
VATELLINI, Sharp, 1880
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2005.00180.x |
|
persistent identifier |
https://treatment.plazi.org/id/2630926F-C17E-214C-6DDB-FA80833A3285 |
|
treatment provided by |
Diego |
|
scientific name |
VATELLINI |
| status |
|
MORPHOLOGY OF VATELLINI View in CoL
The members of this tribe exhibit morphological apomorphies in adults that are unusual among diving beetles and require some explanation and/or novel terminology. The following account is given to provide a basis for the formal descriptions of taxa, to point out and describe characters of particular taxonomic importance, and to provide some context for future comparative morphological studies that include members of the tribe. General aspects of vatelline morphology in the following treatment are based especially on cleared and disarticulated specimens of V. grandis and D. lentus , although other species were also examined to determine generality of the characters.
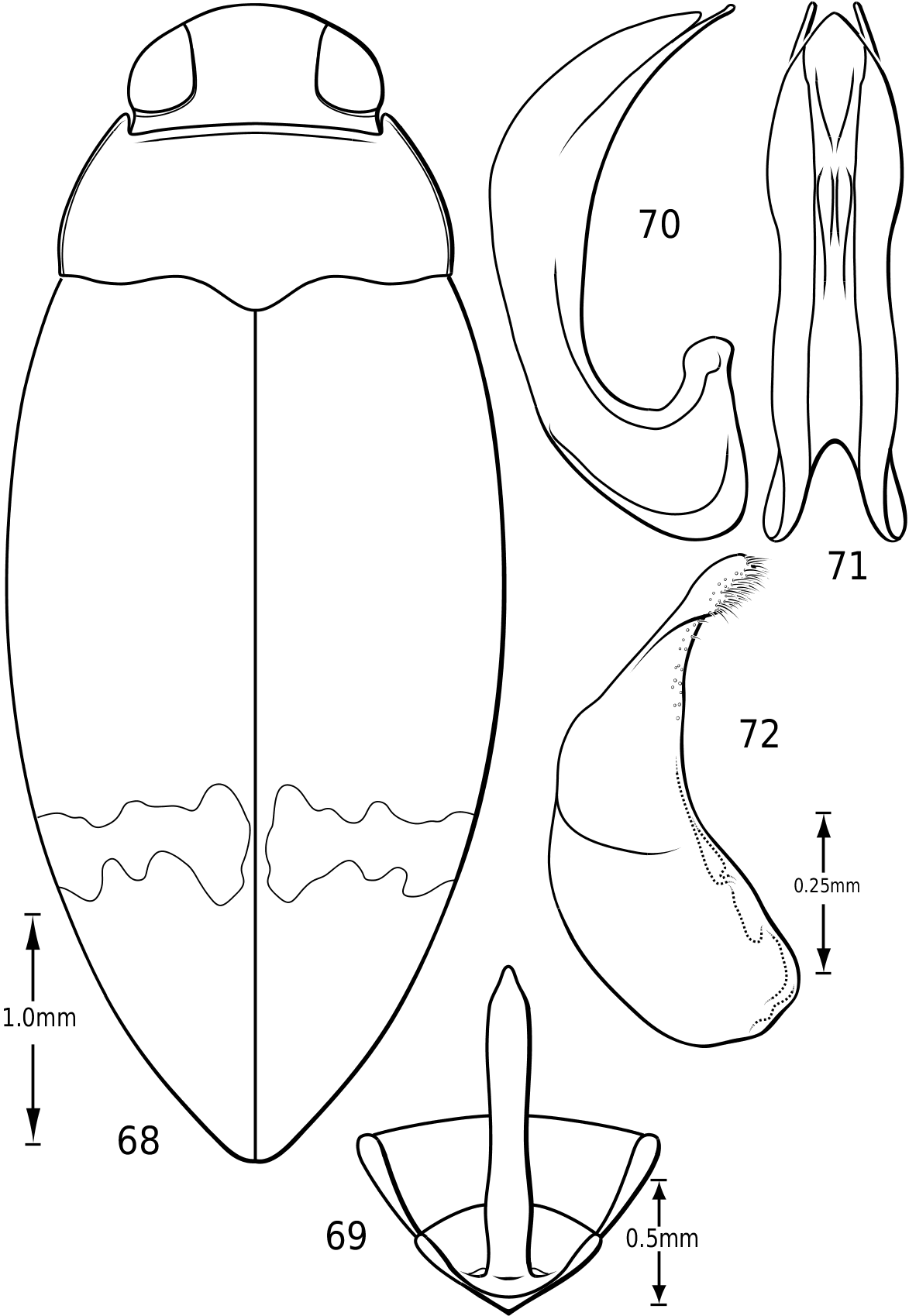
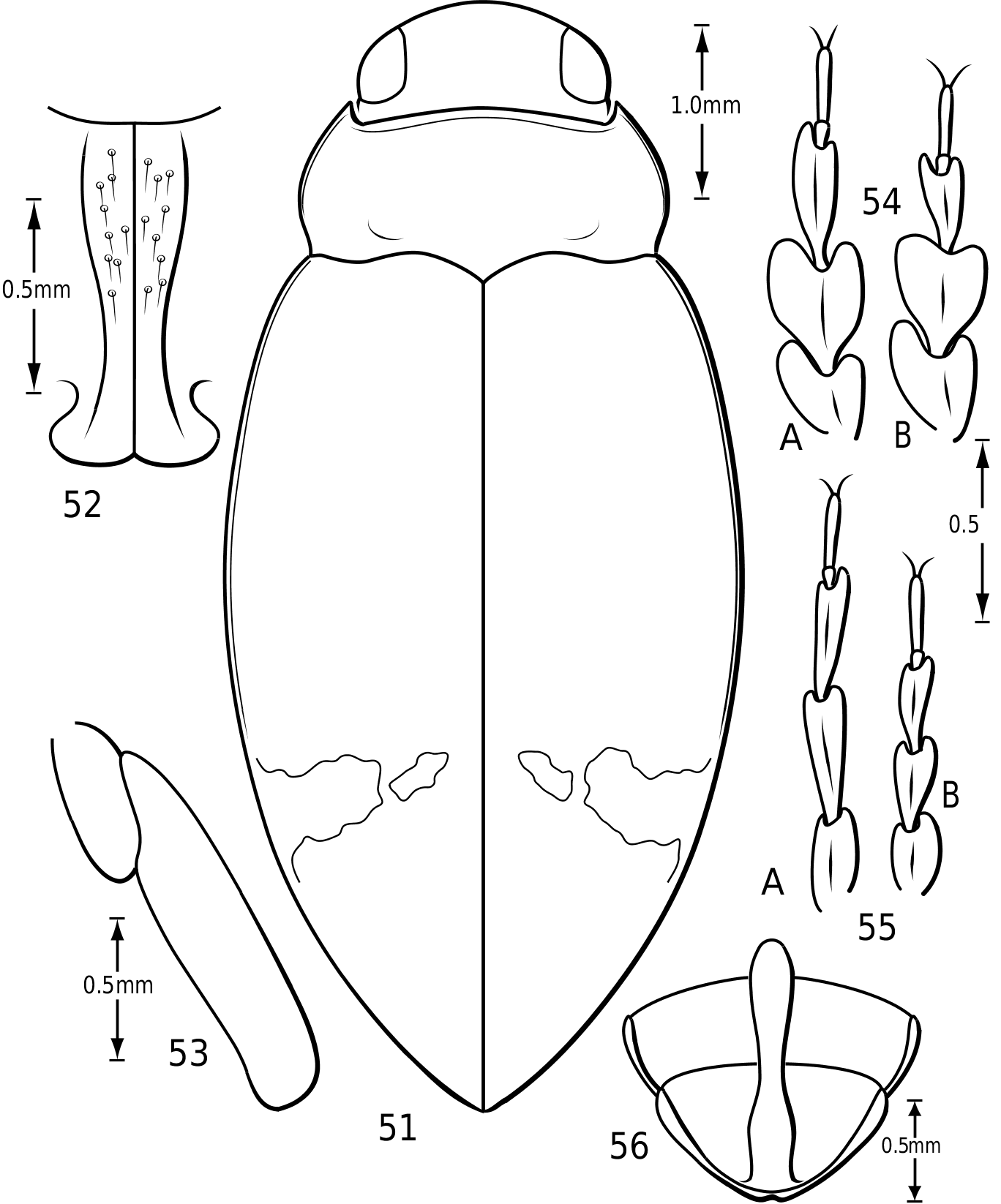
Body form: Members of Vatellus are elongate and have the lateral outline somewhat to strongly discontinuous between the elytron and pronotum with the pronotum often cordate (e.g. Fig. 1). Members of Derovatellus are continuous or somewhat discontinuous along the margin from the pronotum to elytron in dorsal aspect (e.g. Fig. 68 View Figures 68–72 ) or have the lateral margin of the pronotum strongly curved and the lateral outline strongly discontinuous (e.g. Fig. 51 View Figures 51–56 ). Some members of Vatellus are distinctly flattened dorsally and relatively strongly dorsoventrally compressed.
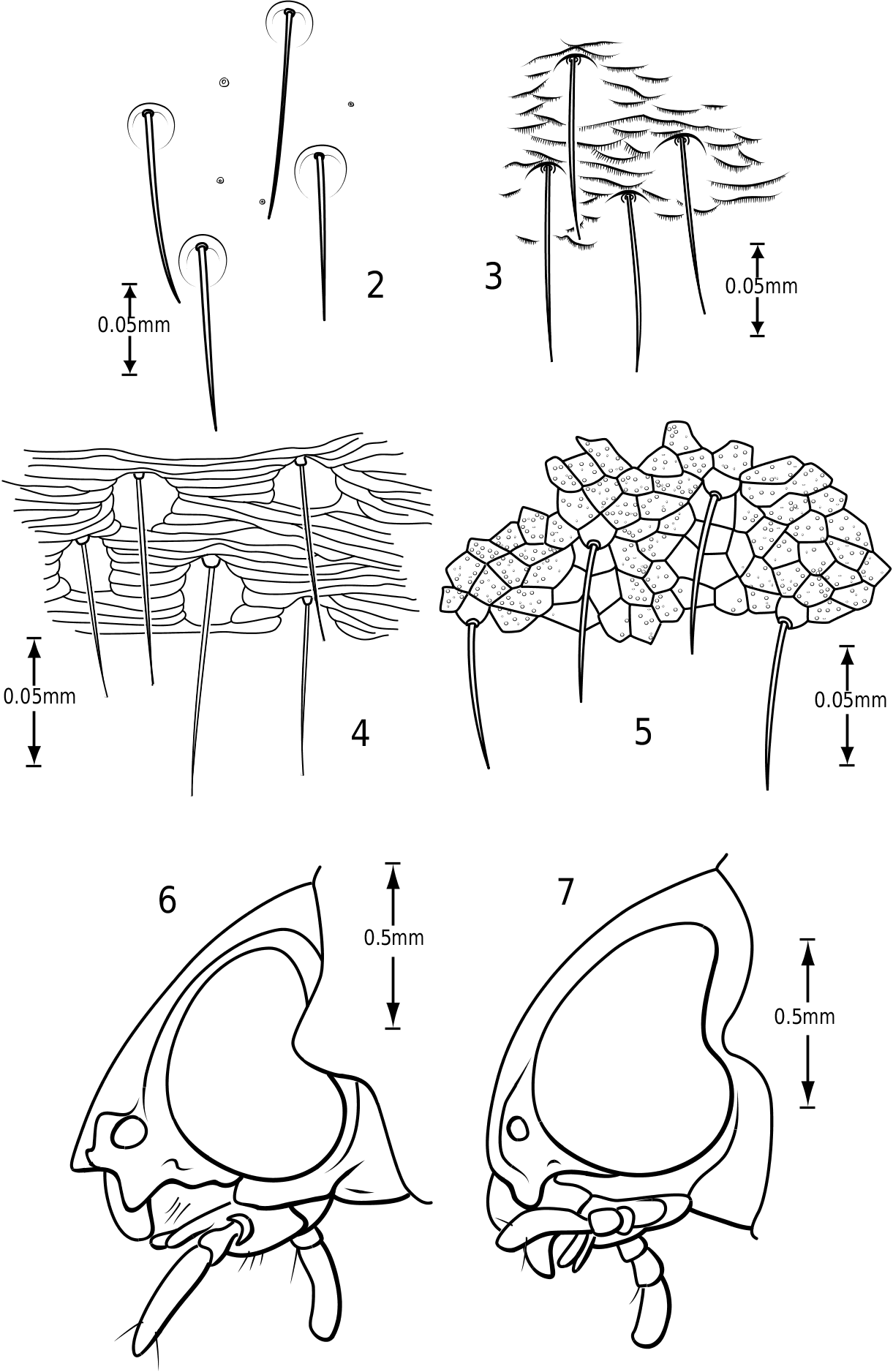
