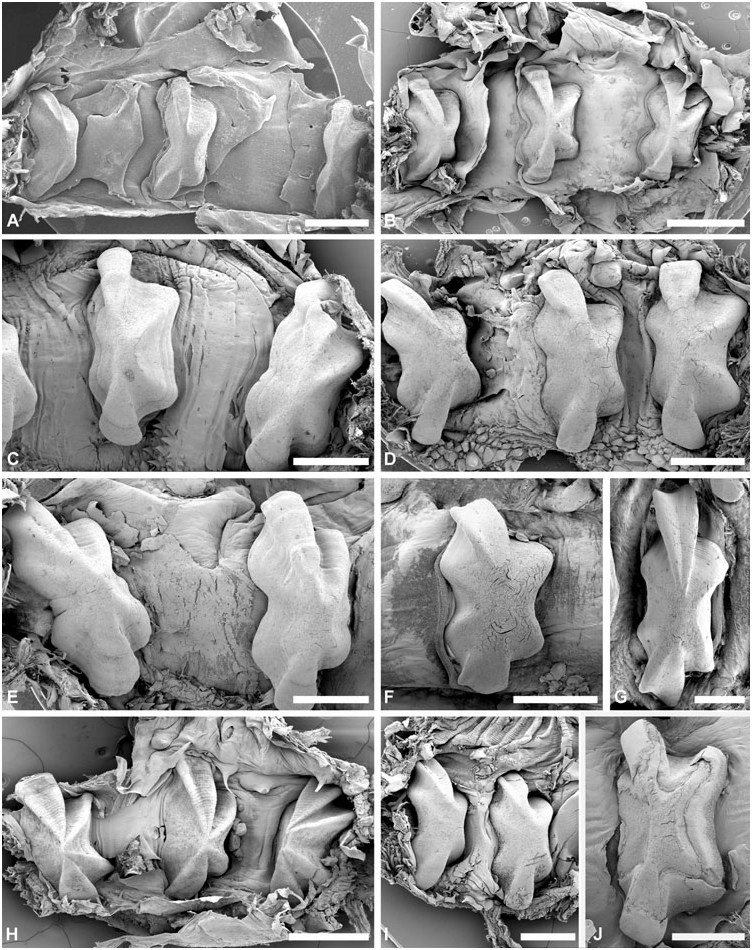

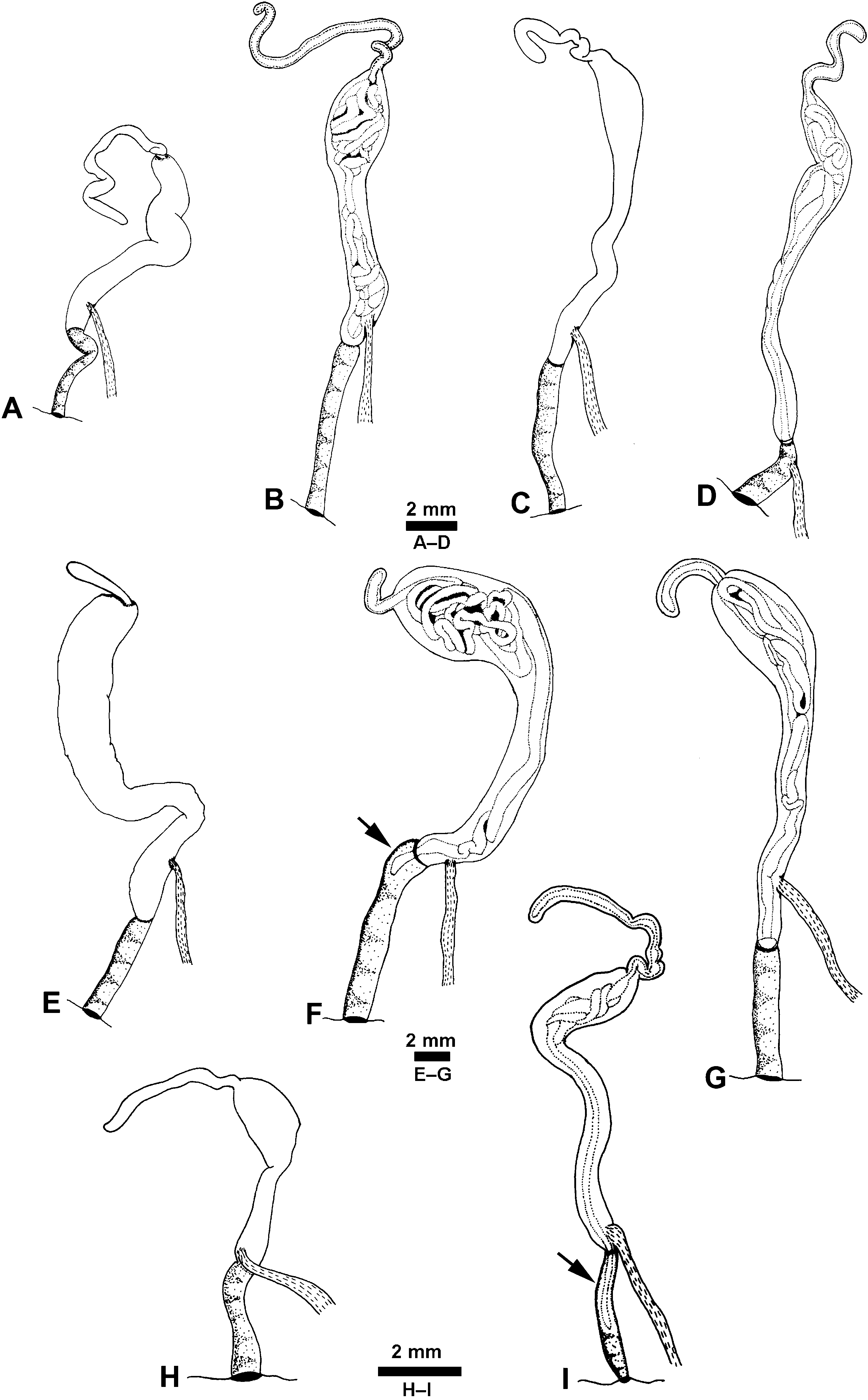
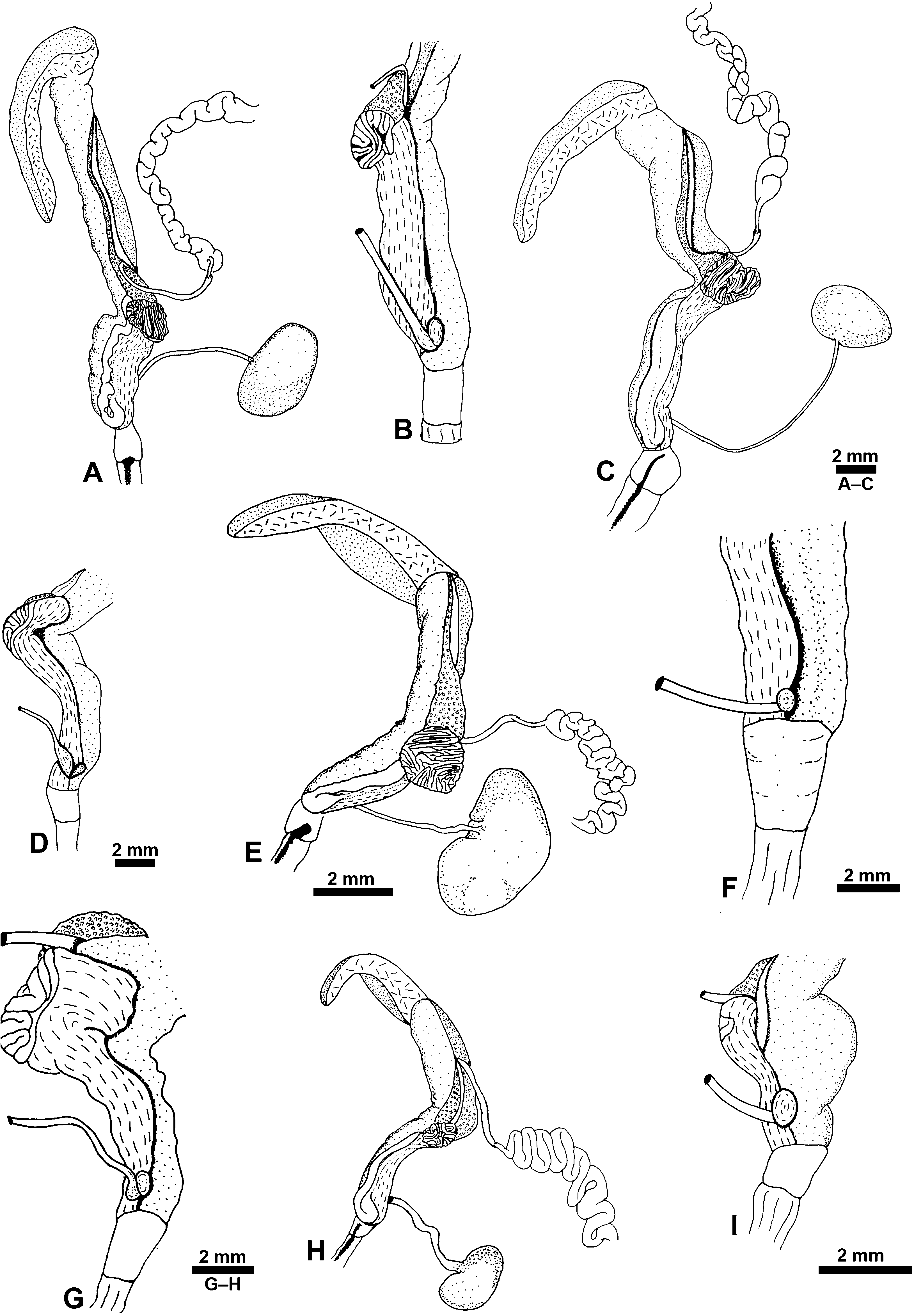


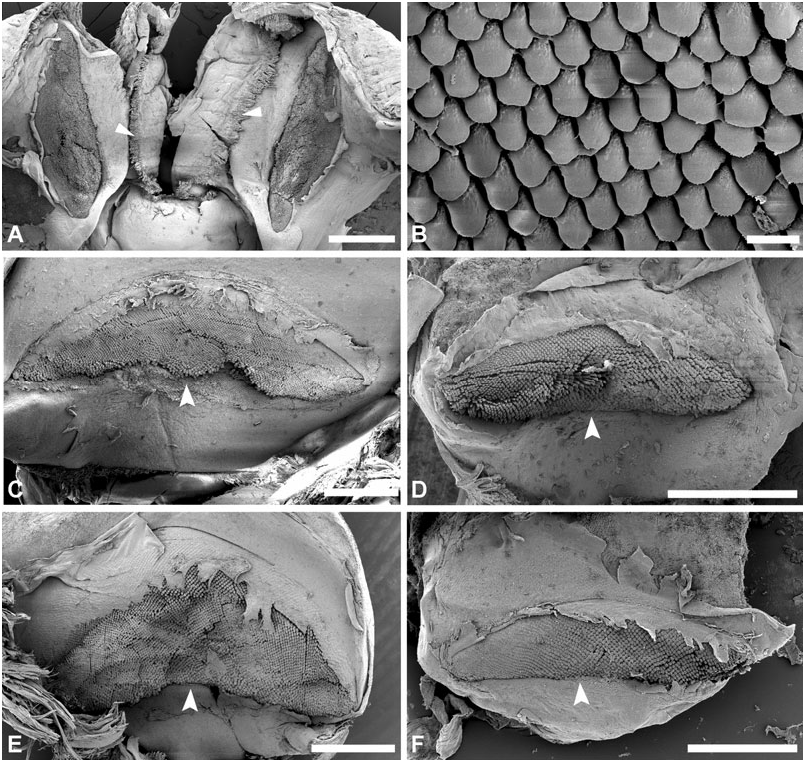



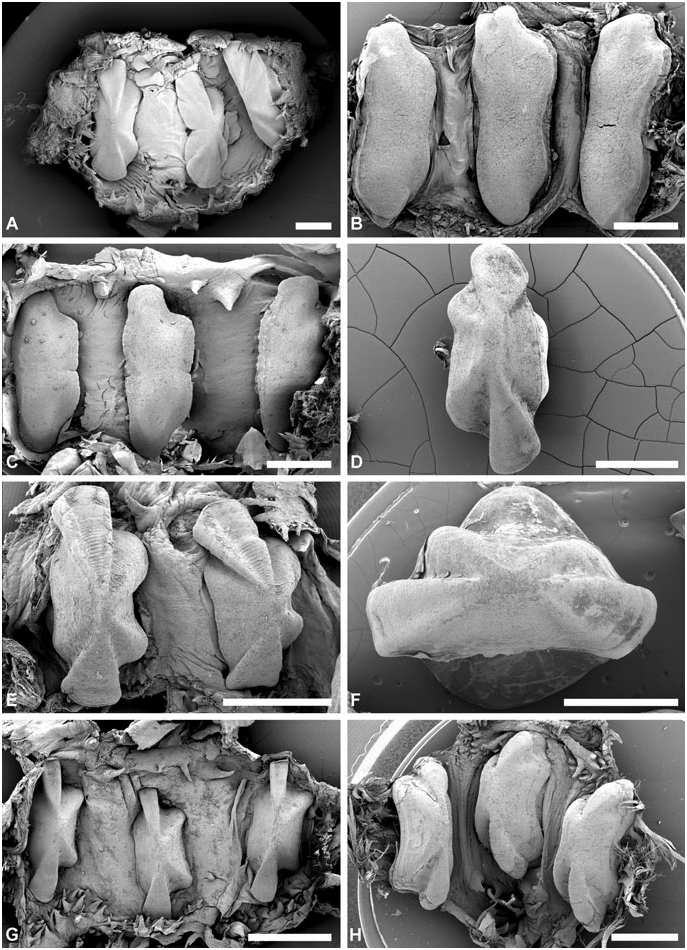
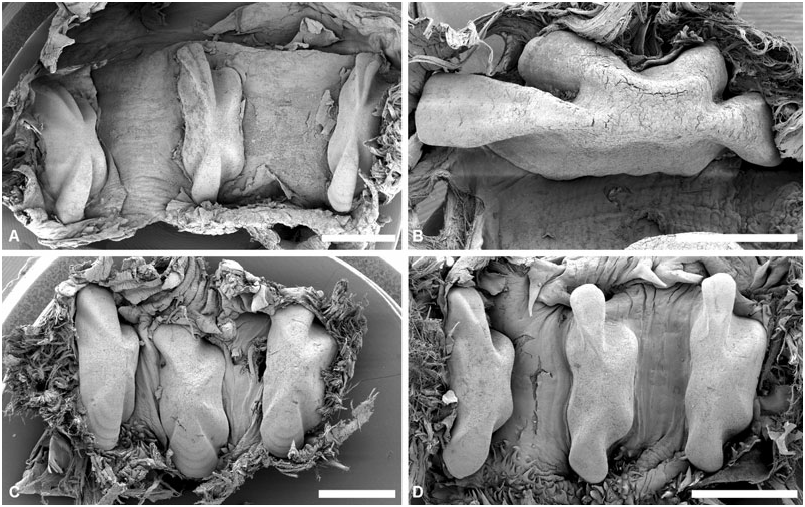
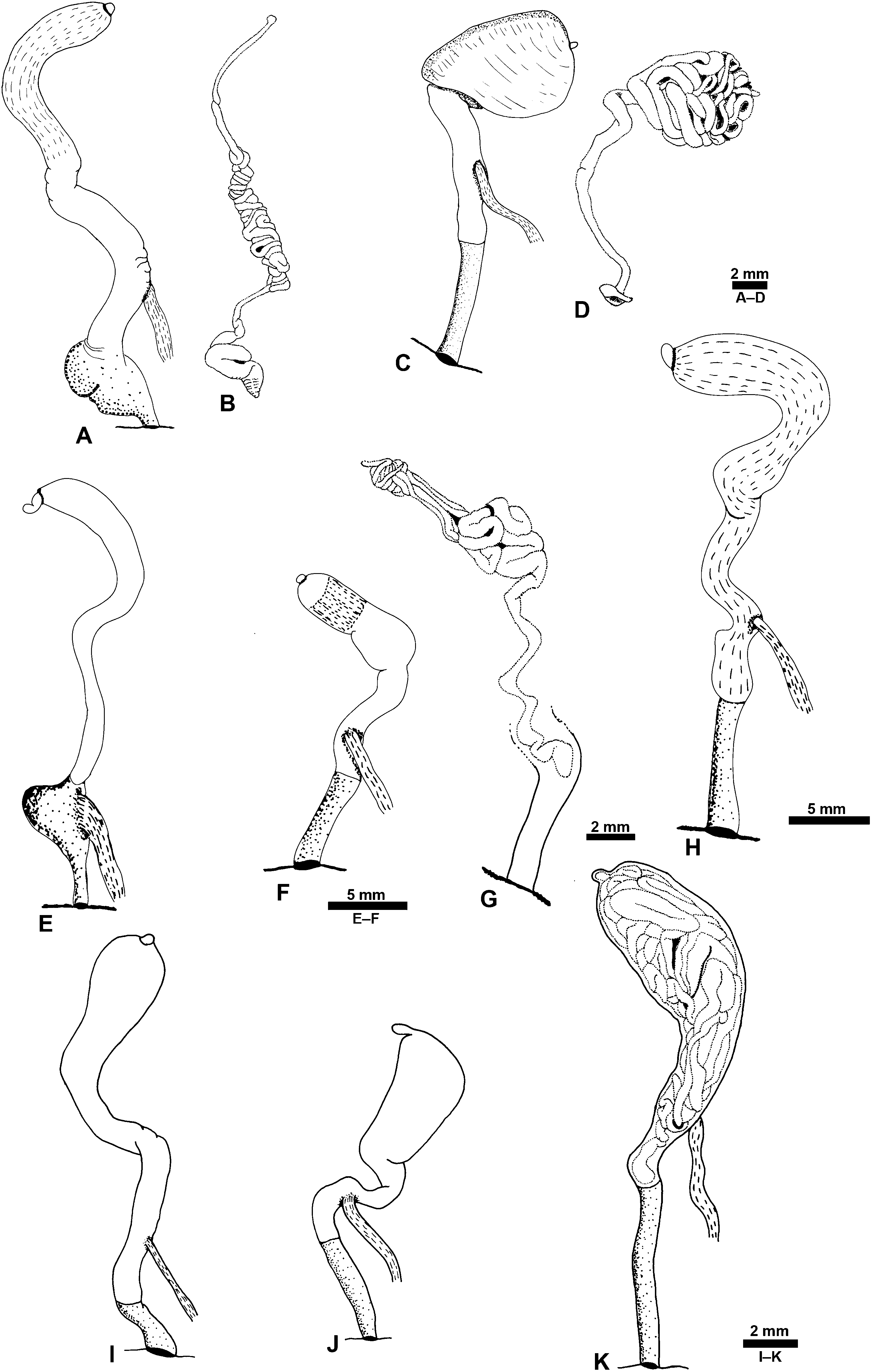
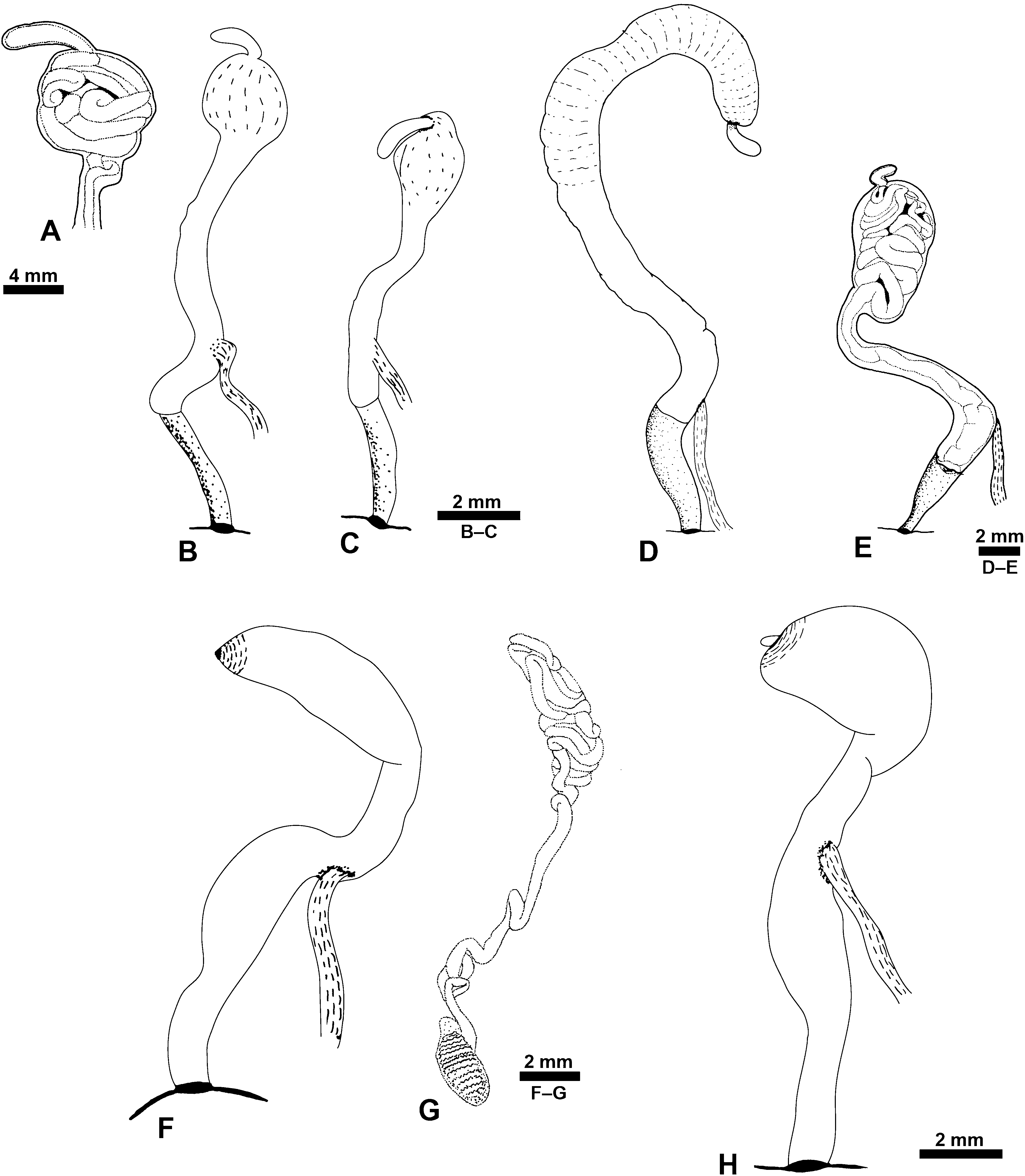



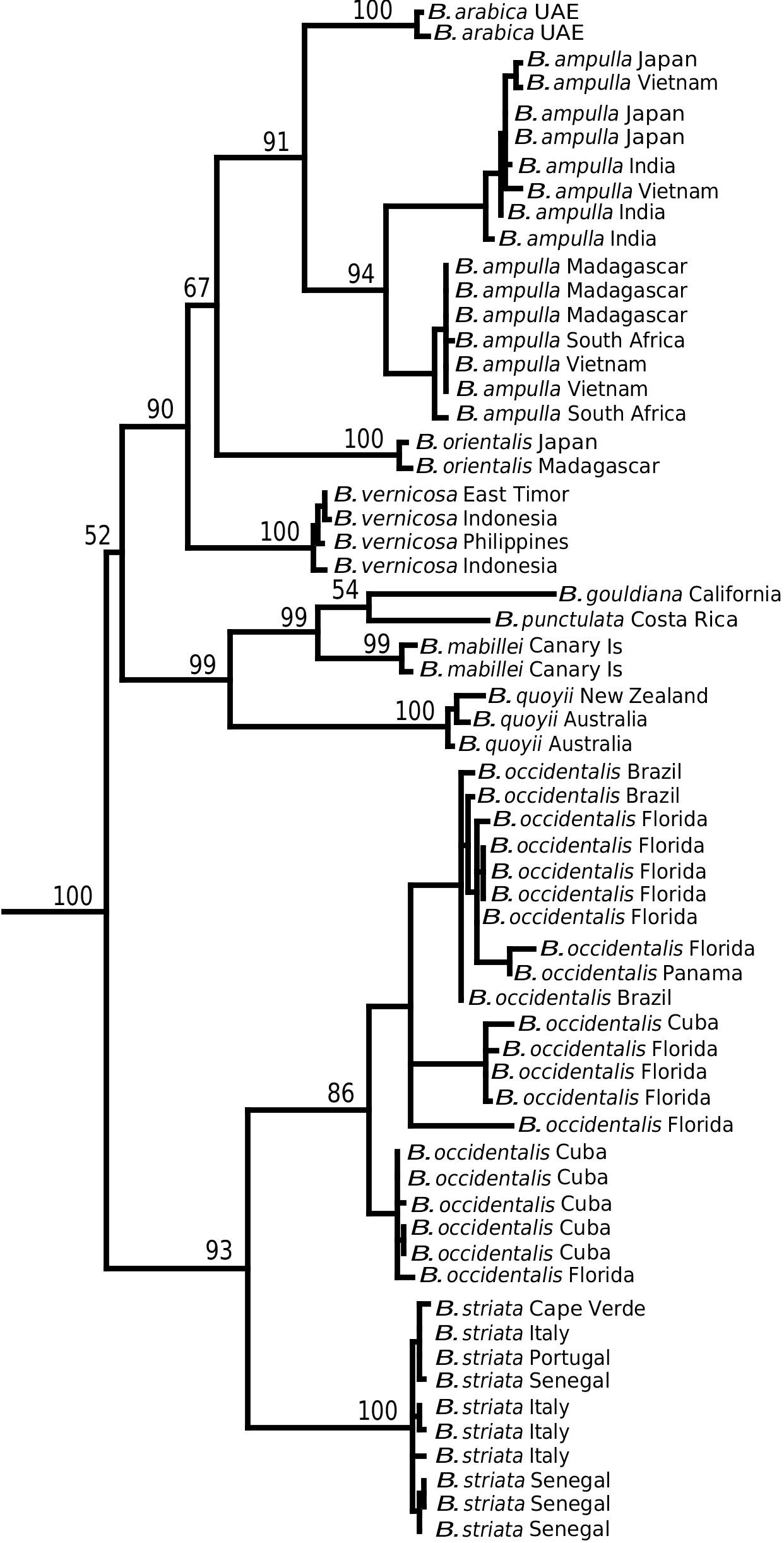
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Bulla
| Malaquias, Manuel António E. & Reid, David G. 2008 |
Bulla punctulata
| Thach NN 2005: 230 |
Bulla ovula
| Higo S & Callomon P & Goto Y 2001: 141 |
| Mabille MJ 1896: 116 |
Bulla ventricosa
| Hori S 2000: 758 |
Bulla ampulla var. bifasciata
| Guangyu L 1997: 95 |
Bulla ampulla
| Fukuda H 1995: 31 |
Bullaria ampulla
| Green S 1994: 71 |
Bulla cruentata
| Drivas J & Jay M 1988: 130 |
Bullaria striata
| Nordsieck F & Garcia-Talavera F 1979: 173 |
Bulla vernicosa
| Kay EA 1979: 423 |
Bulla (Bulla) gouldiana
| Keen AM 1971: 794 |
Bulla (Bulla) rufolabris
| Keen AM 1971: 794 |
Bulla (Bulla) punctulata
| Keen AM 1971: 794 |
Retusa mariateresae
| Parenzan P 1970: 220 |
Bulla (Quibulla) subtropicalis
| Powell AWB 1965: 167 |
Quibulla tenuissima
| Cotton BC 1959: 404 |
Bulla orientalis Habe, 1950: 21
| Burn R 1966: 97 |
| Kira T 1962: 114 |
| Habe T 1950: 21 |
Bulla difficilis
| Thach NN 2005: 230 |
| Hori S 2000: 758 |
| Guangyu L 1997: 94 |
| Habe T 1950: 21 |
Bullaria adansoni
| Marche-Marchad I 1958: 39 |
| Nickles M 1950: 136 |
| Fischer-Piette E 1942: 128 |
Bulla peasiana
| Kuroda T & Habe T 1952: 42 |
| Kuroda T 1941: 133 |
Bulla striata var. minor
| Pallary P 1938: 15 |
Bullaria (Quibulla) tenuissima
| Cotton BC & Godfrey FK 1938: 32 |
Quibulla scotti
| Iredale T 1937: 258 |
Quibulla
| Iredale T 1929: 349 |
Bulla adamsi
| Michel C 1985: 87 |
