
Nubensia nubens (Edwards) Edwards, 2015
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3994.1.5 |
|
publication LSID |
lsid:zoobank.org:pub:22F78B32-A529-474D-9ED4-C2F683F41433 |
|
DOI |
https://doi.org/10.5281/zenodo.6096487 |
|
persistent identifier |
https://treatment.plazi.org/id/255E882C-FFD9-9266-FF70-A707C44ABD61 |
|
treatment provided by |
Plazi |
|
scientific name |
Nubensia nubens (Edwards) |
| status |
comb. nov. |
Nubensia nubens (Edwards) View in CoL , n. comb.
(Figs 2–6)
Selected references to relevant names and material
Pentapedilum (Pentapedilum) nubens Edwards, 1929: 376 [original publication, description of adult male and female]
Chironomini View in CoL type A—Hofmann (1991: 366 and fig. 1) [larva]
Chironomini View in CoL A—Hofmann (1993: 422) [larva]
Polypedilum View in CoL type A—Brooks et al. (2007: 102 and fig. 6.51) [larva]; Cid Puey (2010) [record of larvae]
Polypedilum (Pentapedilum) nubens (Edwards) — Langton & Visser (2003) [pupal exuviae], Langton & Pinder (2007) [adult male], Oyewo & Saether (2008) [adult male, pupa], Saether & Spies (2013) [distribution]
Genus sp. near Tribelos View in CoL — Chaib et al. (2013) [record of larvae]
Material examined. Besides the specimens itemized below, see also the ZSM material recorded in Oyewo & Saether (2008: 82). Additional material from Germany, Greece, Italy, Morocco and Turkey is present at ZSM.
(1) UNITED KINGDOM. England, Westmorland, Lake Windermere, vi.1929, F.W. Edwards—2 syntype adults (now on slides; NHM): 1 male, labelled " BMNH (E) 236289"; 1 female, " BMNH (E) 236284".
(2) GERMANY. Rheinland-Pfalz, Wörth am Rhein (NW of Karlsruhe), Landeshafen Wörth, W shore, coordinates 49.069025 N, 8.307178 E, and 49.062445 N, 8.303401 E; 2.iv.2013, leg. A. Dettinger-Klemm—4 fourth instar + 12 third instar larvae (3+4 at ZSM, remainder in coll. Dettinger-Klemm); as previous, except vii.2013, 2 pharate males in exuviae with attached larval skins, 2 pharate males in exuviae, 1 pharate female in exuviae with attached larval skin (at ZSM). Baden-Württemberg, Bodensee (Lake Constance), Untersee at Gaienhofen-Horn, rubble zone exposed to surf, substrate with coarse gravel, much mud and plant material, adjacent to a reed bed; 12.vii.1993, leg. M. Steinhart—dissected parts of 1 fourth instar larva (at ZSM). Bavaria, Bodensee, Nonnenhorn, Paradies fish breeding station, 24.vii.1963, leg. F. Reiss— 2 males (at ZSM).
(3) ALGERIA, Bechar, Oued Zousfana (a wadi), drift sample (No. 57); 12.iv.1955, leg. E.J. Fittkau—dissected parts of 2 skins of fourth instar larvae (at ZSM).
Larval morphology (Figs 2–4). Data for fourth instar (imaginal discs clearly visible in some specimens), except where stated otherwise.
Size: fourth instar body length up to about 6 mm, head capsule length/width up to 450/335 µm, postmentum length 210, 215 µm ( Algeria, n=2) / 225–233 µm ( Germany, n=4), third instar postmentum length c. 145 µm ( Germany, n=1).
Coloration ( Fig. 1 View FIGURE 1 ): whole larva in life red-brown to the unaided eye, at stereoscope magnification marmorated vividly red and bright green. Head capsule mostly medium to dark brown with lighter colored antennae, labrum, bases of mandibles, areas around eyespots, two strips flanking the frontal apotome, and a transverse strip in the anterior gular region; two eyespots per side, narrowly separated to slightly fused.
Dorsal surface of head (Fig. 2a, b): frons tapering posteriorly, with slight constriction near anterior end, anterolateral corners acute, anterior margin straight, anteromedial 'fenestra' absent; clypeal sclerite slightly wider than frons, uniformly sclerotized, its posterolateral corners carrying setae S3.