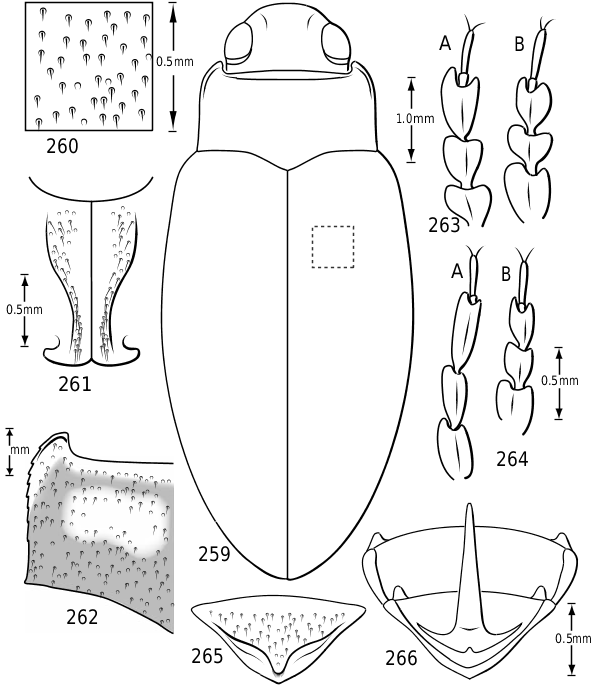
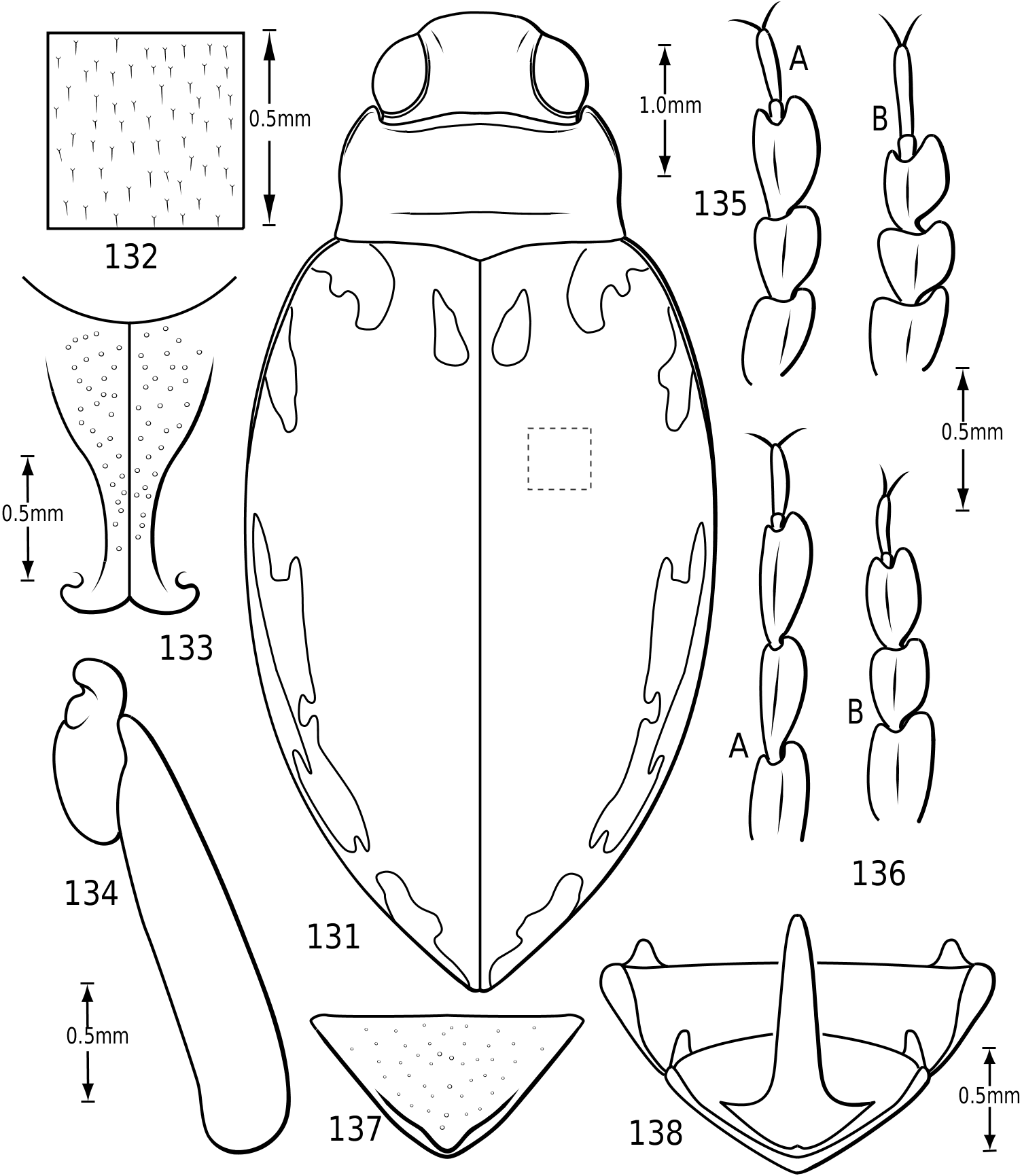
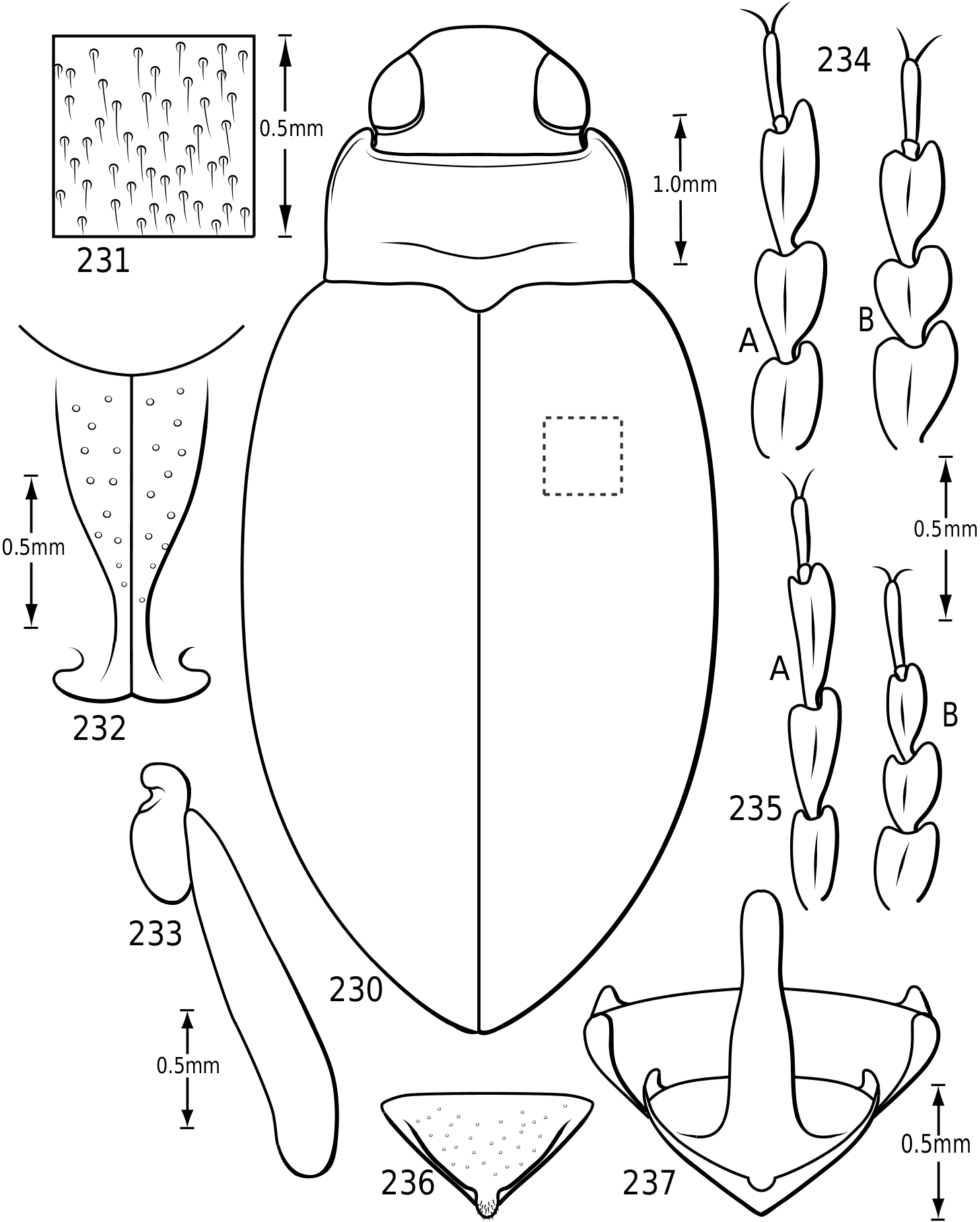
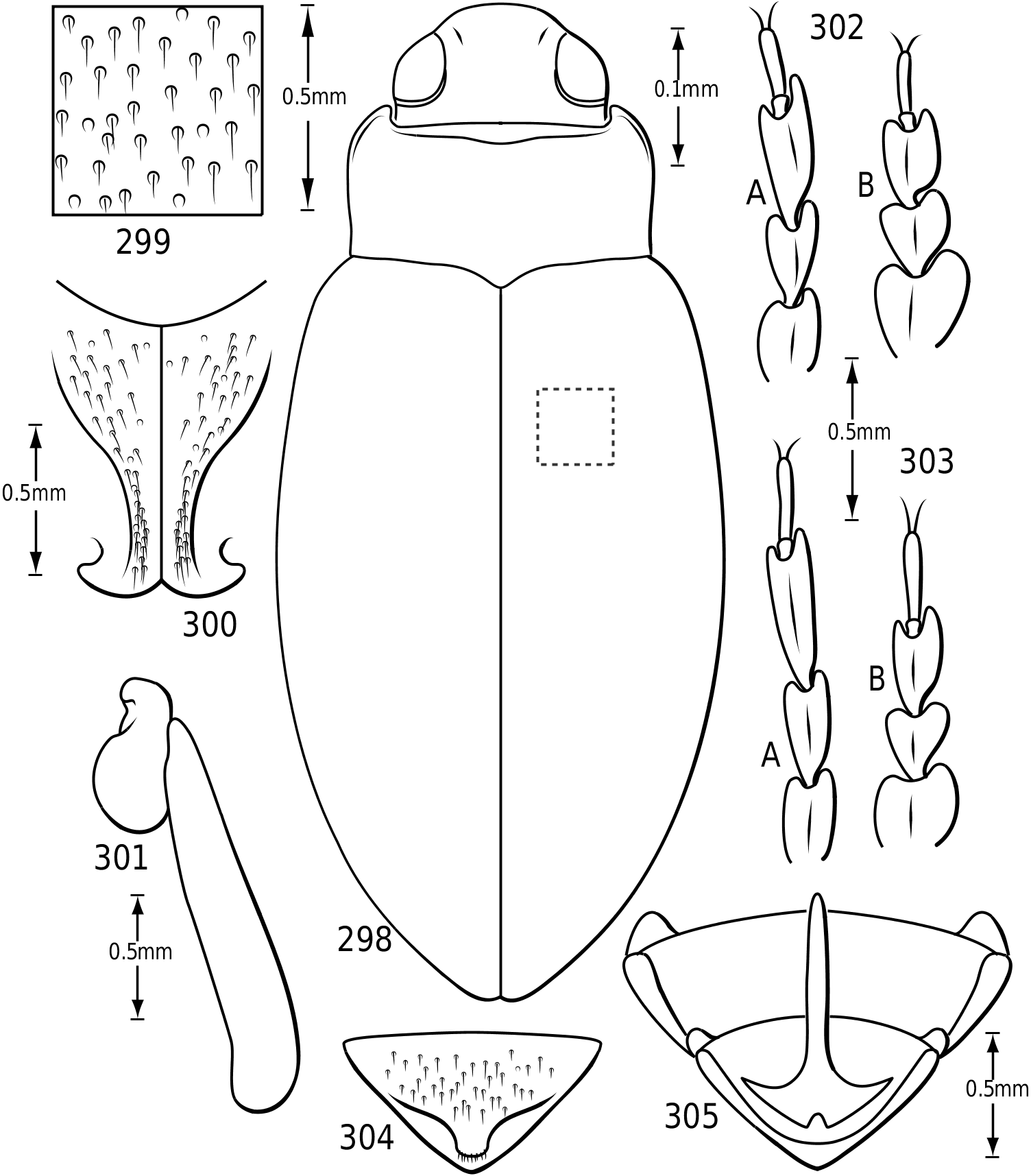
Surface sculpture: Vatellines, like many Hydroporinae (e.g. see Wolfe & Zimmerman, 1984), have a variety of surface features in the form of punctures, setae and microreticulations that vary between species. All species have scattered setae on the pronotum and elytra. The setae are set in prominent pores (some Vatellus , e.g. Fig. 2 View Figures 2–7 ) or they arise from small depressions ( Derovatellus and some Vatellus , e.g. Figs 3, 4 View Figures 2–7 ) or flat areas (some Vatellus , e.g. Fig. 5 View Figures 2–7 ). The surface between the punctures may be relatively smooth and shiny (many Vatellus , e.g. Fig. 2 View Figures 2–7 ), marked with transverse, impressed lines forming transversely orientated microcells ( Derovatellus , e.g. Fig. 4 View Figures 2–7 ) or marked with short, curved striae bearing very short, spinous, posteriorly directed setae (some Vatellus , e.g. Fig. 3 View Figures 2–7 ). Several Vatellus have irregular cells with small microcavities or irregularities on the surface of most cells ( Fig. 5 View Figures 2–7 ). The surfaces of the metasternum, metacoxae and abdominal sterna exhibit similar patterns of surface sculpturing. In some species of Vatellus a portion of the pronotum and, in some cases, the elytron (especially apically) are marked with a large area of fine shagrination comprising extremely fine microtubercles. When fully developed this shagrination forms a pattern with smooth, shiny, impunctate transverse areas lacking shagrination on each side of the midline of the pronotum ( Fig. 262 View Figures 259–266 ).