| Dautzenberg P 1929: 334 |
Quibulla selina
| Iredale T 1929: 349 |
Bulla adamsi
| Eisenberg JM 1981: 152 |
| Dautzenberg P 1929: 334 |
Quibulla botanica
| Iredale T 1929: 349 |
Bulla angasi
| Cobb G & Willan RC 2006: 30 |
| Jansen P 2000: 70 |
| Short JW & Potter DG 1987: 116 |
| Willan RC 1978: 60 |
| Iredale T 1929: 350 |
Bullaria gouldiana
| Baker F & Hanna GD 1927: 127 |
Bullaria punctulata
| Baker F & Hanna GD 1927: 127 |
Bullaria occidentalis
| Peile AJ 1926: 85 |
Bullaria botanica
| Macpherson JH & Gabriel CJ 1962: 242 |
| May WL 1921: 103 |
Bullaria quoyana
| Dall WH 1919: 300 |
Bullaria australis
| Hedley C 1916: 72 |
Bullaria quoyi
| Hedley C 1916: 72 |
Bullaria tenuissima
| Hedley C 1916: 72 |
Bullaria peasiana
| Oliver WRB 1915: 542 |
Bullaria australis
| Suter H 1913: 534 |
Bullaria australis quoyi
| Suter H 1913: 535 |
Bullaria adamsi
| Suter H 1913: 534 |
Bulla adamsi
| Bergh R 1900: 216 |
Bulla ampulla var. mozambicensis
| Bergh R 1900: 210 |
Bulla ampulla
| Bergh R 1900: 210 |
Bulla occidentalis
| Nordsieck F 1972: 28 |
| Locard A 1897: 51 |
Bulla mabillei Locard, 1897: 50
| Rolan E 2005: 389 |
| Ardovini R & Cossignani T 2004: 242 |
| Malaquias MAE & Martinez E & Abreu AD 2002: 73 |
| Valdes A & Heros V 1998: 709 |
| Malaquias MAE & Calado GJP 1997: 154 |
| Wirtz P 1995: 184 |
| Sanchez JMP & Batet EM 1991: 218 |
| Bernard PA 1984: 108 |
| von Cosel R 1982: 31 |
| von Cosel R 1982: 21 |
| von Cosel R 1982: 61 |
| Talavera FG 1978: 126 |
| Nordsieck F 1972: 28 |
| Marcus Ev & Marcus Er 1966: 155 |
| Marche-Marchad I 1958: 39 |
| Marche-Marchad I 1956: 59 |
| Odhner NH 1932: 25 |
| Locard A 1897: 50 |
Bulla quoyi
| Mabille MJ 1896: 115 |
Bulla delorti
| Valdes A & Heros V 1998: 708 |
| Mabille MJ 1896: 116 |
Bulla dubiosa
| Mabille MJ 1896: 113 |
Bulla secunda
