
Temnodontosaurus zetlandicus, (Seeley, 1880)
|
publication ID |
https://doi.org/ 10.1093/zoolinnean/zlab118 |
|
publication LSID |
lsid:zoobank.org:pub:DACBA4A5-8D5F-479B-AFA5-E38B380F8962 |
|
DOI |
https://doi.org/10.5281/zenodo.6536848 |
|
persistent identifier |
https://treatment.plazi.org/id/2663F827-FFD5-AC51-124F-F0C6FAAFF9A9 |
|
treatment provided by |
Plazi |
|
scientific name |
Temnodontosaurus zetlandicus |
| status |
|
TEMNODONTOSAURUS ZETLANDICUS ( SEELEY, 1880) COMB. NOV.
1880 Ichthyosaurus zetlandicus – Seeley
1881 Ichthyosaurus longifrons – Owen
1889b Ichthyosaurus zetlandicus – Lydekker
1890 Ichthyosaurus zetlandicus – Woodward & Sherborn
1891 Ichthyosaurus zetlandicus – Fraas
1922 Stenopterygius zetlandicus – Huene
1925 Stenopterygius zetlandicus – Hauff
1974 Stenopterygius acutirostris – McGowan
1997 Temnodontosaurus acutirostris – Maisch & Hungerbühler
2000 Temnodontosaurus acutirostris – Maisch & Matzke
2003 Temnodontosaurus acutirostris – McGowan
2010 ‘ Ichthyosaurus ’ acutirostris – Maisch
2019? Temnodontosaurus acutirostris – Lomax
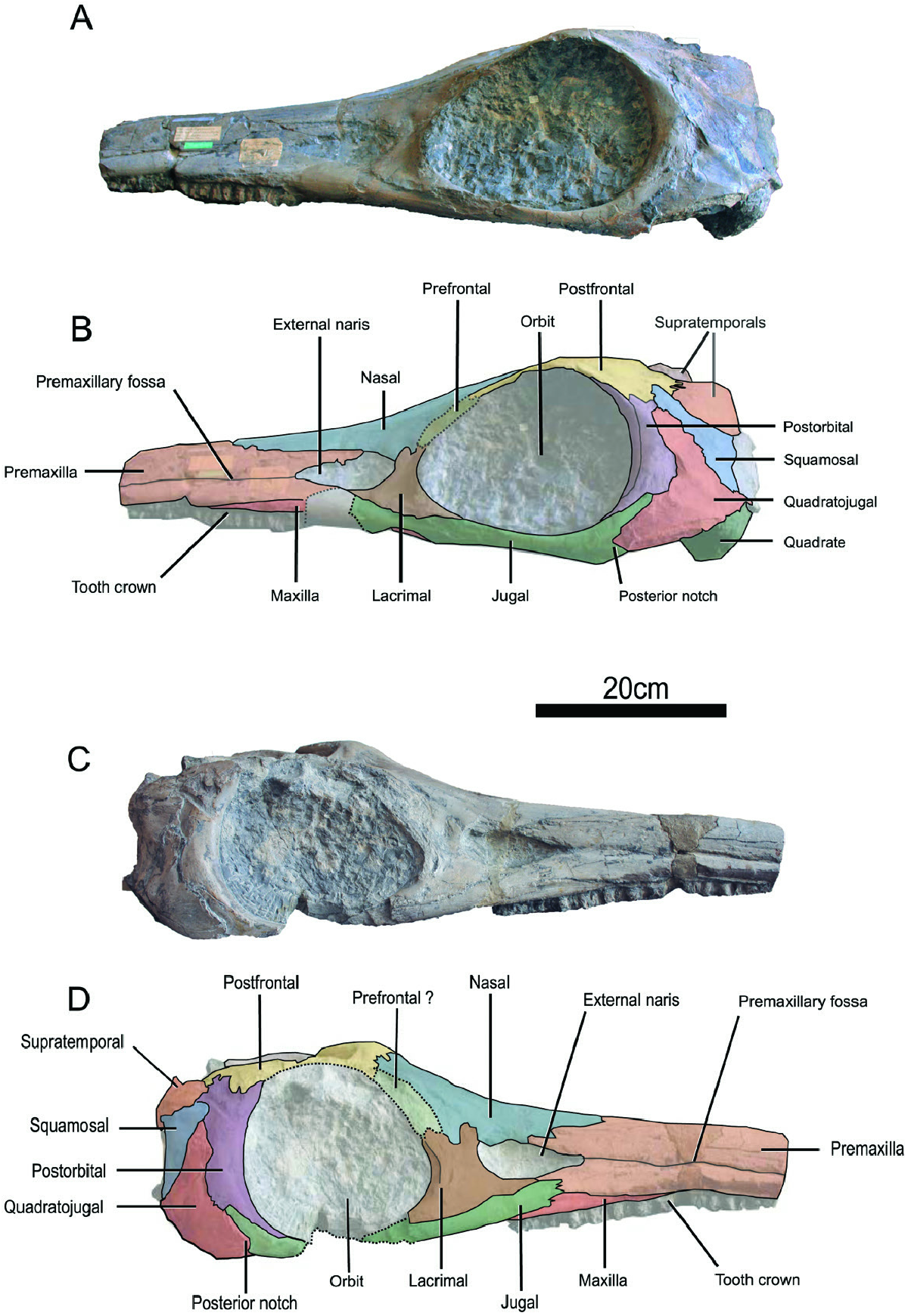
Holotype, stratum typicum and locus typicus: CAMSM J35176 View Materials , a three-dimensionally preserved skull missing the mandible and the anterior third of the rostrum, from the Whitby Mudstone Formation, Lower Toarcian, likely from the Loftus Alum Quarry close to Whitby, Yorkshire, UK.
Emended diagnosis: Temnodontosaurus zetlandicus is characterized by the following unique combination of unusual features among early neoichthyosaurians: a digitate anterior end of the jugal which covers the maxilla, externally separates the maxilla and the lacrimal and slightly overlaps the subnarial process of the premaxilla (shared with T. nuertingensis and T. trigonodon ); bifurcated posterior end of the jugal, resulting in the separation of the ascending process and a smaller anteroventrally oriented process (shared with T. trigonodon ); presence of a prominent mediolaterally oriented ridge which forms the anterior margin of the supratemporal fenestra (shared with T. trigonodon ); slender