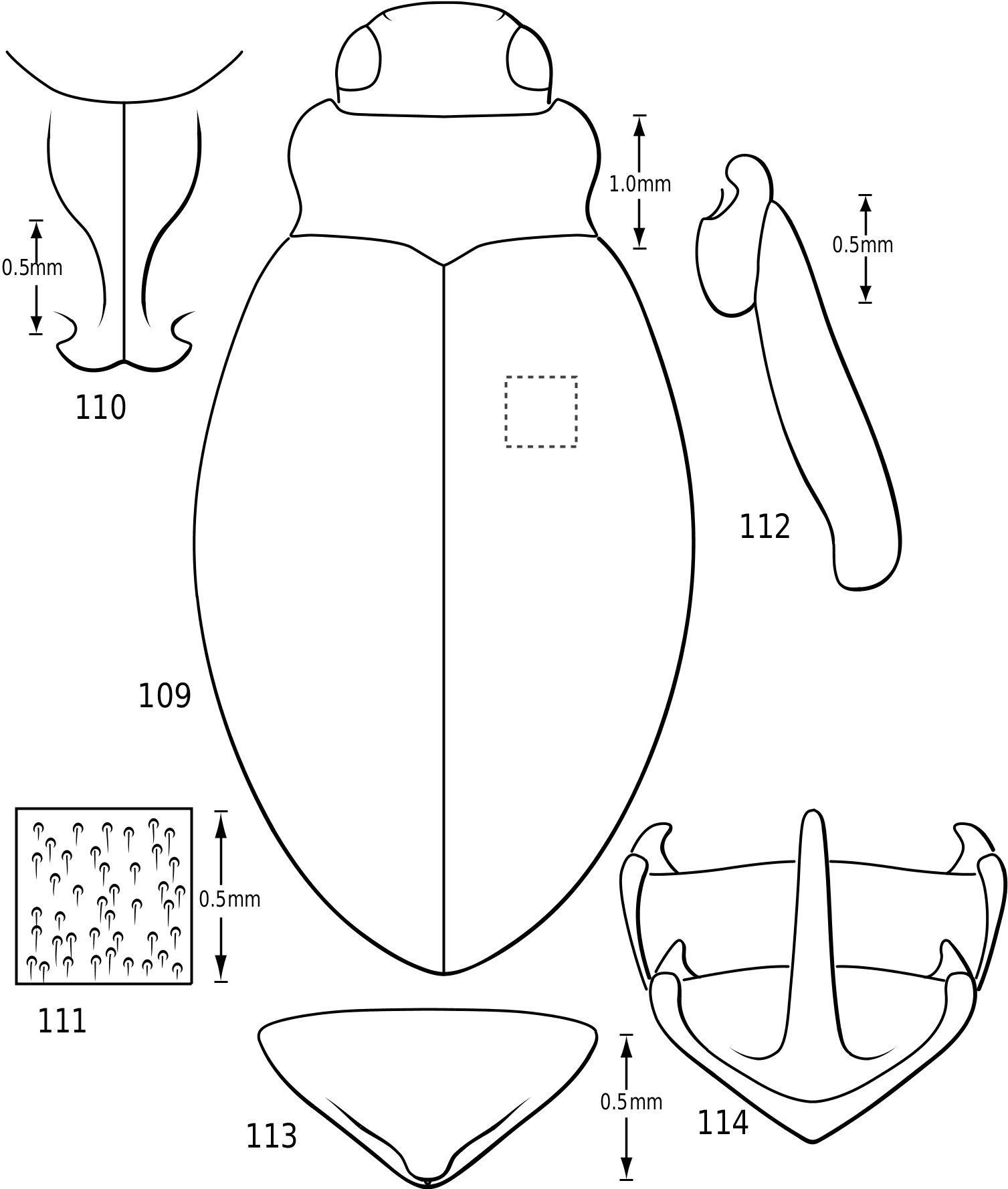
Cranium: The head in vatellines is short, rounded and robust. The eyes are located anterolaterally and are very large and protuberant, especially in some species of Vatellus (e.g. Fig. 131 View Figures 131–138 ). The eyes are slightly emarginate along anterior and posterior margins. A paraocular carina extends from the anterior margin of the eye around the eye dorsally, posteriorly and ventrally. There is a distinct cavity posterad of the eye laterally that appears to receive the anterolateral angles of the pronotum. The frontoclypeal suture is effaced medially and indistinct laterally. The anterior clypeal margin in some Vatellus protrudes anteriorly in a distinct, angulate rim ( Fig. 6 View Figures 2–7 ), whereas in other Vatellus and Derovatellus the anterior margin of the clypeus is evenly rounded ( Fig. 7 View Figures 2–7 ). The tentorial arms are very slender and there are no tentorial bridges. The occipital foramen is very large, about one-half the width of the posterior surface of the head.
Antennae: Vatelline antennae are not modified from a form typical of most Hydroporinae. The scape and pedicel are longer and broader than flagellar antennomeres. The antennae are moderately long and antennomeres vary in relative size between species, but not strongly so.
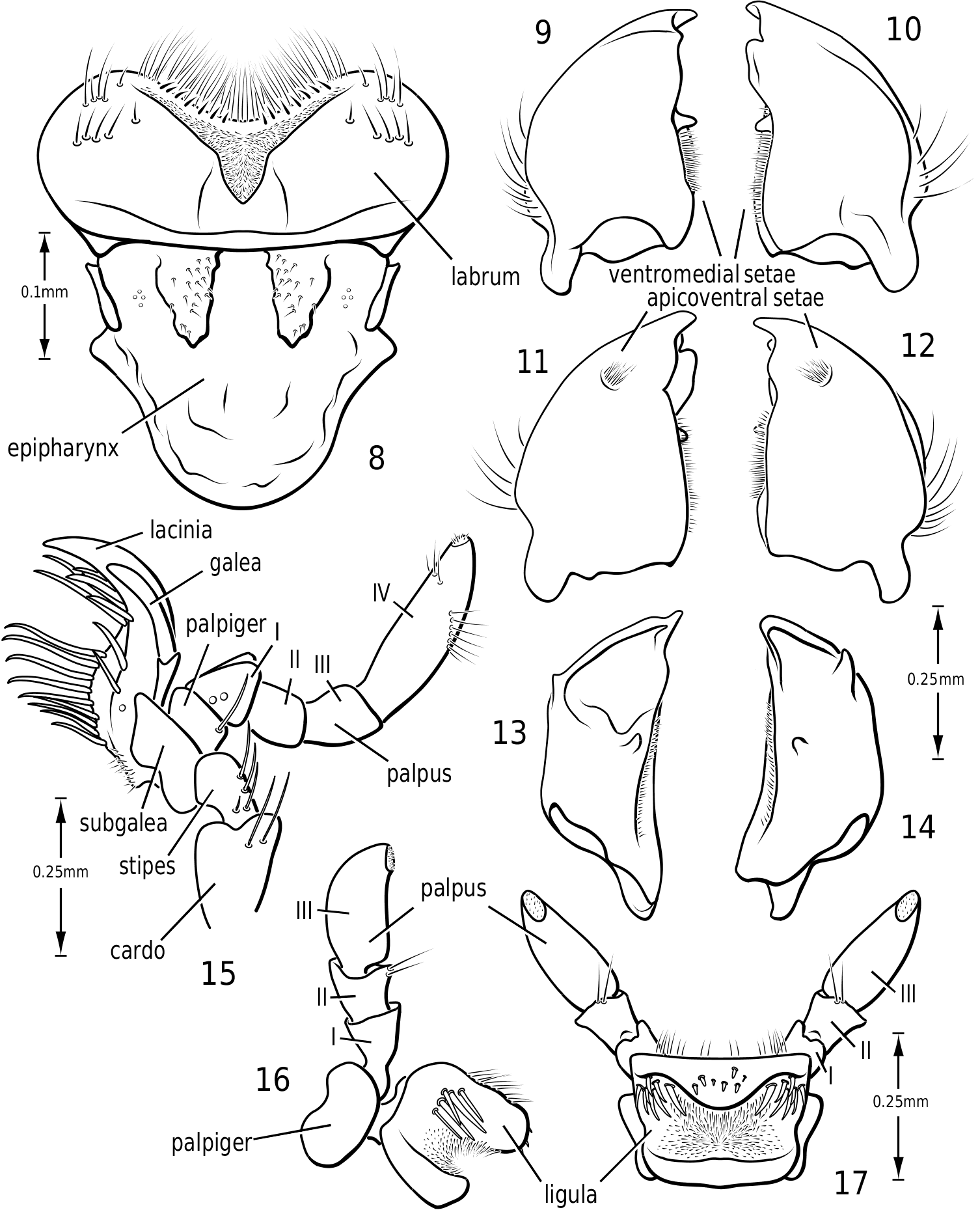
Labrum and epipharynx ( Fig. 8 View Figures 8–17 ): The labrum is broad and apically broadly emarginate medially. The medial margin bears a dense fringe of long fine setae and a series of short stout setae. Laterally on the aboral surface are numerous long, stout setae. Medially on the aboral surface is a broad V-shaped area of dense, short setae. The epipharynx is bordered laterally on each side by a short, stout sclerite. Medially there are two thickened membranous regions bearing short sensory setae.
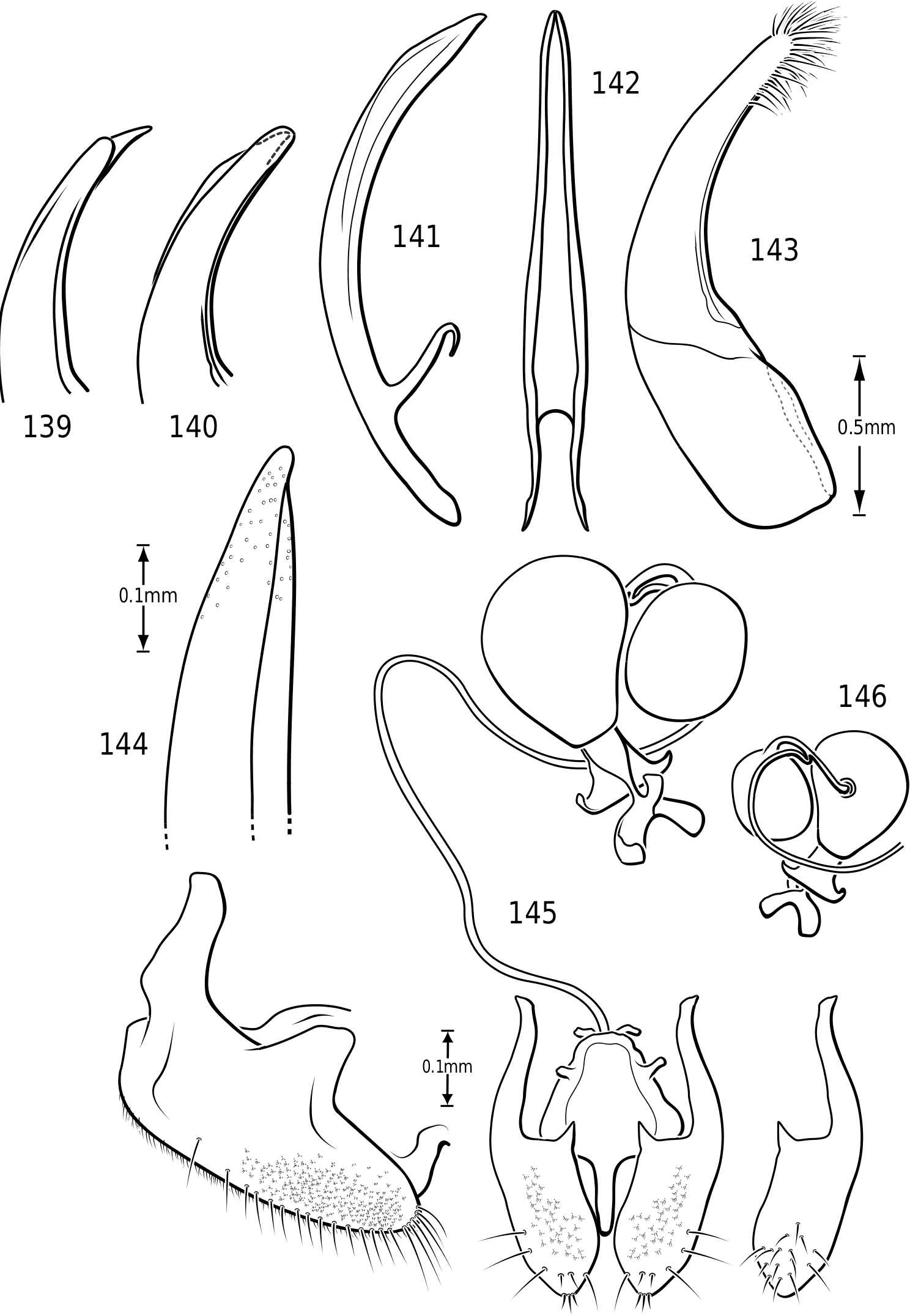
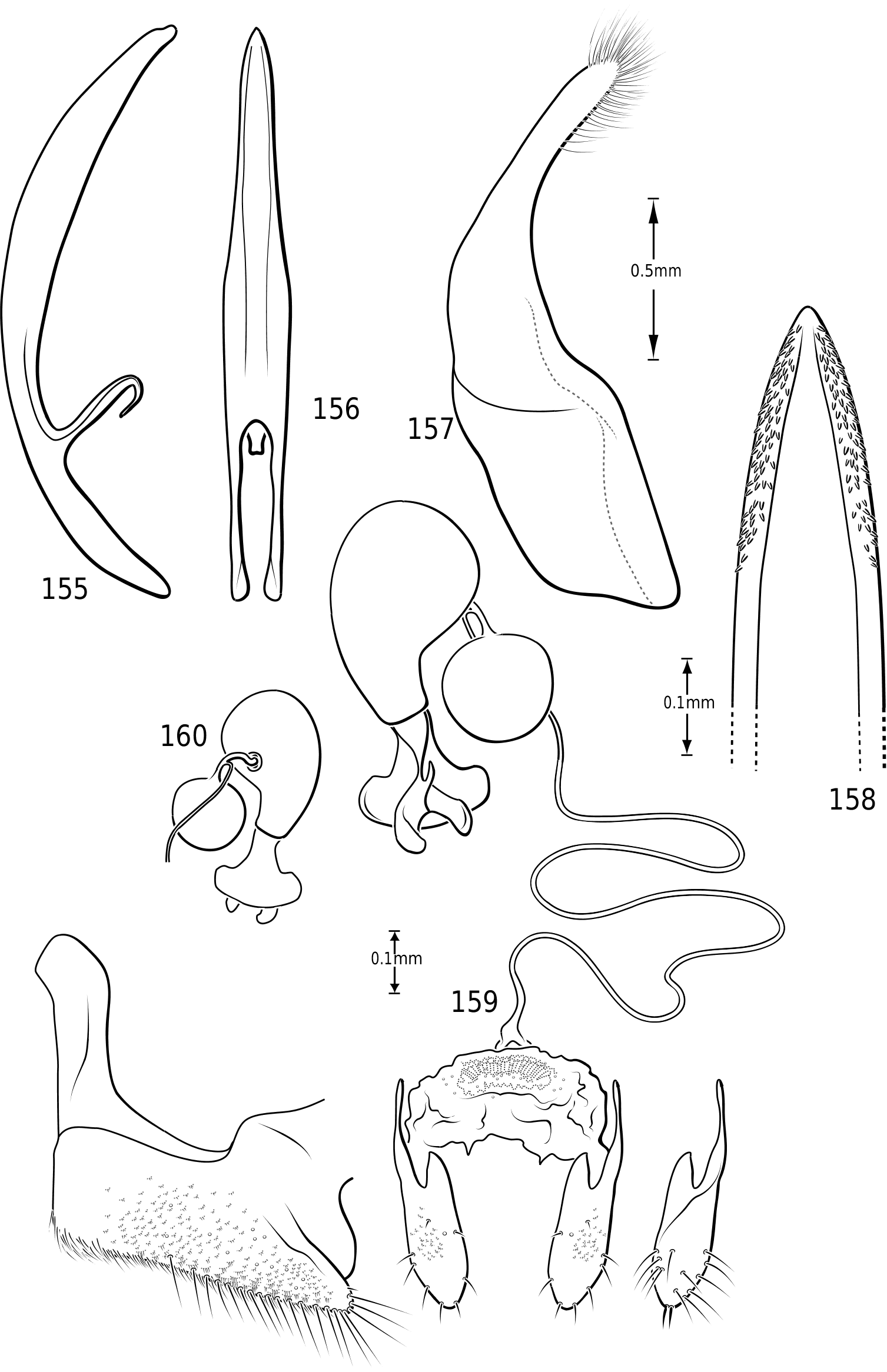
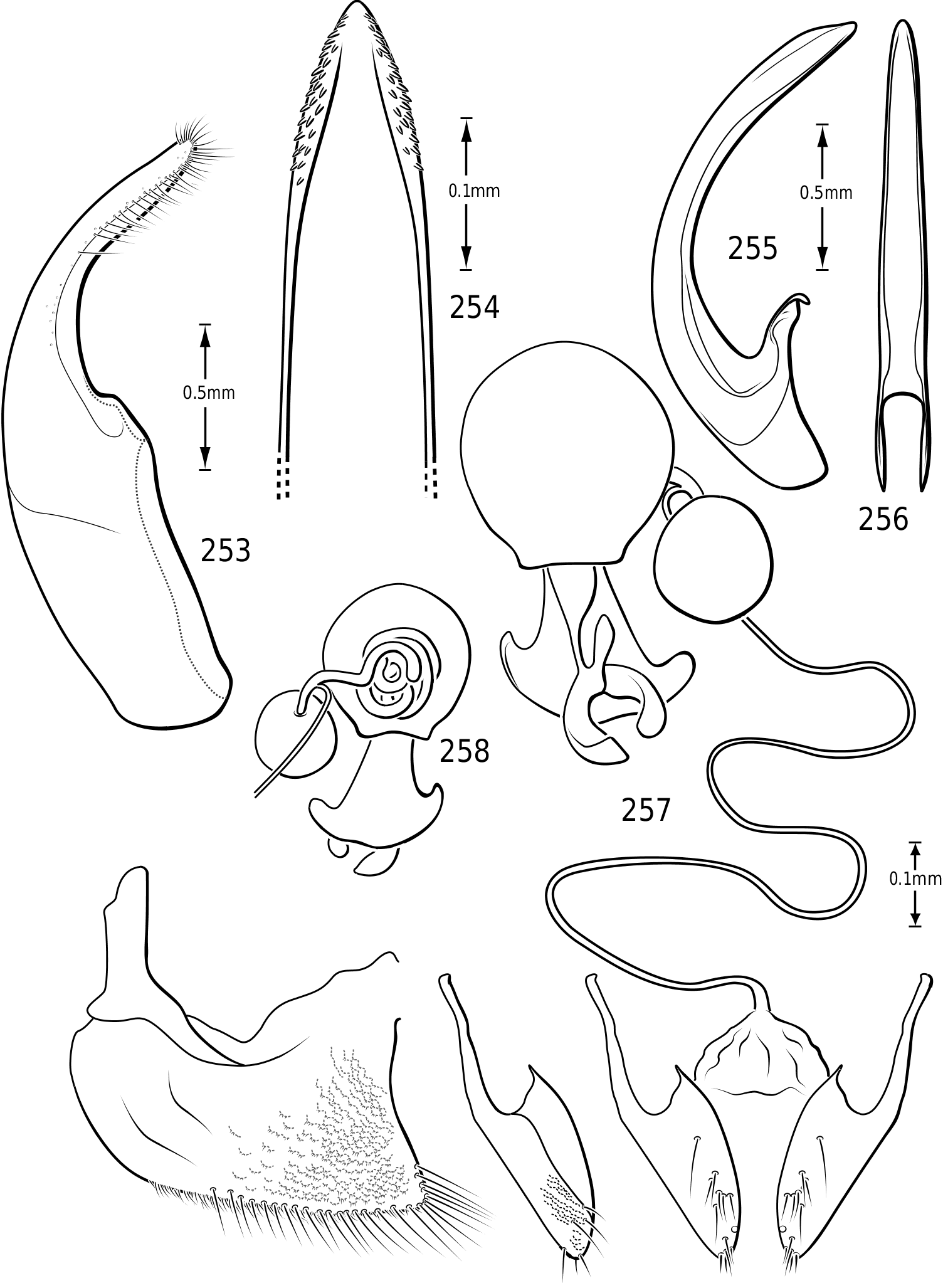
Mandibles ( Figs 9–14 View Figures 8–17 ): The mandibles in Vatellini are asymmetrical. Each is broad and broadly curved laterally. The apices of each are dorsoventrally broad. The left mandible is strongly excavated apicomedially ( Fig. 13 View Figures 8–17 ). The pointed apex of the right mandible fits into this excavation. The medial margin of each mandible bears a long series of short setae. Medially, each mandible bears a small tooth-like retinaculum. Many Hydroporinae lack mandibular setae, a proposed synapomorphy of the subfamily ( Wolfe, 1985, 1988). However, all Vatellini examined possess a medial fringe of setae and a medial patch of setae on the ventral margin ( Figs 11, 12 View Figures 8–17 ). Vatellus grandis (at least) also possesses long lateral setae ( Figs 9–14 View Figures 8–17 ).
Maxilla ( Fig. 15 View Figures 8–17 ): The cardo is stout and apically expanded. The stipes is small and narrow medially. The subgalea is elongate and irregular in shape. The palpus comprises four palpomeres and a basal palpiger. The palpiger and palpomeres I–III are relatively short and stout. Palpomere IV is longer than the basal three and bears an oblique, interrupted ring of sensory setae submedially and a small apical patch of sensillae. The galea consists of two long, slender segments. The lacinia is broad, strongly sinuate and sharply pointed apically. Its medial margin bears numerous long, stout setae.
Labium ( Figs 16, 17 View Figures 8–17 ): The mentum is very broad and flattened with large anterolateral lobes, similar to most Dytiscidae . The ligula is stout and broad with a dorsal transverse excavation which bears stout setae on its lateral surface and a large field of dense, fine setae medially. The anterior surface bears a few long setae. The palpiger is broad and stout. The palpus consists of three palpomeres. The second bears a few apicodorsal setae. The apical palpomere bears a small field of sensillae apically.
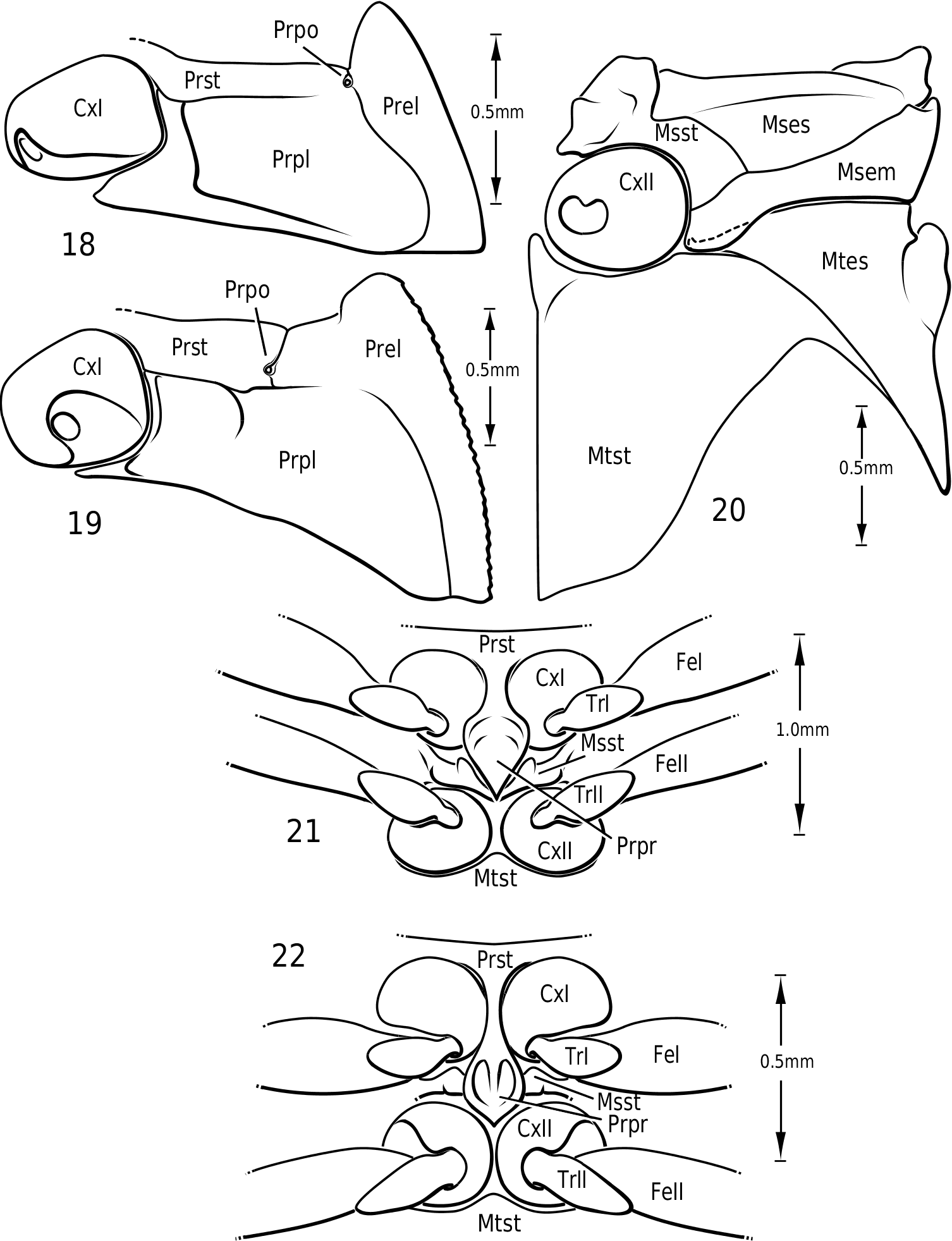
Prothorax: The pronotum varies from subtrapezoidal to conspicuously cordate. In Derovatellus the lateral margins are evenly curved and the pronotum is widest medially (e.g. Fig. 51 View Figures 51–56 ) or near the posterior angles (e.g. Fig. 68 View Figures 68–72 ). In Vatellus the lateral margins are somewhat ( Fig. 259 View Figures 259–266 ) to extremely ( Fig. 109 View Figures 109–114 ) sinuate and are broadest anterad of the middle. The lateral margins are distinctly dentate ( Figs 19 View Figures 18–22 , 262 View Figures 259–266 ). In some species of Vatellus there is a transverse crease or depression in the surface of the pronotum slightly posterad of the middle (e.g. Fig. 230 View Figures 230–237 ). The prosternal pore ( Wolfe, 1985, 1988) in most Hydroporinae is located at or near the anterolateral apex of the prosternum (where the anterolateral margin of the prosternum meets the pronotal epipleuron) ( Fig. 18 View Figures 18–22 ). In vatellines the anterolateral margin of the prosternum is shifted medially and the prosternal pore is shifted posteromedially ( Fig. 19 View Figures 18–22 ). The pore bears a conspicuous lobe ( Fig. 19 View Figures 18–22 ).