Antenna (Fig. 2d): 5-segmented; AR 1.17 ( Germany, Bodensee, n=1) / 1.27, 1.32 ( Germany, Landeshafen Wörth, n=2) / 1.32, 1.42 ( Algeria, n=2), third instar AR 0.92, 1.08 ( Germany, Landeshafen Wörth, n=2); segment lengths (in µm, sclerotized sections only) c. 50/15.5/7/7.5/5 (Bodensee) / c. 46.5/13.5/6.5/7/4.5 (Landeshafen) / c. 46/13/5 /5.5/3 ( Algeria); ring organ near proximal end of segment 1, frequently indistinct; antennal seta absent, small unpigmented spot frequently visible near distal end of segment 1; blade inserted on top of segment 1 next to segment 2, reaching well beyond tip of segment 5; top of segment 2 with small accessory blade, and two small Lauterborn organs in opposite positions.
Labrum and epipharynx (Fig. 2b, c, f): seta S2 not arising from any distinct larger sclerite, S1 inserted on labral sclerite; seta S IVA a short peg, setae S III arising from adjacent sockets just posteromedially of the S II, hair-like, not very fine and short; S II wide and long, apically with a few short splits, setae S I arising from adjacent sockets, shorter but wider than S II, feathered on both edges in distal half; labral lamella absent; seta praemandibularis simple, rather strong and long; pecten epipharyngis of three closely adjacent plates forming 4–5 + 2 + 4–5 rather strong teeth, accessory teeth and dorsal spinules absent; premandible (Fig. 2f) with a very short proximal tooth and two long distal teeth, all with blunt tips, distal half of premandible with conspicuous medial 'beard'.
Mandible (Fig. 2e): subapical dorsal tooth present, strong and dark; apical tooth strong, about as long as combined width of the three inner teeth, all of them dark; pecten mandibularis absent; seta subdentalis simple, longer than apical tooth, reaching beyond proximal inner tooth, SSd origin on ventral side of mandible, about as far from distal end of mola as 4/5 of combined width of the three inner teeth; seta interna originating on dorsal side of mandible, consisting of four basally separate, distally weakly feathered branches; medial edge of mola with 1–2 fine spinose projections, outer edge of mandible weakly crenulate; proximal part of mandible without grooves like those in the Chironomus complex.