projection of the supratemporal which covers the postfrontal dorsally and does not reach the anterior margin of the supratemporal fenestra (shared with T. trigonodon ); presence of a prominent sagittal crest on the parietal and a slender parietal fork that partially overlaps the frontal (shared with T. trigonodon ); absence of a postnarial descending process; presence of two posterior processes of the nasal, the lateral ending by an interdigitated suture, overlapping the anterior edge of the postfrontal which thus is bifurcated and adopts a V-shape, resulting in the separation of a medial and an anterodorsal process; and dorsal region of the lacrimal less extended than in T. trigonodon and marked by a notch.
Comparative description of the holotype CAMSM J35176 View Materials : First described by Seeley in 1880, this specimen ( Figs 2–4 View Figure 2 View Figure 3 View Figure 4 ) is a cranium measuring 720 mm long; the mandible is not preserved. The end of the rostrum appears to have been broken off and we estimate the cranium and the snout to be respectively 990 mm and 680 mm long in vivo by prolonging the lateral edges of the premaxilla. The rostrum is straight, unlike in ‘ Ichthyosaurus ’ acutirostris ( Owen, 1865 –1881) even if this feature could be due to the taphonomic flattening. Some badly preserved teeth are located on the premaxilla and the maxilla ( Fig. 2 View Figure 2 ) and are marked by striations on the crown. The dorsal part of the basicranium is poorly preserved ( Fig. 4 View Figure 4 ); the posterior edge of the parietals, the supraoccipital, the exoccipitals and the dorsal part of the basioccipital are missing. Aside from these elements, the skull is well preserved in three dimensions. All elements of the braincase are made of finished bone and therefore do not have a rugose texture, especially the basioccipital for which the condyle is as smooth as the extracondylar area and for which the opisthotic facets are protruding. This suggests an adult or at least subadult ontogenetic stage for CAMSM J35176 View Materials ( Miedema & Maxwell, 2019).