In most Dytiscidae , the prosternal process extends between the pro- and mesocoxae where its apex contacts the anterior margin of the metasternum. In a few taxa the prosternal process is relatively short but still more-or-less attains the metasternum (e.g. the Ilybius opacus -group of species), whereas in others the process does not extend between the mesocoxae and the mesocoxae are contiguous. This last group includes several phylogenetically disparate groups like Stictotarsus minipi Larson , the Bidessini genus Tyndallhydrus Sharp , some species of Hygrotus (Coelambus) Thomson , and all members of Vatellini . In vatellines the prosternal process extends ventrally between the procoxae and is curved dorsally posterad of the procoxae ( Figs 21, 22 View Figures 18–22 ). The mesocoxae are contiguous. The apex of the prosternal process is strongly flexed dorsad. The apex may be rounded or flattened and truncate to sharply pointed.
Mesothorax: Sharp (1882b) differentiated between Derovatellus and Macrovatellus based on the relative visibility of the mesosternum on each side of the prosternal process. However, I have not found this to be a useful character. In each group it is visible to approximately the same degree. The mesothorax is narrow and relatively undifferentiated.
Metathorax: As recognized most notably by Sharp (1882b), in extant vatellines the metepisternum does not appear to extend to the mesocoxal cavity, instead it is separated from it by the mesepimeron. This is a condition putatively shared in Hydradephaga with Gyrinidae , Hygrobiidae , Noteridae and Laccophilini. These other taxa differ from Vatellini in having the mesepimeron separated both externally and internally from the mesocoxal cavity. In vatellines the metepisternum extends to the cavity, but the posteromedial angle of the mesepimeron extends ventrally as a lobe over the medial end of the metepisternum ( Fig. 20 View Figures 18–22 ). The metasternum is medially broadly convex. In some species of Vatellus the anteromedial process is prominently swollen ventrad ( Fig. 20 View Figures 18–22 ). In Calicovatellus the metepisternum extends to the mesocoxal cavity externally.
Elytron: The elytron is relatively undifferentiated in vatellines. Dorsally there are no conspicuous striae, carinae or plicae. Occasionally very slightly raised longitudinal lines can be discerned on some species, particularly V. mexicanus and closely related species. Ventrally the apicolateral lobe (elytral ligula) is singular and simple, though it may be quite large in some species, especially in some Vatellus . The lateral carina is low and relatively short with no medial lobe. There are elongate friction pads along the lateral and medial margins comprising fields of minute tubercles or spines.
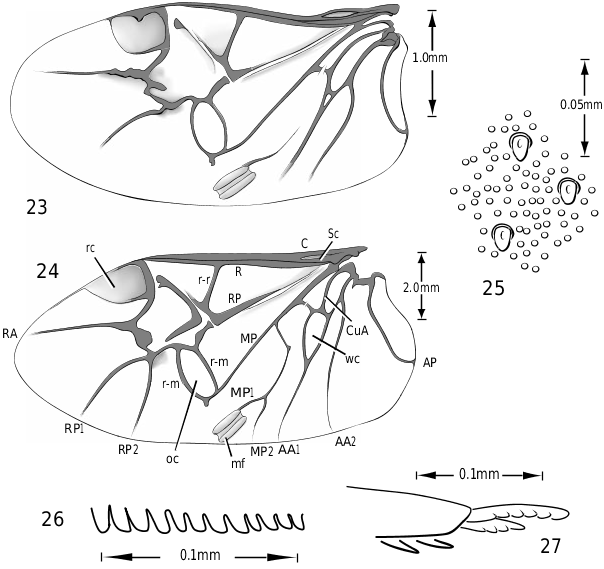
Metathoracic wing ( Figs 23, 24 View Figures 23–27 ): The flight wings in Vatellini are similar in most respects to other hydroporines. The wings are short, broad and strongly rounded apically. The oblongum cell is transversely oval. The longitudinal triangular space between the radius and radius posterior is very broad. The radial cell is broad and strongly rounded apically. The submedial binding patch (this term based on Lawrence & Britton, 1994) is prominent and consists of two elongate, robust, oblique parallel parts and a pigmented area between them. In Vatellus the wedge cell is relatively large and elongate ( Fig. 24 View Figures 23–27 ) whereas in Derovatellus it is very small ( Fig. 23 View Figures 23–27 ), a trend similar in other small Hydroporinae. Derovatellus has reduced venation ( Fig. 23 View Figures 23–27 ), also similar to other small Hydroporinae. In this genus, the media posterior veins (MP) and radius posterior vein 2 (RP2) are weakly visible to absent ( Fig. 23 View Figures 23–27 ). In Vatellus the RP2 is present and branches from the RP1 distad of the oblongum cell ( Fig. 24 View Figures 23–27 ). In Dytiscidae except Hydroporinae and Laccophilini the RP2 arises from the margin of the oblongum cell ( Goodliffe, 1939; Balfour-Browne, 1944; Wolfe, 1985; Wolfe, 1988).