| Valdes A & Heros V 1998: 710 |
| Mabille MJ 1896: 112 |
Bulla subaustralis
| Mabille MJ 1896: 114 |
Bulla roperiana
| Clench WJ & Turner RD 1962: 130 |
| Pilsbry HA 1895: 336 |
Bulla amygdala
| Nordsieck F 1972: 28 |
| Perry LM & Schwengel JS 1955: 192 |
| Dall WH & Simpson CT 1901: 363 |
| Pilsbry HA 1895: 329 |
Bulla angasi
| Pilsbry HA 1895: 347 |
Bulla gouldiana Pilsbry, 1895: 340
| Gosliner TM & Williams GC 2007: 785 |
| Mclean JH 2007: 751 |
| Behrens DW & Hermosillo A 2005: 27 |
| Mikkelsen PM 1996: 436 |
| Stebbins TD 1987: 95 |
| Eisenberg JM 1981: 152 |
| Robles LJ 1975: 279 |
| Abbott RT 1974: 319 |
| Marcus Ev & Marcus Er 1967: 143 |
| Johnson RI 1964: 84 |
| Marcus Er 1961: 4 |
| Pilsbry HA 1895: 340 |
Bulla peasiana Pilsbry, 1895: 348
| Kay EA 1965: 44 |
| Pilsbry HA 1895: 348 |
Bulla australis var. oblonga
| Pilsbry HA 1895: 346 |
B. tenuissima
| Pilsbry HA 1895: 347 |
Bulla punctata
| Melvill JC & Standen R 1897: 396 |
| Whitelegge T 1889: 275 |
Bulla ampulla
| Mienis HK 2004: 33 |
| Zenetos A & Gofas S & Russo G & Templado J 2003: 172 |
| Bosch DT & Dance SP & Moolenbeek RG & Oliver PG 1995: 179 |
| Coulombel A 1994: 112 |
| Barash A & Danin Z 1992: 186 |
| Bosch D & Bosch E 1982: 137 |
| Eales NB 1938: 81 |
| Vayssiere MA 1906: 18 |
| Melvill JC & Standen R 1901: 456 |
| Cooke AH 1886: 130 |
| Watson RB 1886: 637 |
Bulla striata
| Monterosato M 1884: 141 |
Bulla striata var. solida
| Monterosato M 1884: 141 |
Bulla striata var. stricta
| Monterosato M 1884: 141 |
Bulla trifasciata
| Guangyu L 1997: 95 |
| Mabille MJ 1896: 112 |
| Brazier J 1878: 83 |
Bullea solida
| Morch OAL 1875: 173 |
Bulla conspersa
| Pilsbry HA 1895: 349 |
| Pease WH 1869: 72 |
Bulla punctulata
| Cobb G & Willan RC 2006: 30 |
| Nakano R 2004: 30 |
| Hori S 2000: 758 |
| Guangyu L 1997: 93 |