The premaxilla is markedly elongated and not fully preserved ( Figs 2 View Figure 2 , 3 View Figure 3 ). The medial suture is clearly visible. Laterally, the premaxilla is marked by the presence of the premaxillary fossa and forms the supranarial and subnarial processes, as in other early parvipelvians even if this character may be variable in Hauffiopteryx ( Maxwell & Cortés, 2020) . The supranarial process forms approximately half of the dorsal margin of the naris as in Temnodontosaurus eurycephalus ( McGowan, 1974) and in Temnodontosaurus trigonodon ( Maisch, 1998b) . The suture of the supranarial process with the nasal is crenulated while the posterior extremity of subnarial process contacts the anterior end of the jugal as in T. trigonodon ( Maisch, 1998b) . The subnarial process also contacts the anterior edge of the lacrimal. Some badly preserved tooth roots are located on its ventral surface but cannot be precisely described.
The maxilla is dorsoventrally low, slender and extends anteriorly as far as the nasal ( Fig. 2 View Figure 2 ). This particular feature is typical of Temnodontosaurus ( Maisch & Hungerbühler, 1997; Martin et al., 2012; Swaby & Lomax, 2020) among early parvipelvians. The maxilla of T. zetlandicus seems to have a comparable size to that of ‘ I. ’ acutirostris ( Figs 2 View Figure 2 , 8 View Figure 8 ) and lower than in T. nuertingensis which has a higher maxilla ( Maisch & Hungerbühler, 1997). Moreover, the maxilla lacks a narial process and does not participate in the ventral margin of the external naris. The posterior part of the maxilla is covered by the jugal, thus excluding an external contact with the lacrimal as in T. trigonodon ( Maisch, 1998b; Pardo-Pérez et al., 2018).
The external naris has a typical droplet shape ( Fig. 2 View Figure 2 ) and is larger than in Ichthyosaurus De la Beche & Conybeare, 1821 ( Lomax & Massare, 2016) and Leptonectes ( Maisch & Matzke, 2003; McGowan, 1989). The anterior extremity is thinner than the posterior border which is exclusively formed by a shallow notch of the lacrimal.
The nasal is wide and robust, forming a fairly large portion of the dorsal region ( Fig. 3A, B View Figure 3 ). The anterior part of the nasal cannot be precisely distinguished due to the reconstructed area but appears to end abruptly as in Temnodontosaurus crassimanus (Blake, 1876) ( Swaby & Lomax, 2020) , T. nuertingensis ( Maisch & Hungerbühler, 1997) and T. trigonodon ( Pardo-Pérez et al., 2018) . The posterior half of the two nasals form a well demarcated internasal cavity. Posteriorly, the nasal forms two processes (one medial and one lateral) overlapping the anterior part of the postfrontal ( Fig. 3A, B View Figure 3 ). The medial process has no digitation and exclusively contacts the postfrontal medially. Therefore, there is no nasal-parietal contact, at least externally. The lateral process is located more laterally and marked by a three-finger digitation. Laterally, the nasal forms approximately half of the dorsal margin of the external naris and a lateral wing is absent, as is the case in all early parvipelvians with the exception of Leptonectes tenuirostris (Conybeare, 1822) ( Maisch & Matzke, 2003; Massare et al., 2021) and Stenopterygius aaleniensis Maxwell et al., 2012 ( Massare et al., 2021). Furthermore, the nasal does not make a postnarial descending process, unlike Temnodontosaurus platyodon (Conybeare, 1822) (e.g. NHMUK R1158), T. nuertingensis (Maisch, 1997; Massare et al., 2021) and T. trigonodon ( Maisch, 1998b; Maisch & Hungerbühler, 2001).