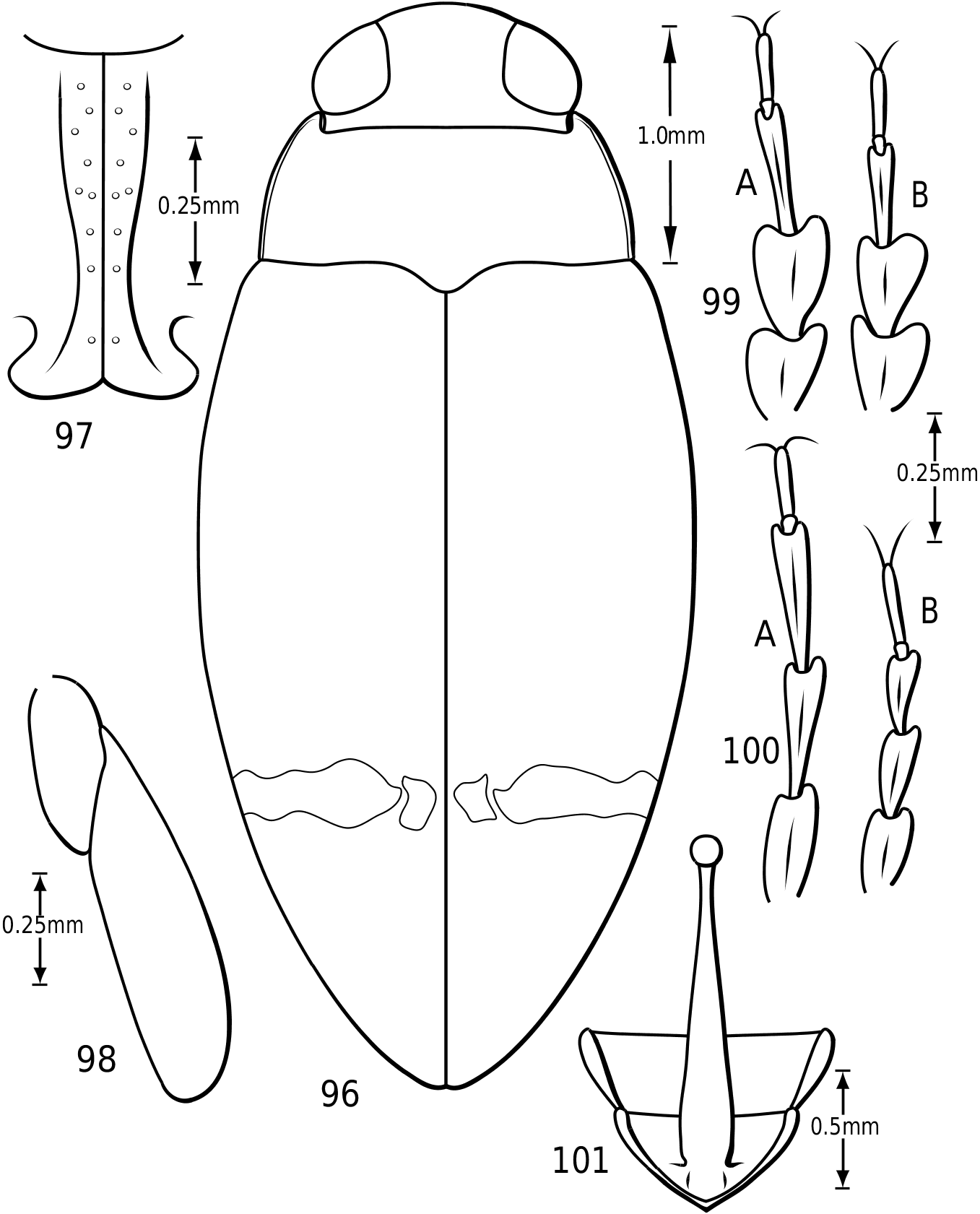
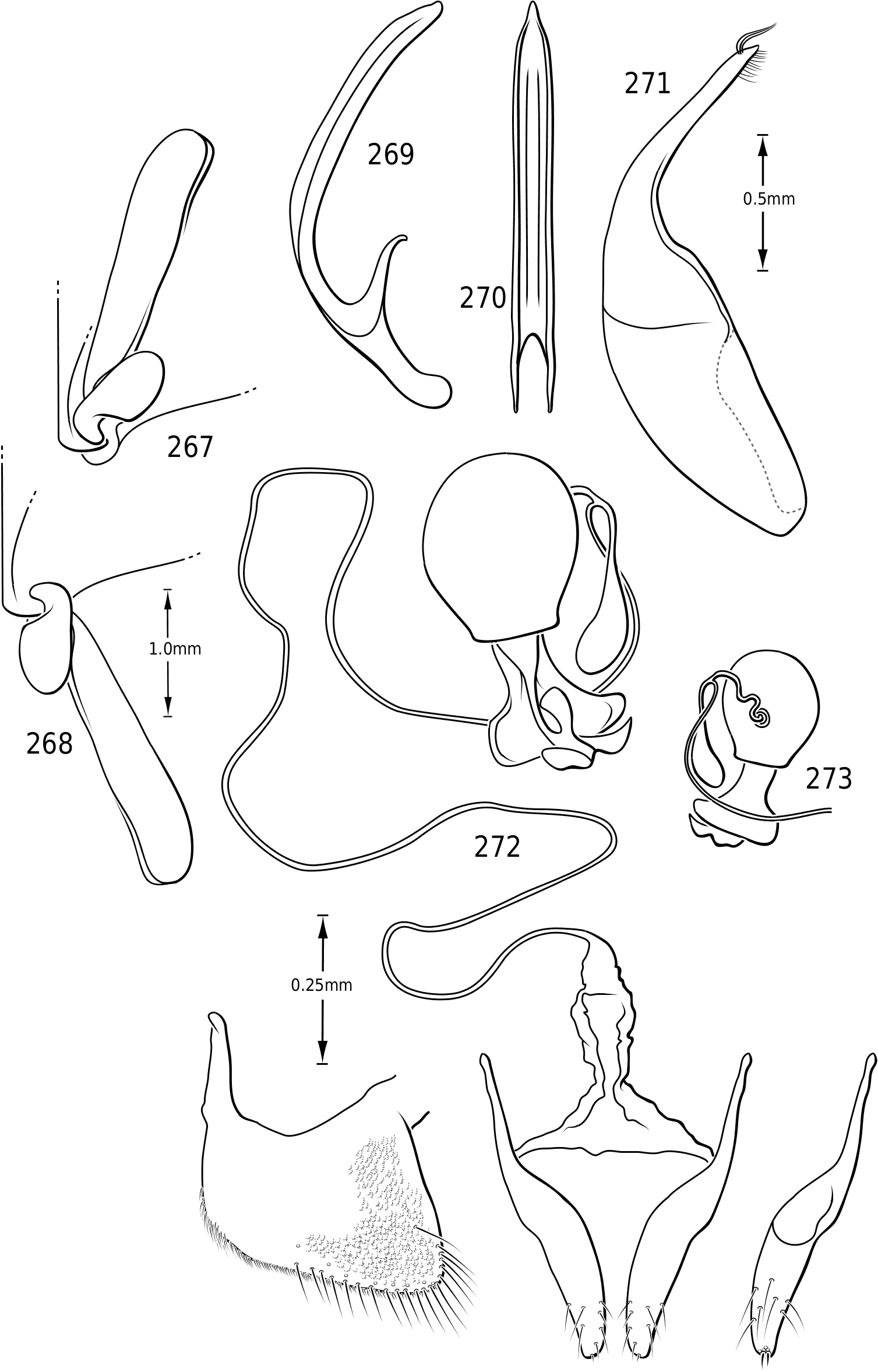
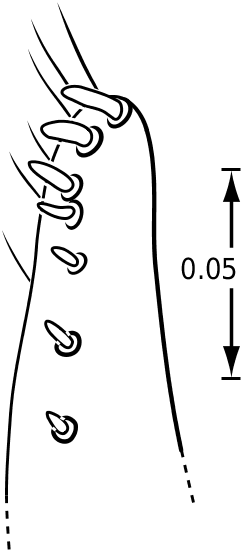
Legs: The legs of vatellines are unusually long. Pro- and mesotarsomeres I–III on both females and males bear a ventral field of adhesive setae ( Fig. 25 View Figures 23–27 ). The apices of the setae are slightly ovoid ( Fig. 25 View Figures 23–27 ) and often dimorphic with many small setae and fewer large setae interspersed ( Fig. 25 View Figures 23–27 ). These tarsomeres are often more strongly expanded in males than in females, though only slightly more so in some species of Vatellus . All tarsi and tibiae bear long natatory setae along the dorsal and ventral margins. Males of many species, especially in Vatellus , have a large cluster of fine, long setae on the apicoventral surface of the mesotrochanter and the base of the mesofemur. The metacoxae are strongly anteriorly expanded and are relatively long. The metacoxal lines are closely approximated in Derovatellus (e.g. Fig. 97 View Figures 96–101 ). In many species of Vatellus the metacoxal lines are closely approximated posteriorly and are strongly divergent anteriorly (e.g. Fig. 300 View Figures 298–305 ). The lateral portion of the metacoxa is variously punctate from nearly impunctate to very coarsely punctate and in some cases there is a distinctive microsculpture similar to that found on the elytra (see above under Surface sculpture). The anteroventral margins of the metatibia and metatarsomeres of those species examined have a series of very fine dentitions ( Fig. 26 View Figures 23–27 ). The metatarsal claws are unequal in length with the posterior claw slightly longer. The metatarsal claws are denticulate with rounded teeth ( Fig. 27 View Figures 23–27 ). The metatrochanter in Vatellus is often strongly offset (e.g. Fig. 301 View Figures 298–305 ). It bears a prominent basal, circular lobe that extends medially under the surface of the metacoxa when the leg is flexed anteriorly and a distinct, broad emargination in the ventral (posterior) surface (e.g. Figs 267, 268 View Figures 267–273 ). The metatrochanter in Derovatellus is not unusually modified ( Fig. 98 View Figures 96–101 ).