| Fukuda H 1995: 31 |
| Short JW & Potter DG 1987: 116 |
| Salvat B & Rives C 1975: 177 |
| Cernohorsky WO 1972: 207 |
| Hedley C 1907: 486 |
| Melvill JC & Standen R 1899: 155 |
| Brazier J 1879: 198 |
| Angas GF 1867: 226 |
Bulla magdelus
| Angas GF 1867: 227 |
Bulla ovulum
| Burn R 1966: 96 |
| Angas GF 1867: 227 |
Bulla marmorea
| Kay EA 1965: 44 |
| Mabille MJ 1896: 116 |
| Pease WH 1860: 431 |
Bulla vernicosa Gould, 1859: 138
| Cobb G & Willan RC 2006: 32 |
| Dharma B 2005: 206 |
| Thach NN 2005: 230 |
| Trondle J & von Cosel R 2005: 320 |
| Burn R & Thompson TE 1998: 958 |
| Guangyu L 1997: 92 |
| Fukuda H 1994: 40 |
| Dharma B 1992: 76 |
| Short JW & Potter DG 1987: 116 |
| Springsteen FJ & Leobrera FM 1986: 284 |
| Lin G & Qi Z 1985: 114 |
| Powell AWB 1979: 275 |
| Willan RC 1978: 63 |
| Powell AWB 1976: 110 |
| Cernohorsky WO 1972: 207 |
| Kuroda T & Hhabe T & Oyama K 1971: 288 |
| Johnson RI 1964: 166 |
| Habe T 1950: 21 |
| Pilsbry HA 1895: 349 |
| Gould AA 1859: 138 |
Bulla amygdala
| Macedo MCC & Macedo MIC & Borges JP 1999: 255 |
| Poppe GT & Goto Y 1991: 195 |
| Nordsieck F 1972: 28 |
| Menke KT 1854: 44 |
Bulla adansoni
| Ardovini R & Cossignani T 2004: 242 |
| Rolan E & Ryall P 1999: 58 |
| Bernard PA 1984: 108 |
| Nordsieck F 1972: 28 |
| Dautzenberg P 1910: 12 |
| Mabille MJ 1896: 118 |
| Menke KT 1854: 44 |
Bulla columellaris
| Hedley C 1916: 72 |
| Iredale T 1914: 667 |
| Menke KT 1854: 26 |
| Menke KT 1854: 43 |
Bulla columellaris var. bifasciata
| Shirley J 1911: 102 |
| Mabille MJ 1896: 112 |
| Menke KT 1854: 26 |
Bulla columellaris var. vermiculata
| Menke KT 1854: 26 |
Bulla adamsi
| Guangyu L 1997: 94 |
| Oostingh CH 1923: 139 |
| Suter H 1913: 534 |
| Hedley C 1910: 370 |
| Melvill JC & Standen R 1899: 155 |
| Pilsbry HA 1895: 345 |
| Menke KT 1854: 43 |