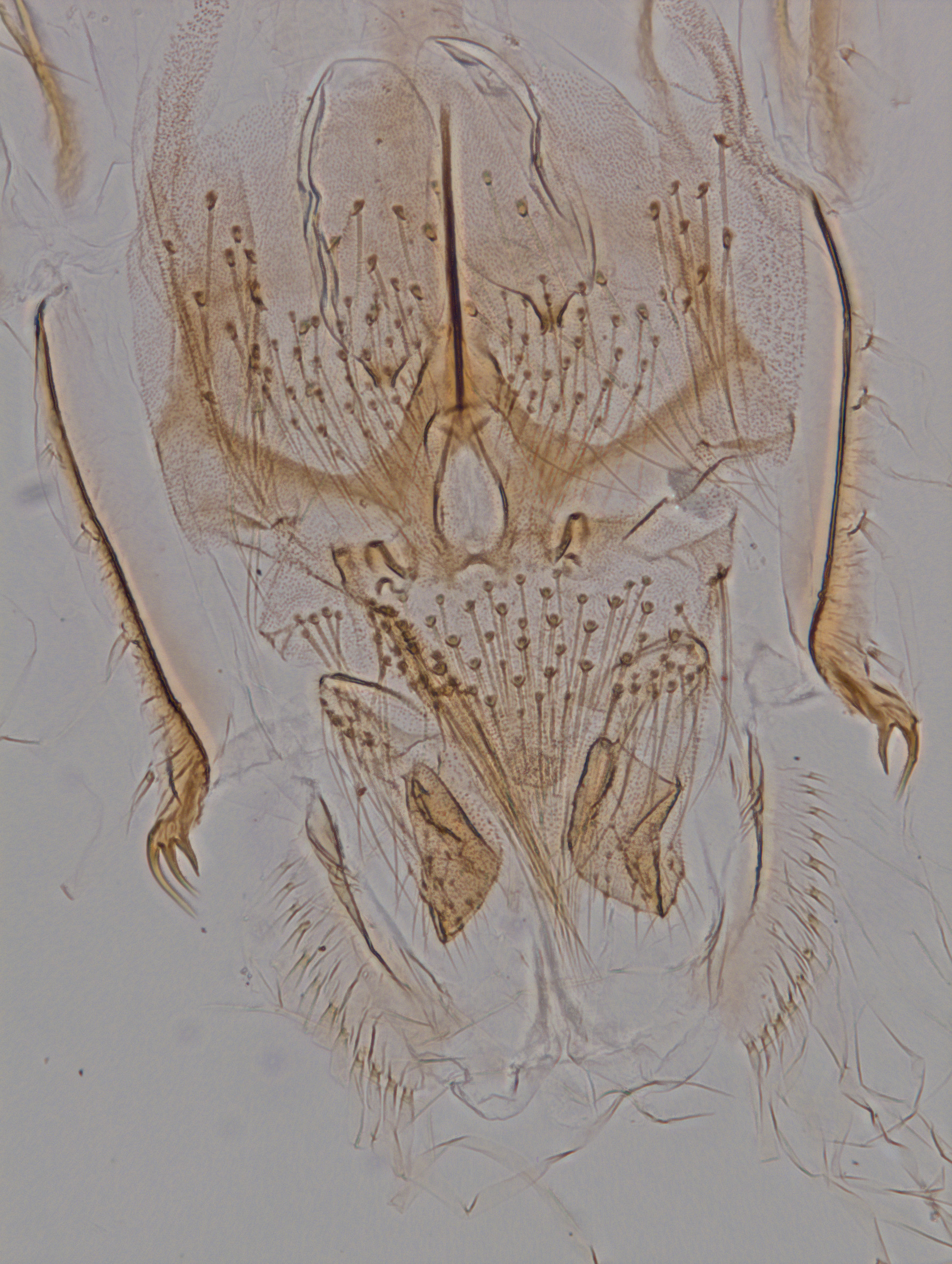
Mentum ( Fig 4 View FIGURE 4 ): entirely dark, often with longitudinal, irregular, coarse striae posterior to the teeth, anterior edge of mentum shallowly convex, with 14 teeth (exceptionally with a minute, recessed medial 15th tooth), the medial pair of teeth strikingly enlarged, the far lateral pair very small; ventromental plate fan shaped, (longitudinal) height/(transverse) width> 0.5, about as wide as 3/4 of mentum, with 30–35 striae that divide radial lamellae with rounded ends dorsal to the anterior plate margin, the latter smooth, medial tips of the two plates widely separated, pointing to anteromedial but short, without long contours leading to bases of mentum teeth. Seta submenti simple, not reaching to anterior margin of ventromental plate.
Body ( Fig. 1 View FIGURE 1 ): abdominal segments VII and VIII without lateral and ventral tubuli; procercus with 6–7 terminal setae that are longer than the anal segment, and with a much shorter lateral seta; supraanal seta simple, about 3/4 as long as procercus terminal setae, longer than posterior parapod; anal tubules in two pairs, shorter than posterior parapods; posterior parapod with subapical field of minute spinules and 12–20 dark terminal claws varying in size, some of the shorter claws with 3–5 shorter teeth between apical tooth and claw base.
Pupal morphology ( Fig. 4 View FIGURE 4 ). In relation to the characterizations in Langton & Visser (2003), who had seen 11 exuviae from ZSM, and in Oyewo & Saether (2008; 10 exuviae), the following data are worth noting.
Body size: exuviae length about 4.0– 4.5 mm ( Germany, Landeshafen Wörth; n=10), 3.8–5.0 mm ( Langton & Visser 2003; n=15), 3.3–4.9 mm ( Oyewo & Saether 2008; n=6).
Cephalothorax: cephalic tubercle indistinct or absent, the latter especially on female pupae; Langton & Visser (2003) interpreted such tubercles as present in the form of "shallow humps". Thoracic horn with about 8 branches in non-dichotomous pattern, several proximal branches distinctly wider than distal ones, distal ends of branches occasionally with short, spinose or digitiform projections; basal ring round, relatively small.