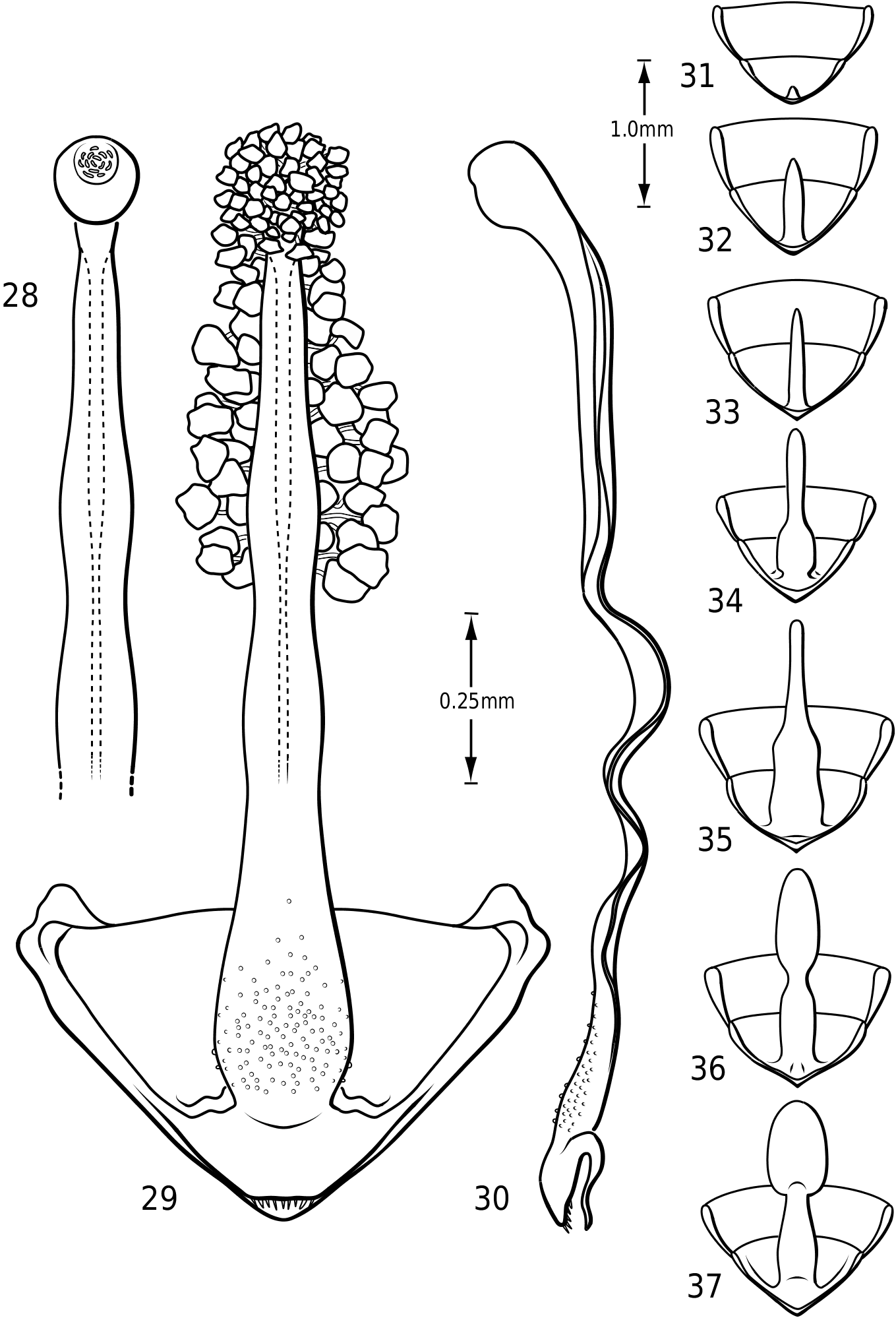
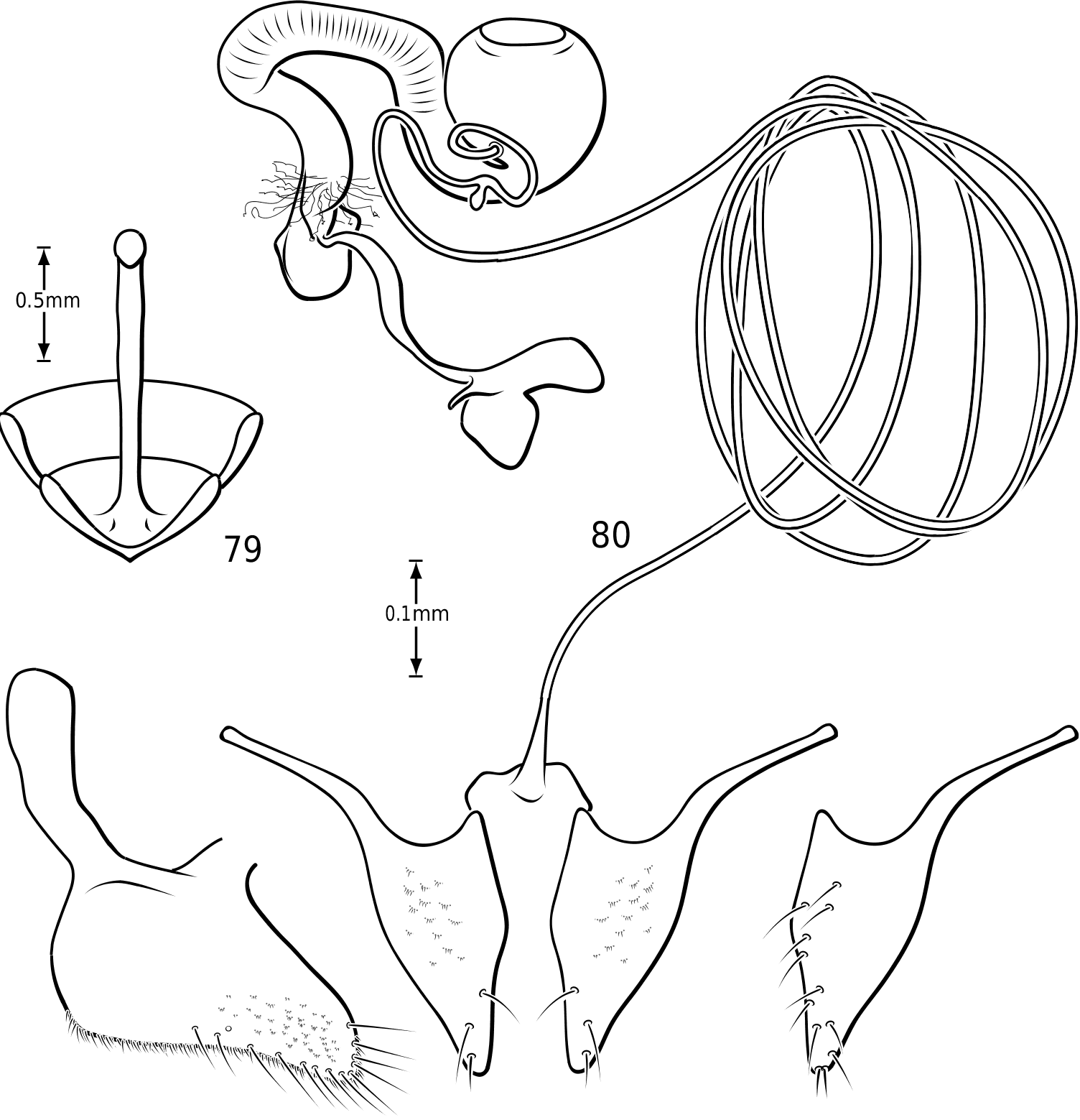
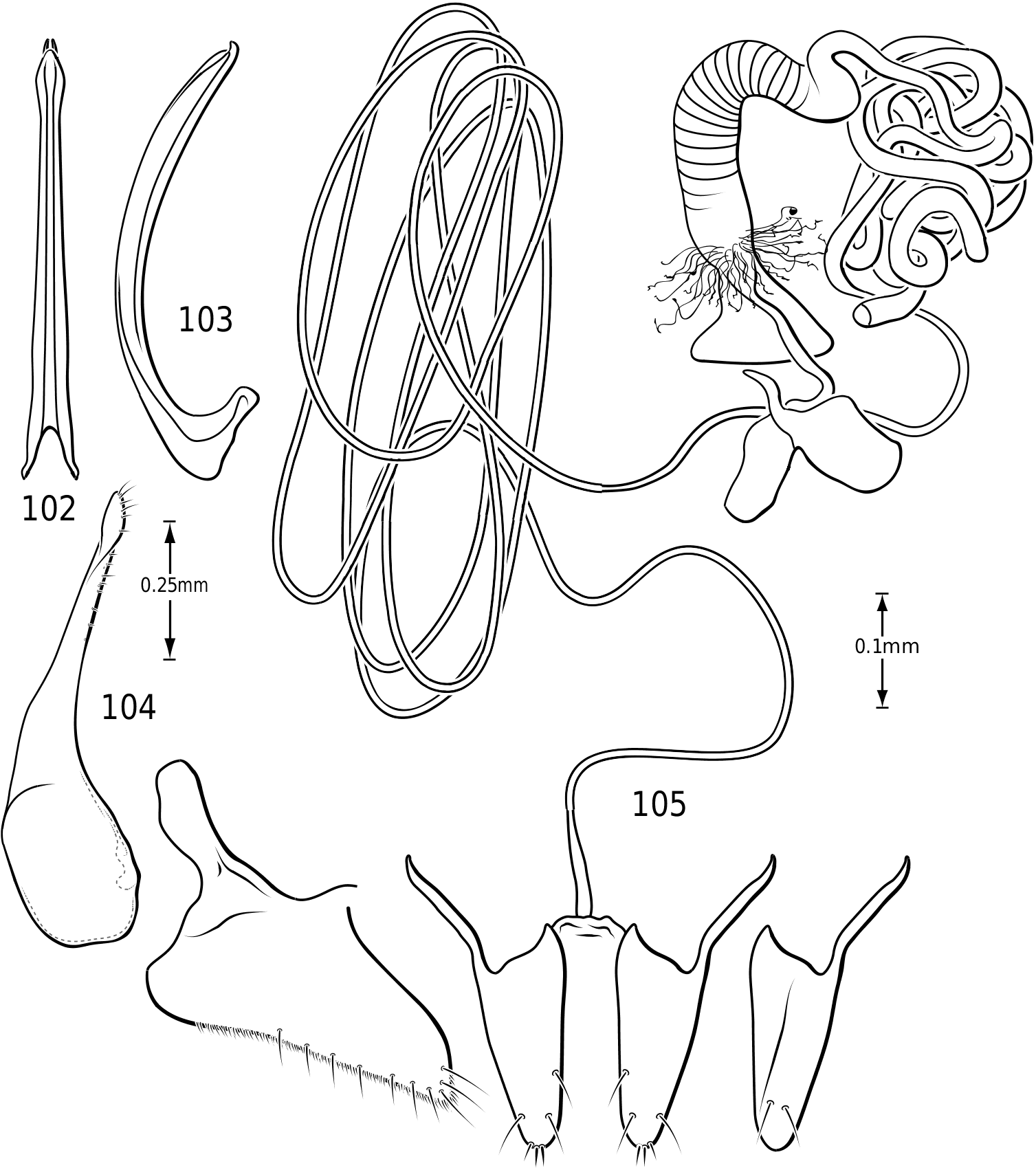
Abdomen: Visible sterna II–V are relatively unmodified compared with other Hydroporinae. However, sternum VI exhibits complex modification in all Vatellini . In vatellines there is an elongate, invaginated, heavily sclerotized structure extending from the apex of the sixth abdominal sternum internally along the dorsal surface of the abdominal sterna ( Figs 28–30 View Figures 28–37 ). Because of the unique nature of this feature, I have coined a new term for it, ‘ speleum ’, Latin for ‘cave’. The speleum is a hollow, sclerotized tube that extends from a broad opening along the apical margin of the sixth visible abdominal sternum. It is generally moreor-less flattened for much of its length and variously dorsoventrally undulate ( Fig. 29 View Figures 28–37 ). The anterior apex is variously expanded and may be dorsally curved ( Fig. 30 View Figures 28–37 ). The apex is typically very darkly pigmented and heavily sclerotized. There appear to be no muscles attached to the structure. The speleum is apparently the reservoir and duct of a large gland system. In fresh specimens of Vatellus and Derovatellus , a large region of white cells is visible surrounding the speleum with numerous tiny ducts emptying into the apex of the speleum or along its length ( Fig. 29 View Figures 28–37 ). Because welldeveloped pygidial glands are present in vatellines, the speleum is apparently not derived from their modification. The orifice forms a groove along the margin of the sixth visible sternum and the ventral margin of the apex may be variously lobed, particularly in Vatellus , or setose. The dorsal surface of the ventral margin of the orifice in Vatellus generally has a swelling that fits into a corresponding cavity on the ventral surface of the dorsal margin. The function of this gland structure is unknown. It occurs in both males and females of all species, though in some Derovatellus (e.g. D. fasciatus Régimbart , Fig. 33 View Figures 28–37 ), it is reduced. The speleum is species-specific in shape in many taxa, particularly in Derovatellus where it exhibits considerable variation, but also in Vatellus . There appears to be very little variation in shape within species. Because of damage to the type specimen, it is not known whether Calicovatellus petrodytes possesses a speleum. No apparently homologous features occur in this position in any of the many other Hydroporinae genera examined.
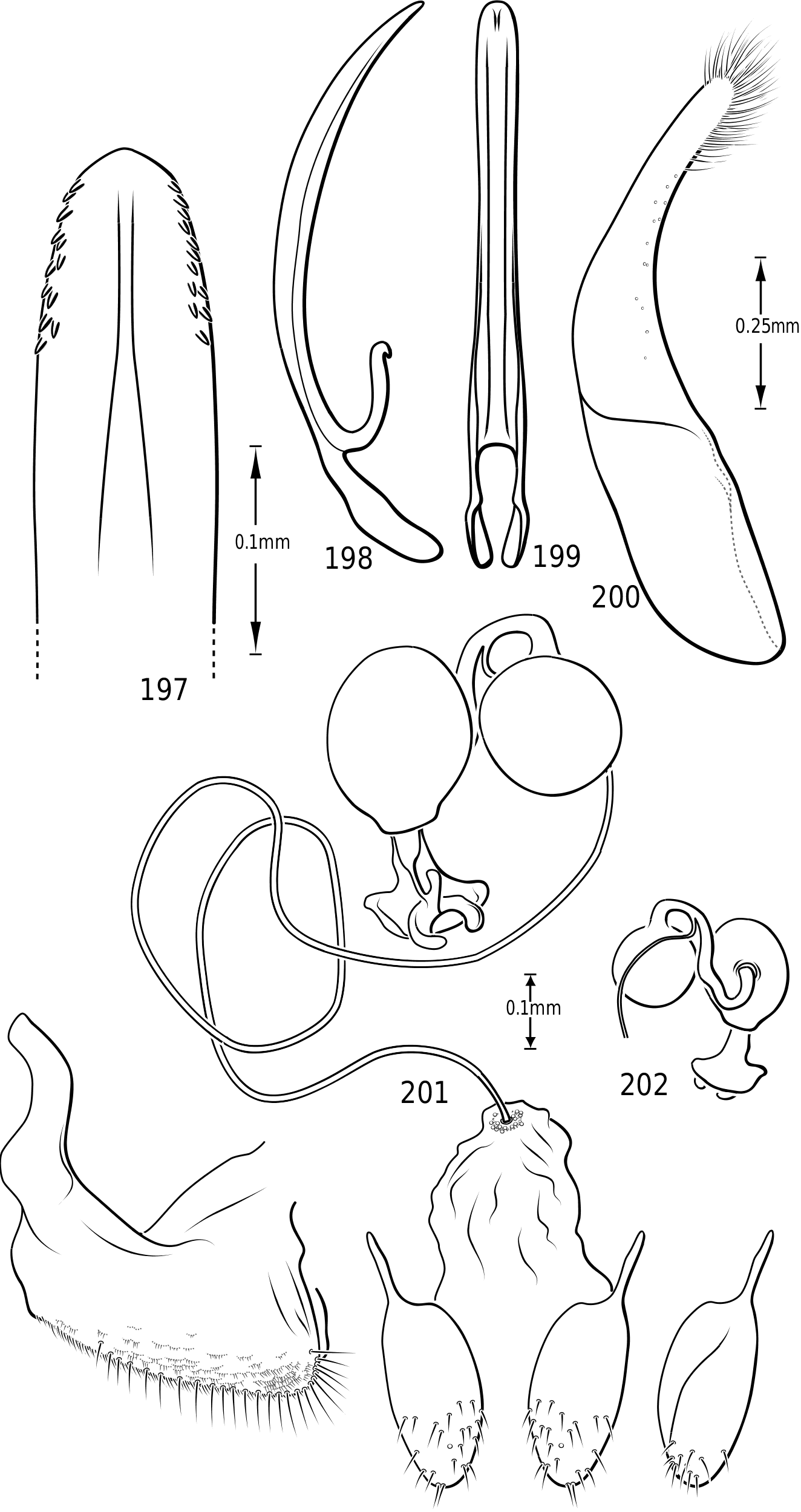
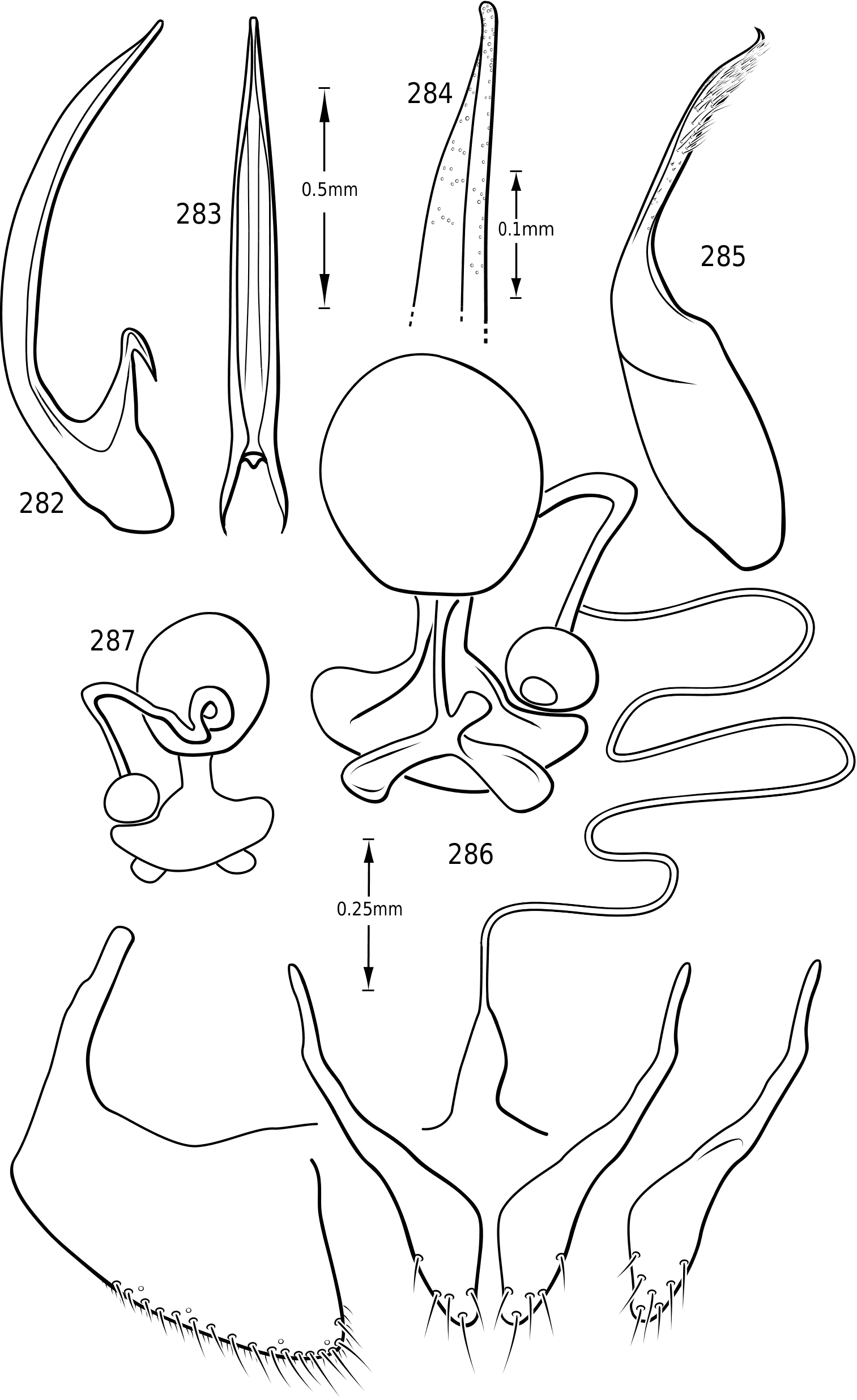
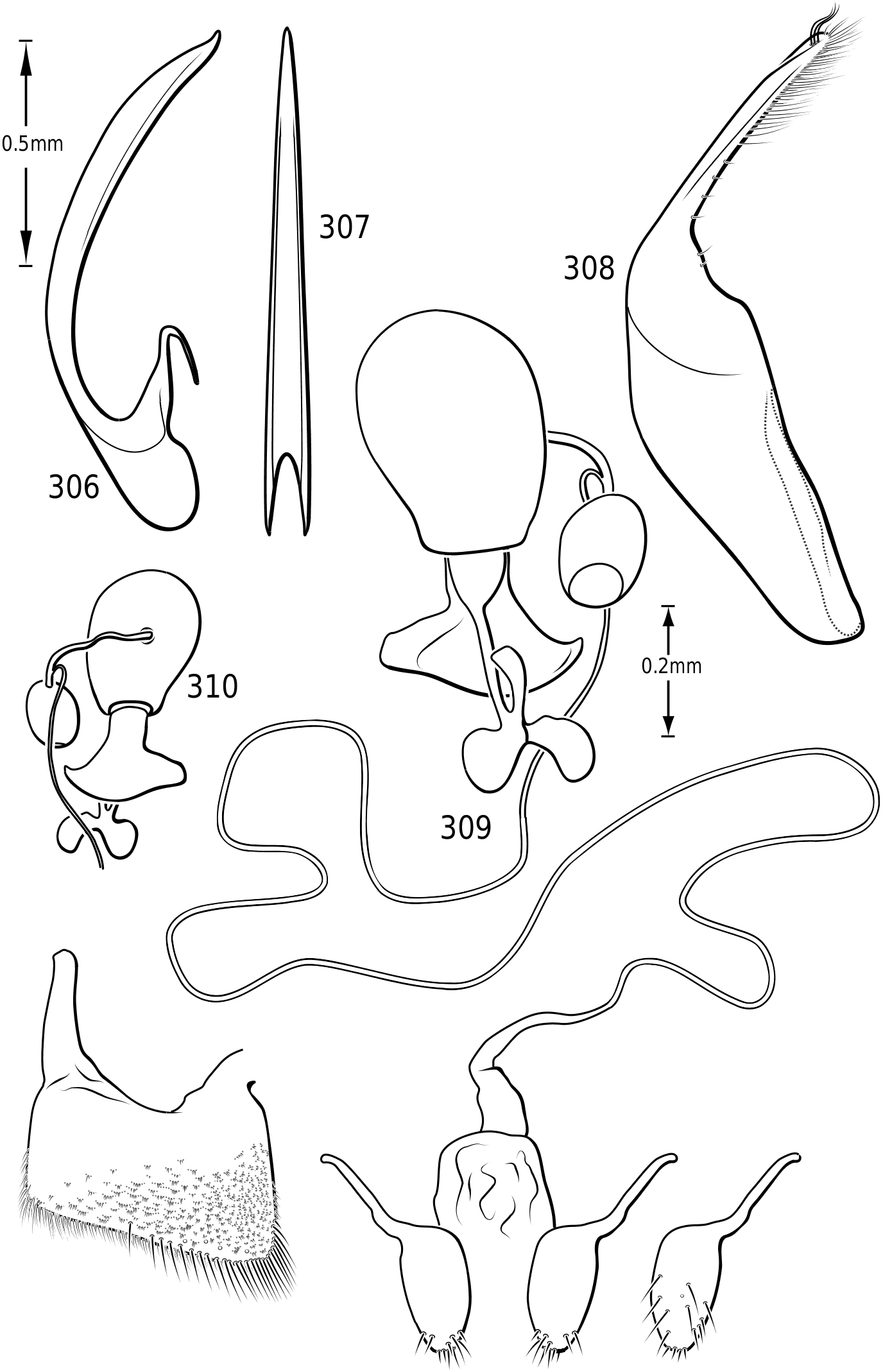
Male genitalia: As in other diving beetles, the genitalia of vatellines are rotated 90∞ in repose and 180∞ when extruded ( Balfour-Browne, 1940; Sharp & Muir, 1912). Thus the concave surface of the median lobe is actually the dorsal surface ( Miller & Nilsson, 2003). Members of the tribe have relatively unmodified, bilaterally symmetrical male genitalia. Members of Derovatellus exhibit considerable autapomorphic variation in shape of the median lobe, but do not exhibit many apparent synapomorphies. By contrast, members of Vatellus have the median lobe very long and evenly curved in general with relatively subtle differences in shape. However, they have several prominent features that appear to be synapomorphies within the group. In all species the basal portion of the lateral lobes are dramatically elongated anteriorly (e.g. Fig. 271 View Figures 267–273 ) and are fused along the dorsal margins. The base of the median