Bulla adansonii var. minor
| Dunker G 1853: 4 |
Bulla (Bullea) dactylis
| Nordsieck F 1972: 28 |
| Menke KT 1853: 137 |
Bulla (Bullea) perdicinia
| Pilsbry HA 1895: 335 |
| Menke KT 1853: 140 |
Bulla (Bullea) omphalodes
| Menke KT 1853: 137 |
Bulla (Bullea) nux
| Pilsbry HA 1895: 350 |
| Menke KT 1853: 140 |
Bulla (Bullea) sulcata
| Menke KT 1853: 138 |
B. substriata
| Menke KT 1853: 136 |
Bulla rubiginosa
| Mabille MJ 1896: 119 |
| Pilsbry HA 1895: 330 |
| Gould AA 1852: 221 |
Bulla occidentalis
| Redfern C 2001: 158 |
| Abbott RT & Dance SP 1991: 279 |
| Damerval M & Damerval D 1990: 190 |
| Eisenberg JM 1981: 152 |
| Marcus Ev 1976: 4 |
| Nordsieck F 1972: 28 |
| Rios EC 1970: 131 |
| Vilas CN & Vilas NR 1970: 91 |
| Perry LM & Schwengel JS 1955: 192 |
| Johnson C 1934: 148 |
| Peile AJ 1926: 85 |
| Mabille MJ 1896: 114 |
| Pilsbry HA 1895: 331 |
| Dall WH 1889: 55 |
| Menke KT 1854: 45 |
| Adams A 1850: 577 |
Bulla cruentata
| Mabille MJ 1896: 116 |
| Adams A 1850: 577 |
Bulla nebulosa
| Johnson RI 1964: 115 |
| Bergh R 1900: 214 |
| Mabille MJ 1896: 115 |
| Carpenter PP 1857: 173 |
| Menke KT 1854: 44 |
| Adams A 1850: 578 |
| Menke KT 1850: 162 |
Bulla quoyii
| Pilsbry HA 1895: 342 |
| Adams A 1850: 576 |
Bulla rufolabris
| Mabille MJ 1896: 115 |
| Menke KT 1854: 44 |
| Adams A 1850: 577 |
Bulla punctata Adams, 1850: 577–578
| Mabille MJ 1896: 115 |
| Adams A 1850: 578 |
| Adams A 1850: 604 |
Bulla punctulata Adams, 1850: 577–578
| Behrens DW & Hermosillo A 2005: 28 |
| Paredes C & Cardoso F & Tarazona J 2004: 213 |
| Valdes A & Camacho-Garcia YE 2004: 488 |
| Higo S & Callomon P & Goto Y 2001: 141 |
| Hickman CP & Finet Y 1999: 116 |
| Emerson WK 1995: 18 |
| Montoya M 1983: 343 |
| Eisenberg JM 1981: 152 |
| Abbott RT 1974: 319 |
| Morris PA 1973: 204 |