The lacrimal has a broad hatchet shape, as is usual for ichthyosaurians ( Fig. 2 View Figure 2 ). The posterior margin participates to the anterior and anteroventral margins of the orbit while the anterior margin also forms the posterior margin of the external naris without forming a rounded bulge or a prominent process within the naris, as can be seen in many specimens of Temnodontosaurus ( Massare et al., 2021) . The lacrimal is marked by a notch on the dorsal edge unlike in Temnodontosaurus platyodon ( Godefroit, 1993a) , Leptonectes spp. ( Maisch & Matzke, 2003), Suevoleviathan Maisch, 1998 spp. (Maisch, 2001) , Ichthyosaurus spp. ( Lomax & Massare, 2016), Protoichthyosaurus Appleby, 1979 ( Lomax & Massare, 2018) and Stenopterygius spp. ( Caine & Benton, 2011) and is not as extended dorsally as in T. trigonodon where the dorsal notch is also absent ( Maisch, 1998b; Maisch & Hungerbühler, 2001; Pardo-Pérez et al., 2018). Moreover, the suture with the prefrontal is slightly crenulated. The ventral edge is more convex and entirely in contact with the jugal.
The prefrontal accounts for approximatively one third of the dorsal margin of the orbit ( Figs 2 View Figure 2 , 3A, B View Figure 3 ). The anterior extremity contacts the lacrimal and this suture appears to be weakly crenulated. The prefrontal does not form a narial process, and hence does not participate to the rim of the external naris, a feature that is shared with most early parvipelvians with the exception of Hauffiopteryx Maisch, 2008 ( Maxwell & Cortés, 2020). The prefrontal is not dorsomedially extended and does not contact the frontal or the parietal, unlike in Leptonectes ( Maisch & Matzke, 2003) , Stenopterygius ( Maxwell et al., 2012) and Hauffiopteryx ( Maxwell & Cortés, 2020) . The suture with the postfrontal seems to be slightly interdigitated.
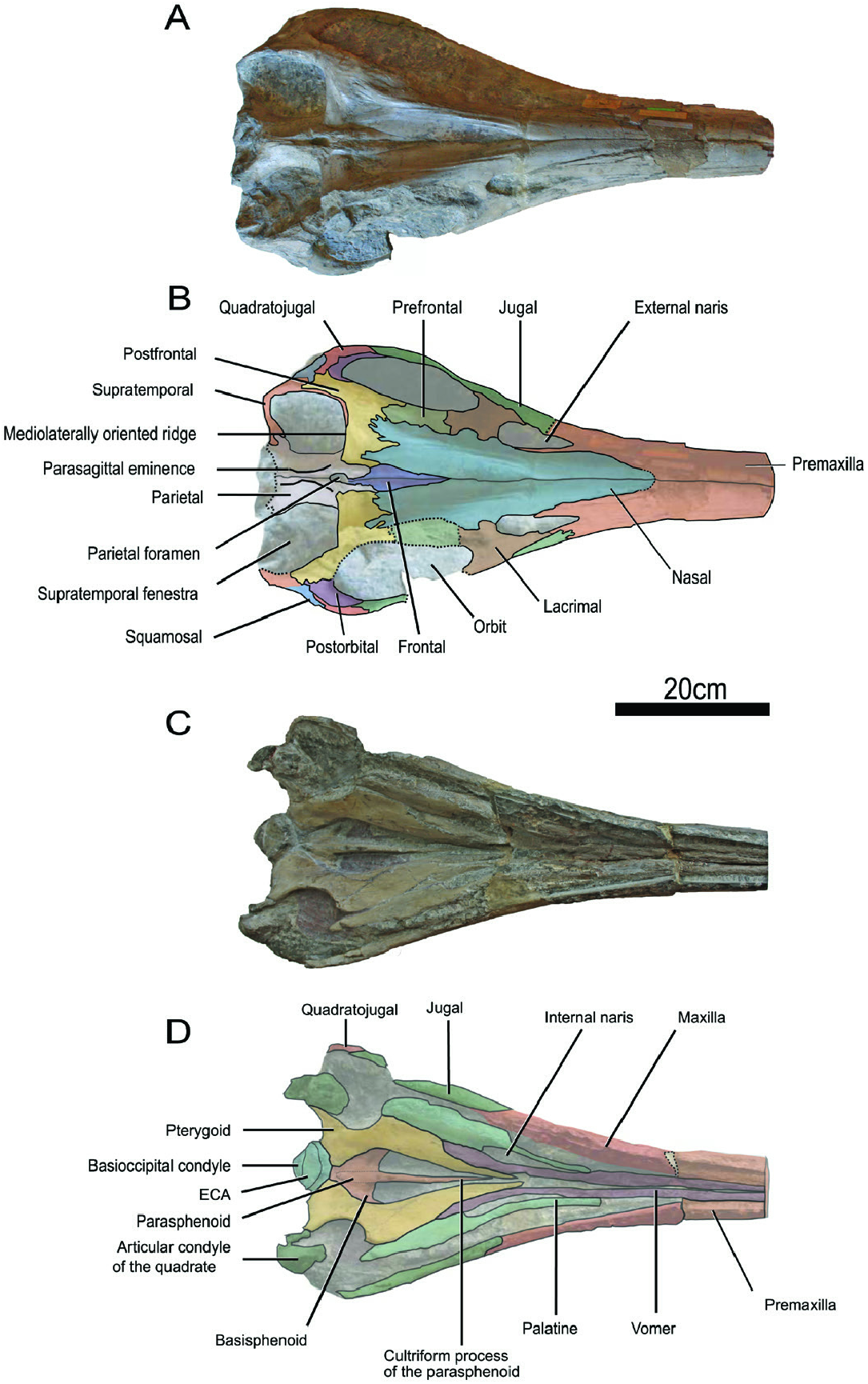
Externally, the frontal is a small lanceolate bone that forms a narrow triangular process anteriorly ( Fig. 3A, B View Figure 3 ). The posterior part is wider and is covered by the slender parietal fork, as in T. trigonodon ( Maisch, 1998b) and therefore differs from Hauffiopteryx ( Maxwell & Cortés, 2020) , Ichthyosaurus ( McGowan, 1973) , Leptonectes ( Maisch & Matzke, 2003; Vincent et al., 2014), Protoichthyosaurus ( Lomax et al., 2020) and Stenopterygius ( Maxwell et al., 2012) in which the parietal anterior termination is broader. The frontal only participates in the formation of the anterior margin of the parietal foramen as in T. trigonodon ( Maisch, 1998b) , but unlike in Hauffiopteryx ( Maxwell & Cortés, 2020) , Leptonectes ( Maisch & Matzke, 2003) , Protoichthyosaurus ( Lomax et al., 2020) , Stenopterygius ( Maxwell et al., 2012) or Wahlisaurus Lomax, 2016, in which frontals also form the lateral margin of the parietal foramen. Laterally, the frontal does not reach the supratemporal fenestra, as in all early parvipelvians ( McGowan, 1973; Maisch & Matzke, 2003; Maxwell et al., 2012; Marek et al., 2015; Lomax, 2016; Maxwell & Cortés, 2020).