Abdomen: tergites II–VI anteriorly with transverse irregular rows of small to medium size points, many of which arise from separate small patches of dark integument (whereas fig. 27 F in Oyewo & Saether 2008 shows the patches lighter and the points darker). Conjunctive IV/V with band of anteriorly directed (not "posteriorly" as in Oyewo & Saether 2008), often hook shaped spines similar in size to those in the segment II hook row and the longer ones in the anterior transverse tergite rows; this conjunctive band 2–3 point rows high, usually medially reduced or briefly interrupted. Pleura V–VI posteriorly with fields of fine shagreen mostly arranged in row groups, especially on VI frequently expressed as comb-like longitudinal rows each including up to a dozen closely set, laterally pointing spinules. Segment VIII with armament of posterolateral corner quite variable, usually including 2–4 loosely set, unequal teeth appearing as a transverse comb, the posterolateral tooth often distinctly bent to posterior ( Fig. 4 View FIGURE 4 ; Langton & Visser 2003: fig. 121-1 c; Oyewo & Saether 2008: fig. 27 D, I). Segment IX of male exuviae about 350–400 µm long (n=6), of female exuviae 250–300 µm (n=7). Anal lobe fringe taeniae relatively few in number ( Langton & Visser 2003: 8–13; n=30) and hardly longer than the lobe itself ( Fig. 4 View FIGURE 4 ).
Adult male morphology. Mostly as diagnosed by Edwards (1929: 376) and described in Oyewo & Saether (2008: 82–85), but with the following corrections and additions (data from syntype = st, except where stated otherwise).
Size: body length c. 4.4 mm ( Oyewo & Saether 2008: "3.63–3.74"), wing length 2.46 mm; length ratio wing/ fore femur 2.67.
Antenna: flagellomeres 1–3 clearly separated, fm2–12 with length/width ratio grading from c. 0.7 (fm2) to c. 1.4 (fm12); AR 1.76 ( Edwards 1929: "about 1.5", Oyewo & Saether 2008: "1.21–1.56"). Frontal tubercles absent. Eye dorsomedial extension with up to 6 facets per diagonal, its medial end subrectangular. Temporal setae 21, medially and laterally in more than 1 row; clypeals 24. Palpomere 2–5 lengths 55, 170, 150, 255 µm; pm3 with 4 subapical sensilla.
Thorax: as described for the female below, except acrostichal setae 18; dorsocentrals 24/ 27 in 1 (anteriorly) to 3 (posteriorly) rows, humeral area bare except for 1/1 rudimentary seta; prealars 7/7; supraalars absent; scutellars 25 ( Oyewo & Saether 2008 recorded "7–8" but their fig. 26 B shows 7 in just one half of the scutellum) in 1–2 (exceptionally 3) rows.
Wing: as described for the female below, except as follows here. For setation, see also Oyewo & Saether (2008): p. 83–84 but not fig. 26 E. Wing with low to moderate anal lobe, width 80 µm just distal of arculus, 155 µm just distal of FCu, 115 µm at fused ends of C and R4+5; VR 1.11. Squama with 16 setae in partly double row.
Legs: fore tibial scale distally rounded, without "triangular" apex as in fig. 26 C in Oyewo & Saether (2008); mid tibia with single spur in posterior part of the fused combs; one hind tibia with 1 spur in anterior part of the fused combs, the other hind tibia with two spurs, i.e. 1 each in either part of the combs; all spur apices more or less curving away from leg axis ( Edwards 1929: "quite straight"). Mid tibiae with 3/3 subapical sensilla chaetica; hind tibiae with 0/0. LR1 1.19 ( Edwards 1929: "about 1.4 for both sexes"; Oyewo & Saether 2008: 1.29–1.45); LR2 0.55; LR3 0.70. Fore leg BV 1.77, SV 1.79; mid leg BV 3.75, SV 3.98 ( Oyewo & Saether 2008: 4.21–4.67); hind leg BV 2.47, SV 2.90. Pulvillus apparently simple, without a narrow more medial branch in addition to the broadly feathered one (see also Edwards 1929: 376, second paragraph).