| Sphon GG & Mulliner DK 1972: 149 |
| Bartsch P & Rehder HA 1939: 15 |
| Pilsbry HA 1895: 341 |
| Stearns REC 1893: 376 |
| Menke KT 1854: 44 |
| Adams A 1850: 578 |
Bulla aspersa
| Mabille MJ 1896: 115 |
| Pilsbry HA 1895: 341 |
| Menke KT 1854: 44 |
| Adams A 1850: 578 |
Bulla adamsi
| Stearns REC 1894: 158 |
| Carpenter PP 1857: 172 |
| Menke KT 1850: 162 |
Bulla oblonga
| Pilsbry HA 1895: 346 |
| Angas GF 1867: 226 |
| Menke KT 1854: 44 |
| Adams A 1850: 577 |
Haminea castanea
| Adams A 1850: 584 |
Bulla australis
| Adams A 1850: 576 |
Bulla panamensis
| Pilsbry HA 1895: 343 |
| Menke KT 1850: 162 |
| Philippi RA 1848: 141 |
Bulla adansonii
| Pilsbry HA 1895: 333 |
| Adams A 1850: 576 |
| Philippi RA 1847: 121 |
Bulla media
| Bergh R 1900: 217 |
| Mabille MJ 1896: 118 |
| Adams A 1850: 576 |
| Philippi RA 1847: 121 |
Bulla quoyii
| Burn R 2006: 7 |
| Jansen P 2000: 70 |
| Burn R & Thompson TE 1998: 957 |
| Burn R 1989: 737 |
| Wells FE & Bryce CW 1988: 136 |
| Burn R 1984: 68 |
| Powell AWB 1979: 275 |
| Willan RC 1978: 58 |
| Pilsbry HA 1895: 348 |
| Menke KT 1854: 44 |
| Gray JE 1843: 243 |
Vesica
| Swainson W 1840: 360 |
Vesica ampulla
| Swainson W 1840: 360 |
Bulla ampulla
| Guerreiro A & Reiner F 2000: 209 |
| d'Orbigny A 1839: 45 |
Bulla australis
| Valdes A & Heros V 1998: 708 |
| Suter H 1913: 534 |
| Shirley J 1911: 102 |
| Mabille MJ 1896: 113 |
| Watson RB 1886: 638 |
| Brazier J 1878: 83 |
| Menke KT 1854: 44 |
| Deshayes GP & Milne-Edwards H 1836: 673 |
| Quoy JRC & Gaimard JP 1833: 357 |
Bulla striata
| Pilsbry HA 1895: 348 |
| Quoy JRC & Gaimard JP 1833: 354 |
Bulla columnae
| Chiaje S 1827: 24 |
Bullea
| de Blainville HMD 1825: 478 |
Bulla australis
| Iredale T 1929: 349 |
| Torr CM 1914: 366 |
| Pritchard GB & Gatliff JH 1902: 214 |
| Pilsbry HA 1895: 346 |