The jugal is slender and forms the ventral and posteroventral margins of the orbit ( Fig. 2 View Figure 2 ). The posterior extremity of the jugal contacts the postorbital and the quadratojugal, unlike in Hauffiopteryx ( Maxwell & Cortés, 2020) and Stenopterygius ( Maisch, 2008) . This extremity appears bifurcated, resulting in the separation of the ascending process and a small anteroventrally oriented process ( Fig. 2C, D View Figure 2 ). Such a bifurcation is also found in T. trigonodon and could be due to an anteroventral process of the quadratojugal ( Maisch & Hungerbühler, 2001). However, the ascending process is broader in T. trigonodon ( Maisch & Hungerbühler, 2001; Pardo-Pérez et al., 2018). The anterior extremity of the jugal is digitated, contacting the subnarial process of the premaxilla and covering the posterior part of the maxilla ( Fig. 2 View Figure 2 ). This contact between the jugal and the premaxilla is also present in T. trigonodon ( Maisch, 1998b; Fig. 12 View Figure 12 ) and in T. nuertingensis ( Maisch & Hungerbühler, 1997) , but not in other early parvipelvians.
The quadratojugal appears to form the frame of the postorbital region and is well exposed ( Fig. 2 View Figure 2 ) as in Ichthyosaurus ( Lomax & Massare, 2016) and Temnodontosaurus ( McGowan, 1974; Godefroit, 1993a; Maisch & Hungerbühler, 2001) and unlike in Hauffiopteryx ( Maxwell & Cortés, 2020) and Stenopterygius ( Godefroit, 1993b; Maisch, 2008). The quadratojugal is wide, especially the ventral part where it extends to the level of the posterior digitation of the jugal (as in T. trigonodon ; Maisch & Hungerbühler, 2001). The quadratojugal forms a large dorsal process, which nearly reaches the dorsal margin of the squamosal ( Fig. 2 View Figure 2 ). The articulation surface with the quadrate is strongly concave medioventrally. The anterior edge the quadratojugal seems to slightly cover the posterior margin of the postorbital. Posterodorsally, the quadratojugal contacts the squamosal along a relatively smooth suture. The postorbital region is large compared to the orbit but less than in T. trigonodon even if this ratio looks variable within the species (ratio length of the postorbital region to length of the orbit: ~0.5 for T. zetlandicus vs. ~0.6–1.0 for T. trigonodon ; see Supporting Information, Table S1 View Table 1 ).
The squamosal is a relatively large triangular element within the postorbital region ( Fig. 2 View Figure 2 ) and has a morphology similar to that of T. trigonodon ( Maisch & Hungerbühler, 2001) . The posteroventral process of the squamosal is long and reaches the articulation between the quadratojugal and the condyle of the quadrate. Laterally, the suture with the quadratojugal is well extended and dorsally, the squamosal contacts the supratemporal and the postfrontal by an anterodorsal process ( Fig. 2 View Figure 2 ).
The crescent-shaped postorbital forms the posterior margin of the orbit ( Fig. 2 View Figure 2 ). Dorsally, the facet with the postfrontal is reduced in comparison to Suevoleviathan (Maisch, 2001) and ventrally, the postorbital contacts the jugal. The orbital crest is not prominent on postorbital, thus resulting in a lack of clear delimitation of the part involved in the formation of the orbit and the part forming the postorbital region. The postorbital thins dorsally and ventrally, buttressing the postfrontal and the dorsal margin of the jugal. Posteriorly, the postorbital contacts the quadratojugal and seems to be slightly overlapped by this element.
The parietals are relatively well preserved except at their posterior part and have a curved aspect ( Fig. 3A, B View Figure 3 ). The anterior extremity of the parietal is marked by a slender parietal fork that covers the posterior part of the frontal as in T. trigonodon ( Fig. 12 View Figure 12 , B, C; Maisch, 1998b). As a result, the posterior and lateral margins of the parietal foramen are formed externally by this element and this foramen is nearly enclosed by the parietals except at its anterior edge ( Fig. 3A, B View Figure 3 , 7 View Figure 7 ). This strongly contrasts with Hauffiopteryx ( Marek et al., 2015) , Stenopterygius ( Maxwell et al., 2012) and Wahlisaurus (Lomax, 2016), where the parietal foramen is nearly completely enclosed by the frontals. The interparietal suture is surrounded by a prominent parasagittal ridge ( Fig. 3A, B View Figure 3 ), which is present on both elements, thus forming a paired structure as in T. trigonodon ( Fig. 12 View Figure 12 ). This ridge becomes thinner and flatter posteriorly and ends anteriorly at the posterior edge of the parietal foramen. Dorsally, the suture between the parietal and the supratemporal adopts a sinusoidal configuration.