Hypopygium: tergite IX with 14 dorsomedial setae; laterosternite IX with 3/4 setae. Anal point with distinctly widened apical section curving to ventral. Sternapodeme with transverse section much more massive than the posterolateral arms. Superior volsellae of st and all non-type males examined as in Langton & Pinder (2007) figs 95 A and 226 C, and as in fig. 26 F/H in Oyewo & Saether (2008), in that the volsellar 'base' is evident only as a proximal-medial projection that has very little height relative to the longitudinal extent of the volsella, and that carries all the volsella's setae (3/ 3 in the st); we have not seen "variation" as in Oyewo & Saether (2008) fig. 26 I –K, with a much larger 'base' and with setae arising near or even beyond mid length of the volsella. Inferior volsella without distinct distal swelling or separate apical section; sockets of several distal setae set on short tubercles, including that of the apical posteriorly directed seta, which arises ventrally below the distal-most dorsal recurved setae and is significantly longer than in fig. 26 F, G in Oyewo & Saether (2008). Gonostylus gradually narrowing at both ends, medially without the row of long and fine setae considered as typical of species in Pentapedilum , but distal half with medial to ventral field of well separated setae that are shorter than the width of the gonocoxite, vary in size and include about 15 quite strong rather than fine setae. Fig. 26 F in Oyewo & Saether (2008) does not reflect this adequately; fig. 226 C in Langton & Pinder (2007) is better (but its tubercular apex of the gonostylus is artefactual).
Adult female morphology ( Figs 5 View FIGURE 5 , 6). Based on 1 syntype (st), except where stated otherwise (E. = Edwards 1929; ph = pharate in exuviae).
Size and coloration: body length c. 2.5 mm, wing length 2.48 mm; length ratio wing/fore femur 2.70. Thorax brownish, lighter than in male (E.), especially the ground color and scutellum; abdomen brownish with pale conjunctives (st; Edwards wrote of pale "posterior margins of tergites"); halters yellow; legs brownish, lighter than in male (E.).
Antenna: with 5 flagellomeres, fm1–4 with distinct distal 'necks' relatively longest on fm2–3, shortest on fm1; AR 0.32; fm5 <(fm3+fm4). Frontal tubercles absent (n=2). Eye with dorsomedial extension about as high as wide, with up to 6 facets per diagonal, its medial end subrectangular. Temporal setae 12–14, medially in short double row (n=2); clypeals 30 (ph). Palpomeres 2–4 50, 145, 135 µm long; pm5 c. 1/5 longer than pm4 (ph); pm3 with 4 subapical sensilla.
Thorax: antepronotal lobes not overreached by pronotum, strongly narrowing from broad lateral ends, dorsal ends distinctly separate; lobes bare of setae. Scutal tubercle absent; acrostichal setae 30 in two rows beginning at anterior end of pronotum; dorsocentrals 43/ 45 in irregular rows (1–3, anteriorly exceptionally 5, setae wide) that reach the anterior humeral area, setal sockets set in distinct clear circles; prealars 8/8 of variable size in 1–2 irregular rows; supraalars absent; scutellars 23 in 1–2 (exceptionally 3) rows.
Wing: with faint clouding (most evident in microscope dark field view; s. a. Edwards 1929: p. 376 and pl. XIX, fig. 15) along major veins in distal wing half, in cells around wing tip, RM and FCu, and centrally in an; wing without anal lobe, relatively narrow proximally, widening from 75 µm just distal of arculus to 185 µm just distal of FCu, subapical width at fused ends of C and R4+5 125 µm; veins C, Sc, R, R1, R4+5, RM, M, M1+2, M3+4, Cu and Cu1 clearly demarcated and entirely setose, R2+3 (bare, running close to R1) and An (setose) no more distinct than the two setose wing 'folds' alongside Cu; cells r4+5, m(except near proximal end)-m1+2, m3+4, an-cu1 densely setose. VR 1.18. Squama with 11–14 setae (n=2), the proximal ones in more than 1 row.
Legs: fore tibial scale without spinose projection; mid tibia either with single spur in posterior part of the fused combs (ph) or with two spurs, i.e. 1 in either part of the combs (st); hind tibia either with 1 spur in anterior part of the fused combs (ph) or with 2–3 spurs, i.e. 1 in either part of the combs plus unilaterally a third spur arising subterminally outside the combs (st); all spur apices more or less curving away from leg axis. Mid tibiae distally with 13/16 sensilla chaetica in partly double row; hind tibiae with 0/0. LR1 c. 1.4 (E. for both sexes), c. 1.2 (ph), fore tarsi of st missing; LR2 0.52; LR3 0.67. Mid leg BV 3.64, SV 4.08; hind leg BV 2.61, SV 2.99. Pulvillus simple, broadly feathered, reaching beyond mid length of claw.