lobe may also be extremely elongated (e.g. Fig. 269 View Figures 267–273 ) or this portion may be shorter (e.g. Fig. 125 View Figures 125–130 ). The apex of the median lobe varies in shape from species to species, though differences are often subtle. The apices of the lateral lobes are often laterally flattened and expanded (e.g. Fig. 200 View Figures 197–202 ), though they may be narrowed (e.g. Fig. 285 View Figures 282–287 ). Some species have two or three long, stout, curved, subapical setae on the ventral surface (e.g. Fig. 308 View Figures 306–310 ).
Members of Dytiscidae have a ring-like sclerite surrounding the genital capsule considered the joined last dorsal and ventral sclerites by Sharp & Muir (1912) and regarded by Roughley & Pengelly (1981), Franciscolo (1979) and others as the fused basal portion of sternum IX, pleuron IX and tergum IX. In Hydroporinae (and a few other dytiscids such as some Aciliini) the ventral sclerite in this ring is absent. In vatellines this ring is very elongate with the apices of the lateral arms expanded apically ( Figs 44, 47 View Figures 38–50 ). The apices are fused in Vatellus ( Fig. 44 View Figures 38–50 ) whereas in Derovatellus they are separated by clear membrane ( Fig. 47 View Figures 38–50 ). In Vatellus the anteromedial lobes of the fused sterna VIII are greatly prolonged anteriorly ( Fig. 42 View Figures 38–50 ), but
4546 0.25mm
44 47mm
0.25mm
50 48 49
they are short in Derovatellus ( Fig. 45 View Figures 38–50 ). The spiculum in Vatellus is long, expanded on each end and medially bent ( Fig. 43 View Figures 38–50 ). In Derovatellus it is short, not bent and expanded on one end ( Fig. 46 View Figures 38–50 ). As with other Dytiscidae , there is no differentiated internal sac. There is a sclerotized structure present on the ejaculatory duct ( Figs 48, 49 View Figures 38–50 ). This structure is present throughout Dytiscidae , but it has seemingly not been previously mentioned or described. There has been no published account of the potential function of this ejaculatory sclerite.
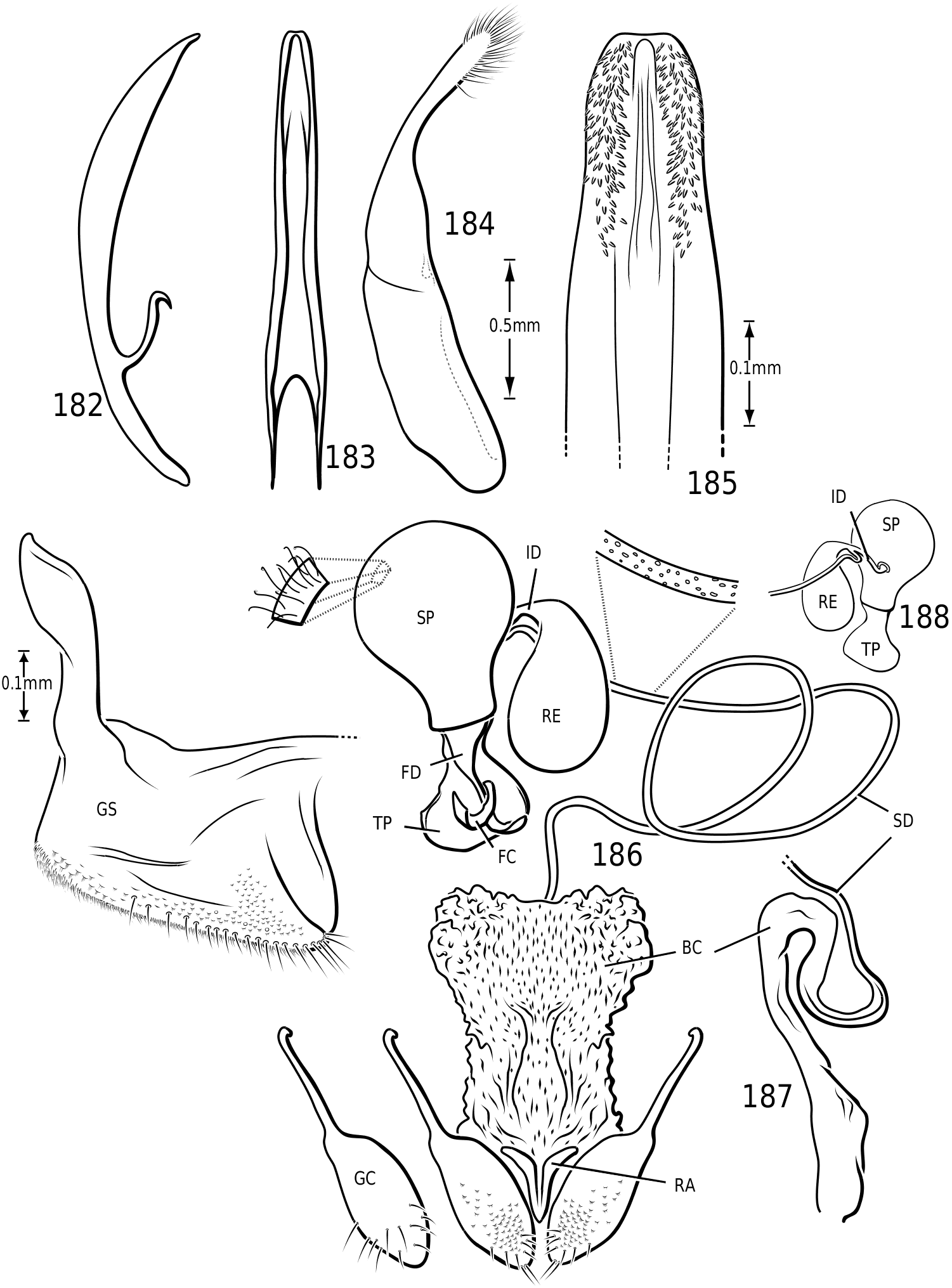
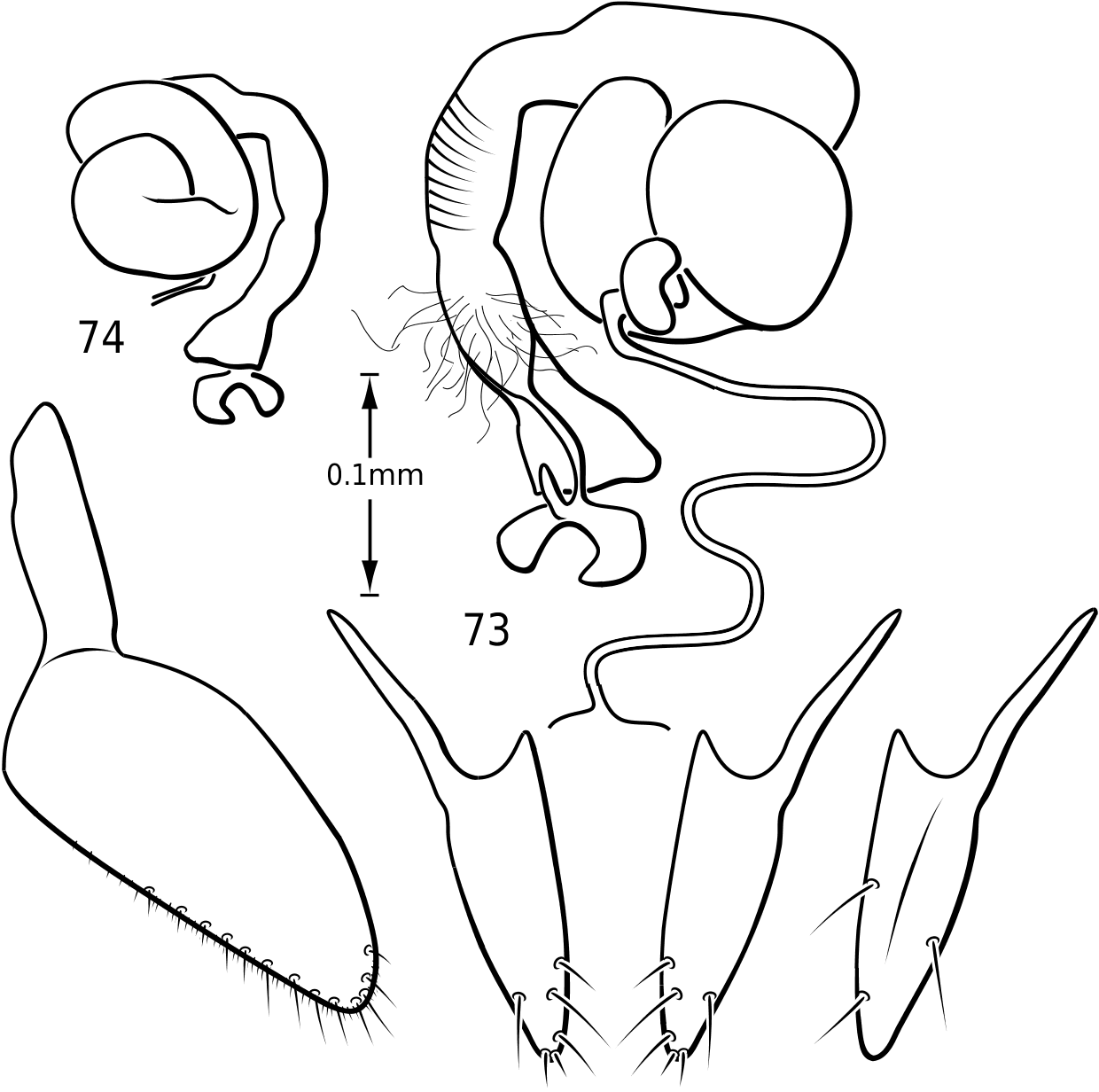
Female genitalia: Vatelline female genitalia are, in general configuration, similar to most ‘derived’ Hydroporinae [those taxa with Hydroporine-type female genitalia and receptacle present (Miller, 2001)]. There is considerable variation within the group, however. Members of Vatellus have relatively broad gonocoxosternites with long anterior lobes (e.g. Fig. 186 View Figures 182–188 ) whereas those of Derovatellus are elongate-triangular with the apices relatively acutely rounded and with long anterior lobes (e.g. Fig. 73 View Figures 73–74 ). In both there is generally a dense series of short, fine setae along the entire posterior margin and a series of long, stout, evenly spaced setae along the medial half of the posterior margin and posterior half of the medial margin (e.g. Figs 73 View Figures 73–74 , 186 View Figures 182–188 ). Members of both genera have gonocoxae relatively consistent with the generalized form
mm
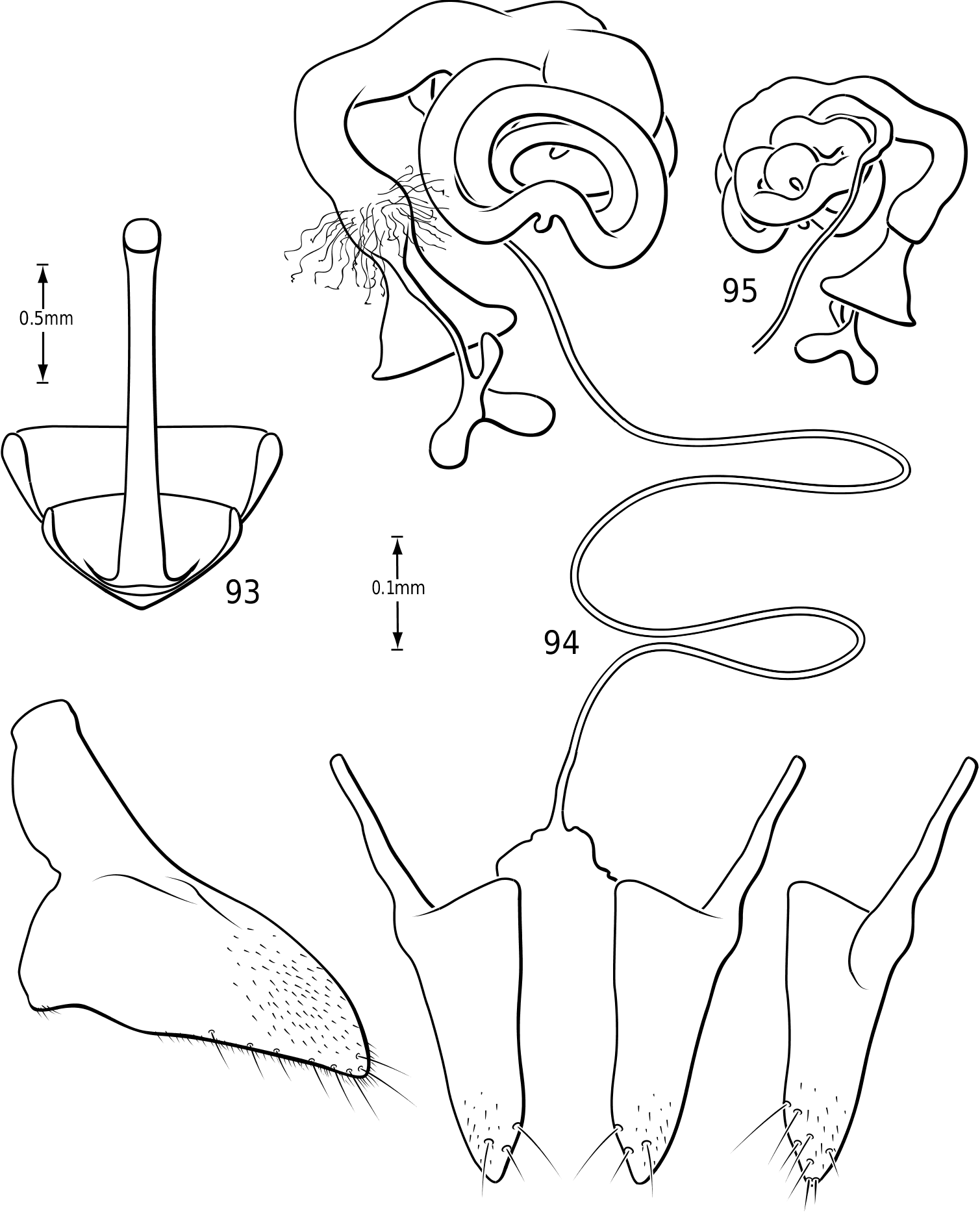
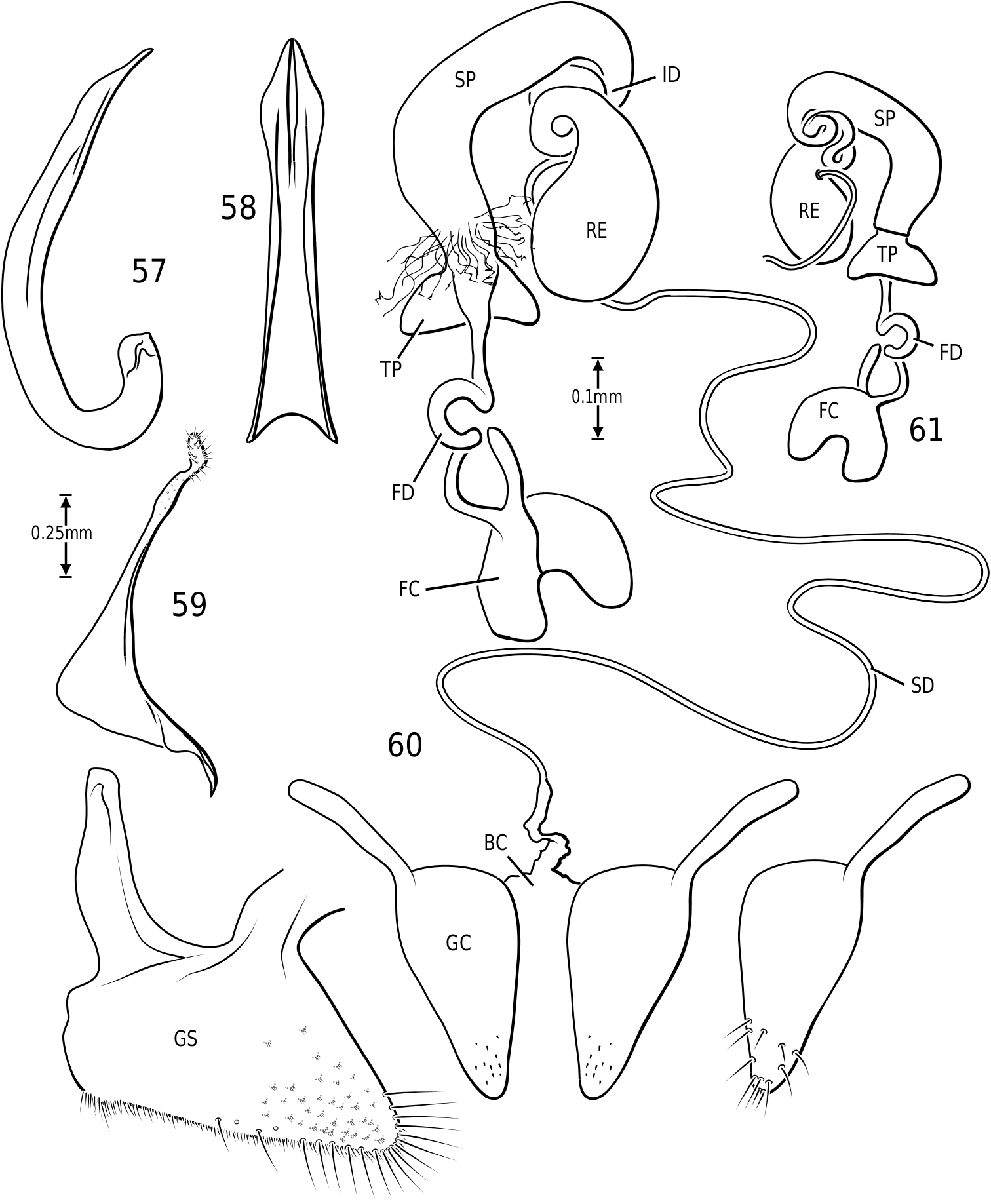