| Whitelegge T 1889: 275 |
| Brazier J 1885: 89 |
| Gray JE 1843: 243 |
| Gray JE 1826: 490 |
| Gray JE 1825: 408 |
Bulla australis Férussac, 1822: 575
| Valdes A & Heros V 1998: 708 |
| Ferussac D 1822: 575 |
Bulla amygdalus
| Dillwyn LW 1817: 480 |
Bulla amygdalus
| Adams A 1850: 575 |
| Dillwyn LW 1817: 480 |
Bullaria
| Rafinesque CS 1815: 142 |
Bullus
| de Montfort D 1810: 331 |
Bulla nebulosa Schröter, 1804: 20
| Schroter JM 1804: 20 |
Bulla marmorea Schröter, 1804: 20
| Schroter JM 1804: 20 |
Bulla umbilicata Röding, 1798: 15
| Roding PF 1798: 15 |
Bulla ampulla striata Röding, 1798: 15
| Cernohorsky WO 1972: 207 |
| Roding PF 1798: 15 |
Bulla ovum vanelli Röding, 1798: 15
| Cernohorsky WO 1972: 207 |
| Roding PF 1798: 15 |
Bulla fasciata Röding, 1798: 15
| Roding PF 1798: 15 |
Bulla bulla rubra Röding, 1798: 15
| Roding PF 1798: 15 |
Bulla flammea Röding, 1798: 16
| Roding PF 1798: 16 |
Bulla striata Bruguière, 1792: 372
| Rolan E 2005: 203 |
| Costa F & Costa M & Salpietro L & Turavano F 2002: 128 |
| Guerreiro A & Reiner F 2000: 211 |
| Malaquias MAE & Morenito P 2000: 119 |
| Macedo MCC & Macedo MIC & Borges JP 1999: 255 |
| Rolan E & Ryall P 1999: 58 |
| Murillo L 1996: 36 |
| Rios E 1994: 197 |
| Fernandes F & Rolan E 1993: 41 |
| Barash A & Danin Z 1992: 186 |
| Poppe GT & Goto Y 1991: 195 |
| Barash A & Danin Z 1989: 250 |
| Garcia-Talavera CF 1983: 34 |
| Luque AA 1983: 55 |
| Abbott RT & Dance SP 1982: 279 |
| von Cosel R 1982: 21 |
| von Cosel R 1982: 61 |
| Templado J 1982: 249 |
| Eisenberg JM 1981: 153 |
| Marcus Ev 1977: 4 |
| Marcus Ev 1976: 125 |
| Nordsieck F 1972: 27 |
| Pasteur-Humbert C 1962: 175 |
| Nobre A 1932: 57 |
| Si T 1931: 29 |
| Mabille MJ 1896: 116 |
| Pilsbry HA 1895: 332 |
| Smith EA 1890: 296 |
| Lamarck & JBPAdeM 1822: 33 |
| Bruguiere JG 1792: 372 |