The supratemporal is a strongly curved bone ( Fig. 3A, B View Figure 3 ). It forms the posterior and lateral margins of the supratemporal fenestra, forming an elongated anteromedial process that covers the postfrontal laterally and reaches the anterior ridge of the supratemporal fenestra ( Fig. 3A, B View Figure 3 ) formed by the dorsal part of the postfrontal (see below) as in T. trigonodon ( Fig. 12 View Figure 12 ). The suture with the postfrontal is digitated and the contact with the squamosal is extensive ( Fig. 2 View Figure 2 ). The descending process of the supratemporal forms the lateral margin of the braincase, where it contacts the occipital lamella of the quadrate ( Fig. 4 View Figure 4 ).
The left postfrontal is completely preserved while the anterolateral portion of the right one is damaged ( Figs 2 View Figure 2 , 3A, B View Figure 3 ). The postfrontal is an extensive structure participating in the dorsal and postorbital regions of the cranium. Anteriorly, this element is partially covered by the posterior processes of the nasal, giving it a V-shape dorsally, resulting in the bifurcation of a medial and a lateral process, which it is not seen in any early parvipelvians ( Maisch, 1998b; Maisch & Matzke, 2003; Maxwell et al., 2012; Vincent et al., 2014; Marek et al., 2015; Lomax, 2016; Lomax & Massare, 2016; Lomax et al., 2020; Maxwell & Cortés, 2020). The suture with the prefrontal is not well preserved but appears undulated. The postfrontal forms the entire anterior margin of the supratemporal fenestra where this element forms an extensive mediolaterally oriented ridge ( Fig. 3A, B View Figure 3 , 7 View Figure 7 ). This unusual structure is actually shared with T. trigonodon ( Fig. 12 View Figure 12 ). A short ridge is also present in Protoichthyosaurus prostaxalis Appleby, 1979 ( Lomax & Massare, 2018). However, this postfrontal ridge appears shorter than in T. zetlandicus (medially vanished around mid-supratemporal fenestra width) and also appears to be oriented along the anteromedial-posterolateral axis whereas this ridge is mediolaterally oriented in T. zetlandicus ( Figs 3A, B View Figure 3 ). Apart from these three species, the presence of a postfrontal ridge which forms the anterior edge of the supratemporal fenestra is absent in all other early parvipelvians ( McGowan, 1973; Maisch, 1998a; Maisch & Matzke, 2003; Maxwell et al., 2012; Vincent et al., 2014; Marek et al., 2015; Lomax, 2016; Lomax & Massare, 2016; Lomax et al., 2020; Maxwell & Cortés, 2020).
The basioccipital is partially preserved, having its dorsal part damaged ( Fig. 4 View Figure 4 ). Thus, its contribution to the foramen magnum cannot be characterized. The condyle is prominent and is clearly separated from the extracondylar area, a character shared by members of Temnodontosaurus ( Swaby & Lomax, 2020) and more generally, non-ophthalmosaurid parvipelvians ( Motani, 1999b; Fischer et al., 2012) and the notochordal pit is located on the centre of the condyle. The extracondylar area is slightly expanded laterally and seems to be well developed ventrally and marked by the presence of a basal tubera, thus being saddle-shaped ventrally, unlike in Stenopterygius quadriscissus (Quenstedt, 1856) ( Miedema & Maxwell, 2019) and Chacaicosaurus Fernández, 1994. The protrusion of the opisthotic and stapedial facets is discernible, resulting in concave and well-delimited facets.