in Hydroporinae. Members of Vatellus are somewhat variable in the length and degree of roundedness of the gonocoxae whereas those of Derovatellus are generally elongate and apically pointed (e.g. Fig. 73 View Figures 73–74 ). The bursa in most taxa is small and relatively undifferentiated though there may be variation in the presence of lobes (e.g. Fig. 145 View Figures 139–146 ), unusual sculpture near the insertion of the spermathecal duct (e.g. Fig. 159 View Figures 155–160 ), size and shape. In one taxon, V. haagi , the bursa is exceptionally long, flattened and dorsoventrally sinuate ( Figs 186, 187 View Figures 182–188 ). The spermathecal duct varies from moderately long and slender in Vatellus and many Derovatellus to exceptionally long and very slender in a few Derovatellus (e.g. Fig. 80 View Figures 79–80 ). The spermathecal duct near the receptacle, the receptacle and the duct between it and the spermatheca are variable in length, shape and degree of curvature in Vatellus and extremely variable in Derovatellus . Close examination of the spermathecal duct reveals microstructure (e.g. Fig. 186 View Figures 182–188 ). The receptacle in Vatellus is generally relatively large and subspherical (e.g. Fig. 186 View Figures 182–188 ) though it may be reduced (e.g. Fig. 272 View Figures 267–273 ). In Vatellus the intermediate duct between the receptacle and spermatheca is generally relatively short, robust and somewhat sinuate (e.g. Figs 186, 188 View Figures 182–188 ), though in a few species it is very long and strongly coiled on the dorsal surface of the spermatheca (e.g. Fig. 258 View Figures 253–258 ). Members of Derovatellus have the receptacle variable from very large and anteriorly directed (e.g. Fig. 80 View Figures 79–80 ) to reduced or nearly absent (e.g. Fig. 105 View Figures 102–105 ). The intermediate duct may be short or nearly absent (e.g. Fig. 80 View Figures 79–80 ) to long, moderately and strongly twisted, or coiled (e.g. Figs 94, 95 View Figures 93–95 ). A small diverticulum is present on the spermathecal duct in some species (e.g. Fig. 80 View Figures 79–80 ). The spermatheca in vatellines comes in two general forms. In Vatellus it is subspherical anteriorly (e.g. Fig. 186 View Figures 182–188 ) whereas in Derovatellus it is elongate and bent medially (e.g. Fig. 60 View Figures 57–61 ). All vatellines have a broad, subtriangular, truncate process at the posterior apex of the spermatheca which varies in size (e.g. Figs 60 View Figures 57–61 , 186 View Figures 182–188 ). In D. roosevelti it is apically rounded rather than truncate in shape ( Fig. 80 View Figures 79–80 ). Members of both genera have the spermatheca bearing fine gland ducts. In Vatellus the glands cover the surface of the spermatheca ( Fig. 186 View Figures 182–188 ) whereas in Derovatellus the glands occupy a region near the origin of the fertilization duct (e.g. Fig. 105 View Figures 102–105 ). The fertilization duct is generally short but may be longer in some species (e.g. Fig. 80 View Figures 79–80 ). The ventral surface of the common oviduct bears a large, oval region of short spines in all species. Finally, the rami are relatively broad anteriorly and posteriorly convergent to a sharply pointed apex (e.g. Fig. 186 View Figures 182–188 ).
Larva: Third instar larvae of D. ibarrai (= D. floridanus ) and V. mexicanus were described by Spangler (1963, 1966b) and this diagnosis is based on his descriptions. Larvae of Vatellini are characterized by an exceptionally elongate and slender nasale that is apically expanded and rounded. The nasale bears a long, slender, curved process on each side. These rami are apically bifid. The nasale and the lateral rami are prominently spinous. Abdominal segments VII and VIII are entirely sclerotized. Abdominal segment VIII terminates in a very long, slender posterior projection that is subequal in length to the urogomphi. The urogomphi are unsegmented and bear many small setae.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.