Bulla striata Bruguière, 1792: 372
| Mikkelsen PM 1996: 436 |
| Andrews J 1994: 75 |
| Diaz JM & Puyana M 1994: 244 |
| Rios E 1994: 197 |
| Sterrer W 1992: 142 |
| Garcia-Talavera CF 1983: 34 |
| Vokes HE & Vokes EH 1983: 33 |
| Abbott RT & Dance SP 1982: 279 |
| Eisenberg JM 1981: 153 |
| Andrews J 1977: 162 |
| Marcus Ev 1977: 4 |
| Marcus Ev 1976: 125 |
| Abbott RT 1974: 319 |
| Nordsieck F 1972: 27 |
| Marcus Er 1957: 390 |
| Perry LM & Schwengel JS 1955: 192 |
| Johnson C 1934: 148 |
| Pilsbry HA 1895: 332 |
| Lamarck & JBPAdeM 1822: 33 |
| Bruguiere JG 1792: 372 |
Bulla solida
| Mikkelsen PM 1996: 436 |
| Diaz JM & Puyana M 1994: 244 |
| Marcus Ev 1977: 4 |
| Bandel K 1976: 89 |
| Marcus Ev 1976: 121 |
| Morris PA 1973: 264 |
| Johnson C 1934: 148 |
| Pilsbry HA 1895: 335 |
| Krebs H 1864: 93 |
| Gmelin FJ 1791: 3434 |
Bulla ampulla
| Dharma B 2005: 206 |
| Thach NN 2005: 230 |
| Hori S 2000: 758 |
| Apte D 1998: 85 |
| Burn R & Thompson TE 1998: 957 |
| Steyn DG & Lussi M 1998: 176 |
| Guangyu L 1997: 93 |
| Dharma B 1992: 76 |
| Drivas J & Jay M 1988: 130 |
| Gosliner TM 1987: 45 |
| Short JW & Potter DG 1987: 116 |
| Springsteen FJ & Leobrera FM 1986: 284 |
| Lin G & Qi Z 1985: 115 |
| Kirtisinghe P 1978: 57 |
| Cernohorsky WO 1972: 207 |
| Macnae W 1962: 185 |
| Dodge H 1955: 23 |
| Risbec J 1951: 143 |
| Habe T 1950: 21 |
| Dautzenberg P 1929: 335 |
| Shirley J 1911: 102 |
| Iredale T 1910: 71 |
| Bergh R 1900: 210 |
| Mabille MJ 1896: 111 |
| Pilsbry HA 1895: 343 |
| Whitelegge T 1889: 275 |
| Watson RB 1886: 637 |
| Angas GF 1877: 189 |
| Hanley S 1855: 204 |
| Adams A 1850: 575 |
| Deshayes GP & Milne-Edwards H 1836: 668 |
| Lamarck & JBPAdeM 1822: 33 |
| de Montfort D 1810: 330 |
| Bruguiere JG 1792: 371 |
| da Costa EM 1776: 174 |
| Linnaeus C 1758: 727 |