The stapes possesses a robust shaft that is only slightly waisted ( Fig. 4 View Figure 4 ). The proximal head seems to be massive as in T. trigonodon ( Maisch, 2002) , Eurhinosaurus longirostris (Mantell, 1851) (e.g. SMNS 18648), Hauffiopteryx ( Marek et al., 2015) and Stenopterygius quadriscissus ( Miedema & Maxwell, 2019) but unlike in Ichthyosaurus ( McGowan, 1973) , Protoichthyosaurus ( Lomax et al., 2019) and Temnodontosaurus azerguensis Martin et al., 2012 . The medial surface of the proximal head is concave, matching the slightly convex corresponding facet on the basioccipital. The distal facet for articulation with the quadrate is elliptic and weakly expanded with respect to the shaft.
Both quadrates are poorly preserved ( Fig. 4 View Figure 4 ). The dorsal suture with the supratemporal and the laterally to the right quadrate with the stapes is nevertheless visible. The condyle, located at the ventrolateral end of the posterior face, is better preserved and massive. With a reniform aspect, the dorsolateral edge adopts a shape complementary to the contact facet of the quadratojugal and is therefore largely convex. The ventral edge of the condyle is more concave and would serve to accommodate the surangular which is not preserved. The ventromedial edge contacts the quadrate wing of the pterygoid.
The pterygoid is a long robust element, markedly constricted at mid-length ( Fig. 3C, D View Figure 3 ), as is usually the case in parvipelvians ( McGowan, 1973). The markedly concave borders of the pterygoid delimit the suborbital fenestra laterally and the large interpterygoid vacuity. The narrow palatine ramus of the pterygoid sharply terminates medially of the internal naris. Laterally, the pterygoid contacts the palatine, and also the vomer more anteriorly by a long straight suture, and there is no postpalatine process. The pterygoid expands posteriorly, forming two horizontal lamellae in the region of the quadrate ramus. The larger lateral lamella contacts the condyle of the quadrate and the smaller medial one contacts the ventral extremity of the extracondylar area of the basioccipital.
The palatine is fairly elongated and forms posteriorly the anterior margin of the suborbital fenestra ( Fig. 3C, D View Figure 3 ). Its concave medial edge forms the lateral margin of the internal naris. The anterior half of the palatine becomes considerably thinner and its extremity ends by a junction with the vomer.
The vomer is also elongated; its widest portion is set posteriorly, where it forms the medial margin of the internal naris ( Fig. 3C, D View Figure 3 ). The vomer extends anteriorly as a slit-like process, contacting the premaxilla and the palatine. Posteriorly the vomer is not well extended as in T. nuertingensis and almost excludes a contact between the pterygoid and the palatine ( Maisch & Hungerbühler, 1997).
The basisphenoid appears crescent shaped in ventral view as its anterior edge is concave ( Fig. 3C, D View Figure 3 ). The posterior surface in contact with the basioccipital is hidden laterally by the pterygoids, as well as the basipterygoidal processes laterally. The basisphenoid is medially crossed throughout its entire length by the parasphenoid; even though the carotid foramina is not visible, this extension of the parasphenoid indicates that this foramen was paired like all members of Temnodontosaurus ( Fraas, 1913; von Huene, 1931; Godefroit, 1993a; Maisch & Matzke, 2000; Maisch, 2002; Martin et al., 2012).
The parasphenoid forms a relatively robust cultriform process ( Fig. 3C, D View Figure 3 ). The parasphenoid extends from the posterior end of the basisphenoid to approximately half the length of the palate region but does not reach the interpterygoidal suture.




| NHMUK |
Natural History Museum, London |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Temnodontosaurus zetlandicus
| Laboury, Antoine, Bennion, Rebecca F., Thuy, Ben, Weis, Robert & Fischer, Valentin 2022 |
Ichthyosaurus longifrons
| Owen 1881 |
Ichthyosaurus
| De la Beche & Conybeare 1821 |