Genitalia (n=2); see Figs 5 View FIGURE 5 , 6: sternite VIII posteriorly with pair of separate fields of c. 35 setae each; gonocoxapodeme straight in ventral view, narrowing towards medial end at origin of dorsomesal lobe; coxosternapodeme undulating, with two bends to anterior flanking one bend to posterior. Notum 160 (ph), 185 (st) µm long; rami c. 70 µm, distally curving towards each other. Seminal capsule elongate oval (slightly compressed in both slide mounts), with sclerotized neck, combined length 135 µm; spermathecal ducts indistinctly preserved, possibly S-shaped. Gonapophysis VIII ( Fig. 5 View FIGURE 5 ) with dorsomesal lobe posteriorly broad, squarish or rounded; ventrolateral lobe narrowing from base to tip, distally brush-like, recurved to anteromedial; apodeme lobe, if present, reduced to hardly more than the apodeme (similar to Oyewo & Saether 2008: fig. 46C). Labia indistinctly preserved, without noticeable trichia. Tergite IX covered with 40–45 setae, gonocoxite IX with 5 setae (n=2). Segment X with 8–11 lateral setae (n=2); postgenital plate a broad, rounded triangle (ph). Cercus 90–110 µm long (n=2), 95 µm high (st).
Ecology and distribution. In the spring and summer of 2013 the present junior author found larvae of N. nubens living in algae and fine sediments on coarse boulders along the W shore of Landeshafen Wörth, an artificial oxbow on the W bank of the river Rhine, in an area reconstructed as a harbor for container ships. Larval densities were <100 individuals/m ² in the spring (2.iv.2013), but>1000 individuals/m ² in summer (19.vii.2013).
In addition to the collecting sites of larvae examined for the present study (see the Material examined section above), further records are known from GERMANY, Rheinland-Pfalz, Eifel region: Meerfelder Maar, at 9 cm depth in a sediment core, and Weinfelder Maar, rare in two sediment profiles ( Hofmann 1993); FRANCE, dépt. Haute-Loire, Lac du Bouchet, subfossil to Recent sediments ( Hofmann 1991); northeastern SPAIN, lower rio Ebro; c. 150 larvae, most of them found in places without measurable current, but one sample of 32 larvae taken at a flow rate of 65 cm /sec ( Cid Puey 2010); and from northeastern ALGERIA, Guelma, Oued Seybouse drainage, Oued El Nil near confluence to Oued Cherf; temporary stream, unpolluted site with very coarse gravel substrate, water temperature <10 °C, and current speed> 20 cm /sec ( Chaib et al. 2013).
Hofmann (1991) wrote that in Lac du Bouchet, " Chironomini type A" had been found from the sediment surface down to a depth of 1.20 m—the local boundary between Holocene and older sediments—and that it "should therefore still be present in the lake." Other taxa in the same profile zone clearly indicated a process of considerable eutrophication from the early Holocene to the present, but the numbers of larvae recorded for N. nubens in various parts of the sediment profile were not considered as reflecting that environmental change. Hofmann (1993) did not specify the Weinfelder Maar sediment horizon or zone in which he had found " Chironomini A", but the lake has kept an oligotrophic state until today. Meerfelder Maar, on the other hand, was highly eutrophicated by the end of the 19th century already; thus, a larva found at no more than 9 cm sediment depth may be surmised not to have lived in oligotrophic conditions. The two samples from the German upper Rhine system certainly come from eutrophic waters. No comparable details are known from the remaining collection sites.
These collecting sites of larvae agree well with the range of Nubensia nubens based on samples of other life stages ( Saether & Spies 2013). Therefore, the verified distribution now extends from the British Isles and central Europe to the western Mediterranean, including northern Africa, with possible extensions to Turkey and central Asia (based on unassociated adult males). However, the record from Norway in Oyewo & Saether (2008) is doubtful (see the morphology section in the discussion below).

| ZSM |
Bavarian State Collection of Zoology |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Nubensia nubens (Edwards)
| Spies, Martin & Dettinger-Klemm, Andreas 2015 |
Pentapedilum (Pentapedilum) nubens
| Edwards 1929: 376 |